
Common Ragweed (Ambrosia artemisiifolia L.) Causes Severe Yield Losses in Soybean and Impairs Bradyrhizobium japonicum Infection

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Greenhouse Trial
2.2. Field Trial
2.3. Measurements
2.4. Statistical Analysis
3. Results
3.1. Growth Performance of Soybean
3.1.1. Greenhouse Trial
3.1.2. Field Trial
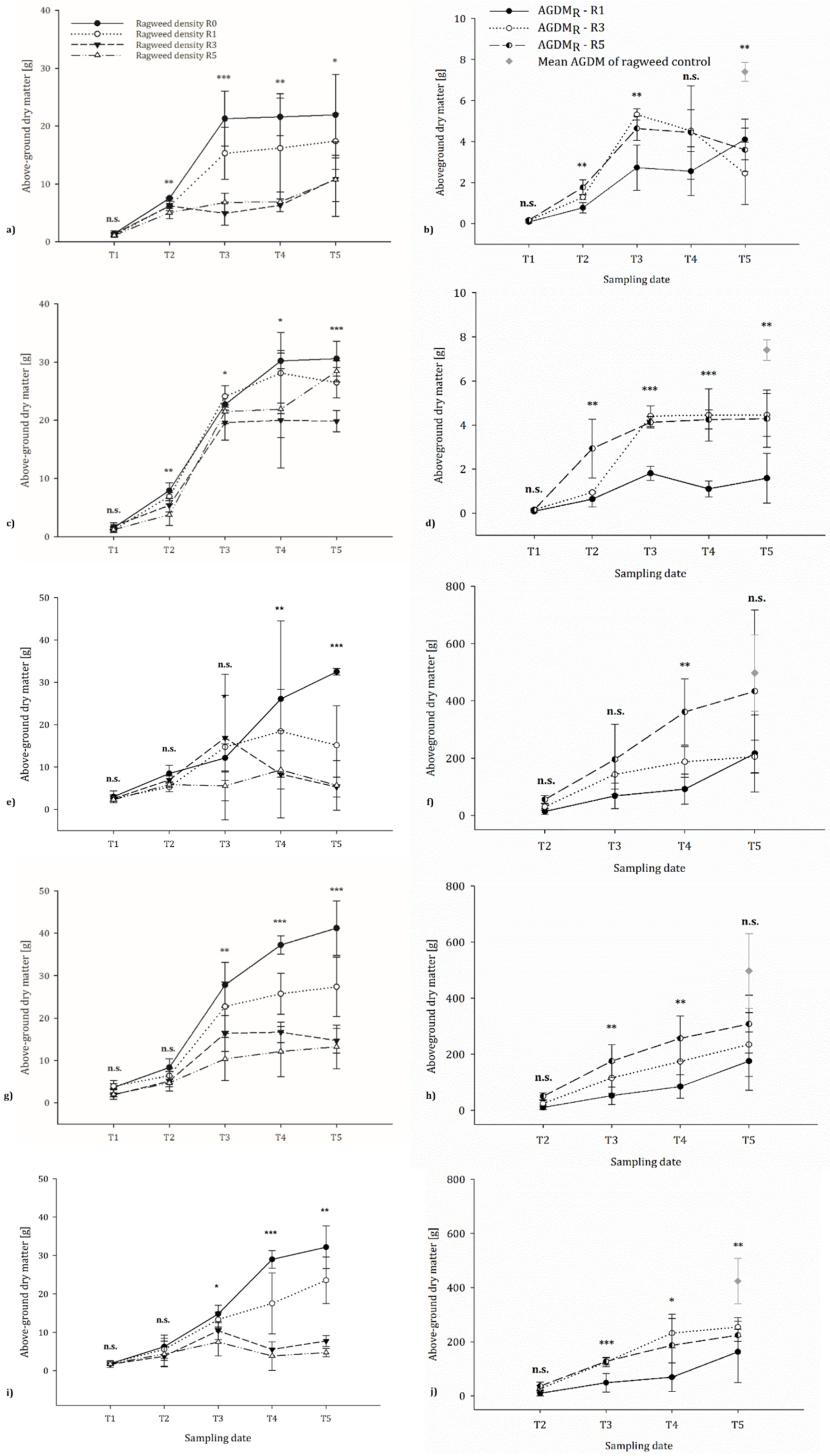
3.2. Growth Performance of Ragweed
3.2.1. Greenhouse Trial
3.2.2. Field Trials
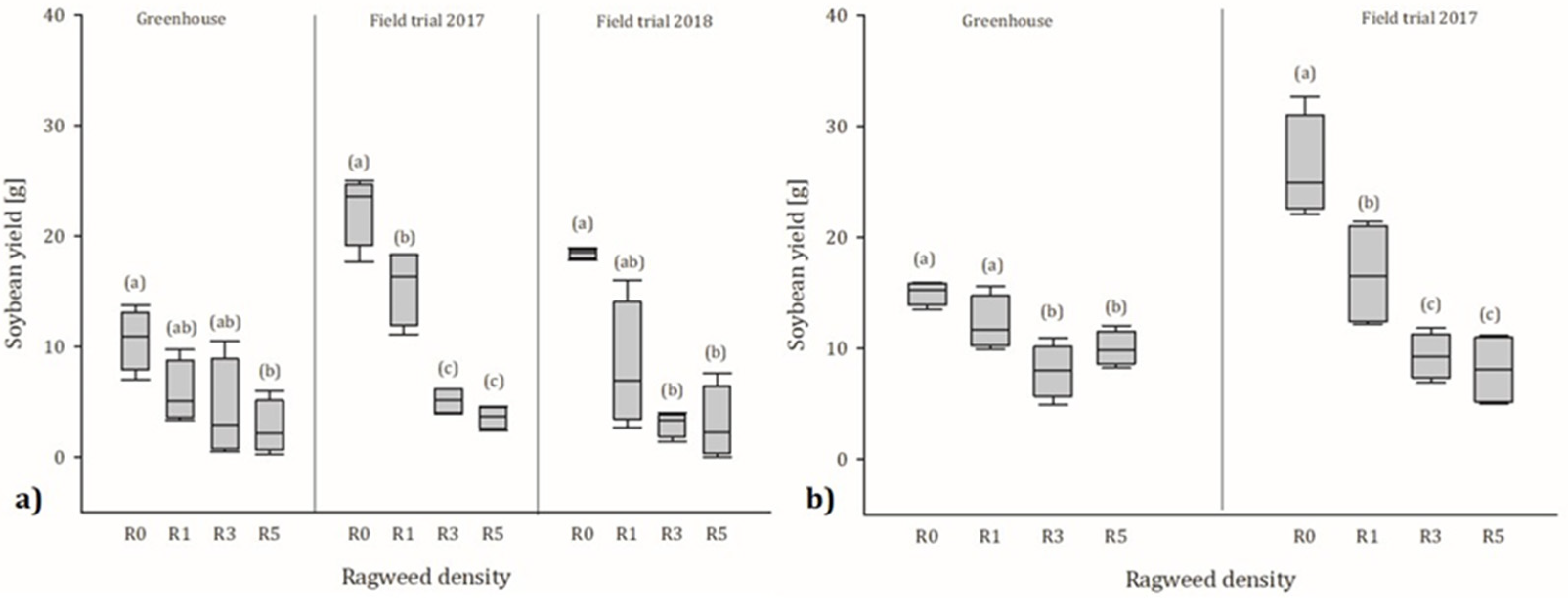
3.3. Yield of Soybean
3.3.1. Greenhouse Trial
3.3.2. Field Trials
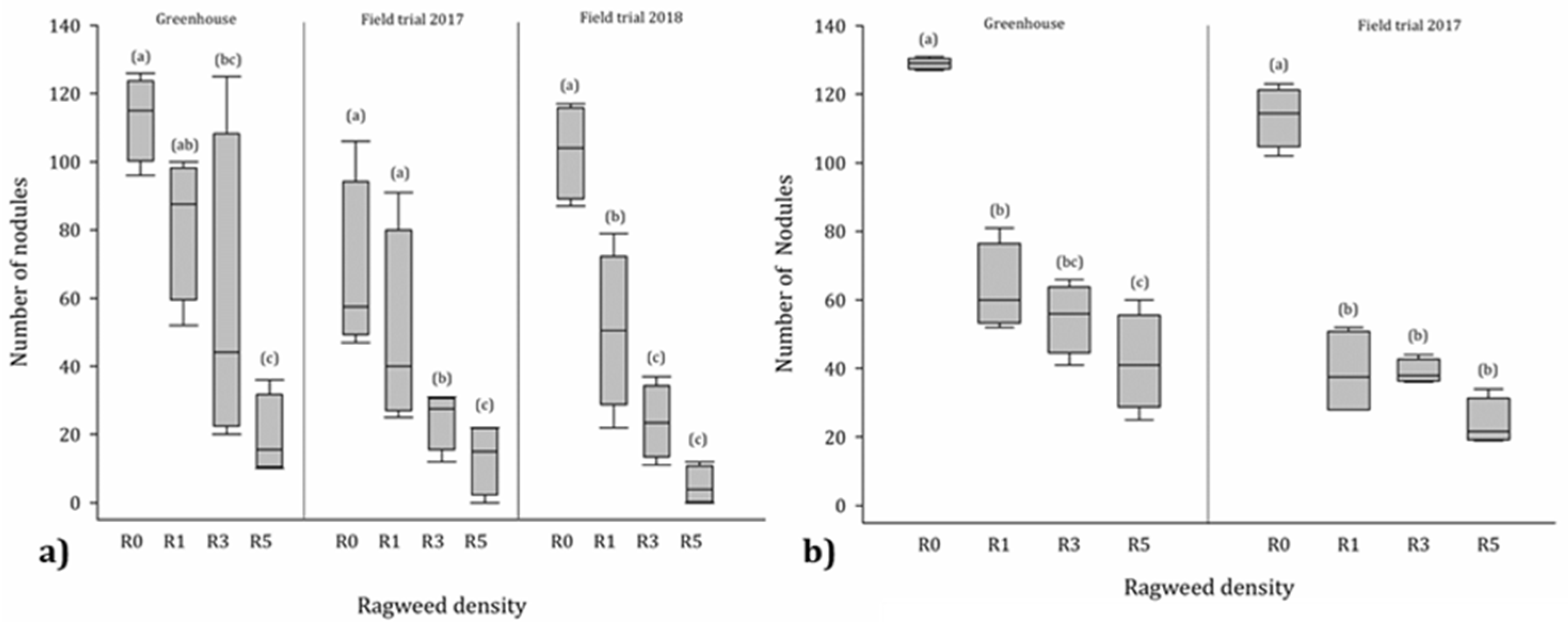
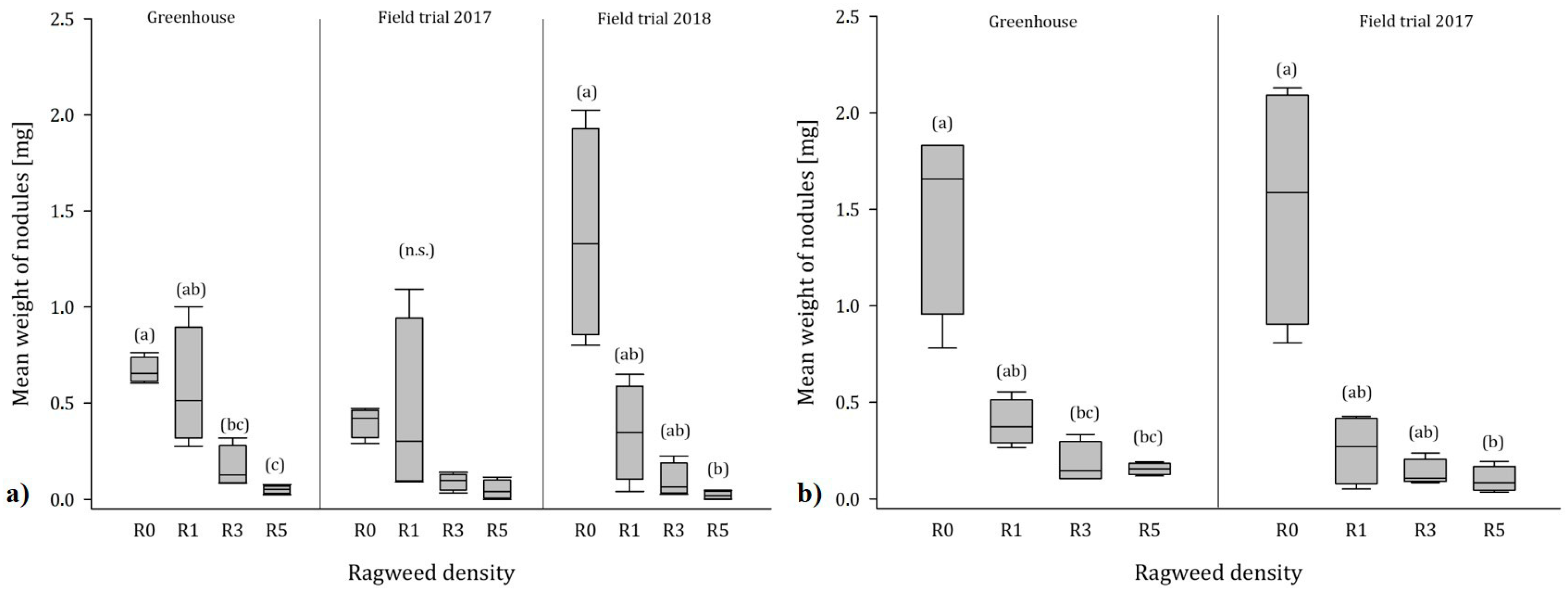
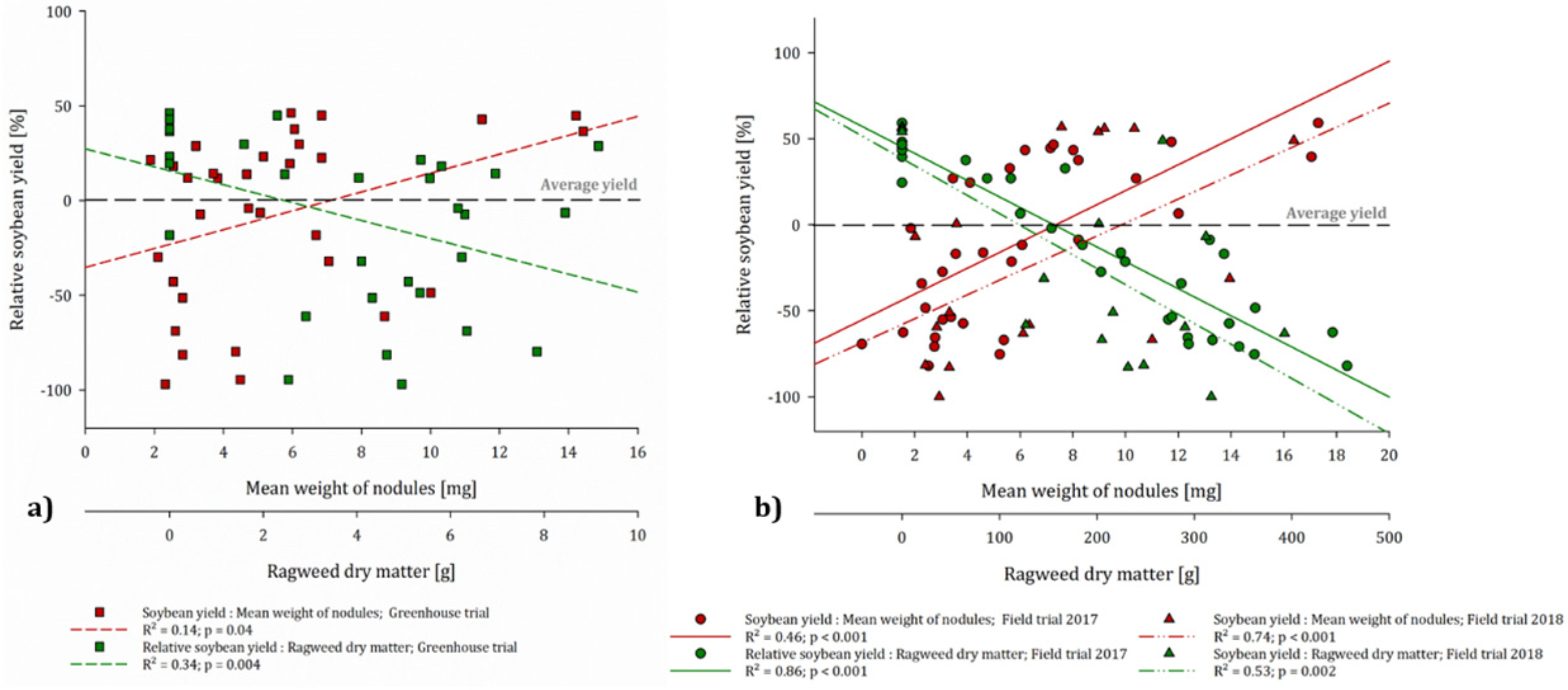
3.4. Nodules
3.5. Growth and Yield of Runner Bean
4. Discussion
4.1. Growth Performance of Ragweed
4.2. Soybean Yield
4.3. Influence of Ragweed on Nodulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pagano, M.C.; Miransari, M. The importance of soybean production worldwide. In Abiotic and Biotic Stresses in Soybean Production, 1st ed.; Miransari, M., Ed.; Elsevier Academic Press: Cambridge, UK, 2016; pp. 1–24. [Google Scholar]
- Zollitsch, W.; Wetscherek, W.; Lettner, F. Use of differently processed full-fat soybeans in a diet for pig fattening. Anim. Feed Sci. Technol. 1993, 41, 237–246. [Google Scholar] [CrossRef]
- Vollmann, J.; Fritz, C.N.; Wagentristl, H.; Ruckenbauer, P. Environmental and genetic variation of soybean seed protein content under Central European growing conditions. J. Sci. Food Agric. 2000, 80, 1300–1306. [Google Scholar] [CrossRef]
- Watkinson, A.R.; Freckleton, R.P.; Robinson, R.A.; Sutherland, W.J. Predictions of biodiversity response to genetically modified herbicide-tolerant crops. Science 2000, 289, 1554–1557. [Google Scholar] [CrossRef]
- Sandermann, H. Plant biotechnology: Ecological case studies on herbicide resistance. Trends Plant Sci. 2006, 11, 324–328. [Google Scholar] [CrossRef]
- Owen, M.D.K.; Pedersen, P.; De Bruin, J.L.; Lux, S.J.; Franzenburg, D.; Grossnickle, D. Comparison of genetically modified and non-genetically modified soybean cultivars and weed management systems. Crop Sci. 2010, 50, 2597–2604. [Google Scholar] [CrossRef]
- Johnson, B.; Loux, M.; Nordby, D.; Sprague, C.; Nice, G.; Westhoven, A.; Stachler, J. Biology and Management of Giant and Common Ragweed; U.S. Department of Agriculture: Washington, DC, USA, 2007; pp. 4–16.
- Karrer, G.; Milakovic, I.; Kropf, M.; Blöch, C.; Dlugosch, A.; Leitsch-Vitalos, M.; Hackl, G.; Follak, S.; Fertsak, S.; Schwab, M.; et al. Ausbreitungsbiologie und Management Einer Extrem Allergenen, Eingeschleppten Pflanze—Wege und Ursachen der Ausbreitung von Ragweed (Ambrosia artemisiifolia) Sowie Möglichkeiten Seiner Bekämpfung—Endbericht; Bundesministerium für Landwirtschaft, Regionen und Tourismus: Vienna, Austria, 2011; pp. 31–44.
- Richter, R.; Berger, U.E.; Dullinger, S.; Essl, F.; Leitner, M.; Smith, M.; Vogl, G. Spread of invasive ragweed: Climate change management and how to reduce allergy cost. J. Appl. Ecol. 2013, 50, 1422–1430. [Google Scholar] [CrossRef] [Green Version]
- Fumanal, B.; Chauvel, B.; Sabatier, A.; Bretagnolle, F. Variability and Cryptic Heteromorphism of Ambrosia artemisiifolia Seeds: What Consequences for its Invasion in France? Ann. Bot. 2007, 100, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Bullock, J. Assessing and Controlling the Spread and the Effect of Common Ragweed in Europe; Centre for Ecology & Hydrology: Wallingford, UK, 2010; pp. 26–46. [Google Scholar]
- Smith, M.; Cecchi, L.; Skjøth, C.A.; Karrer, G.; Šikoparija, B. Common ragweed: A threat to environmental health in Europe. Environ. Int. 2013, 61, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.M.; Wagentristl, H.; Karrer, G.; Winter, A.; Czerny, R.; Kaul, H.-P. Common Ragweed (Ambrosia artemisiifolia L.) Causes Severe Yield Losses in Different Soybean Varieties by Reducing the Infection Potential of Bradyrhizobium Japonicum; Tagungsband: 29. Deutsche Arbeitsbesprechung über Fragen der Unkrautbiologie und-bekämpfung; Julius Kühn-Institut: Braunschweig, Germany, 2020; pp. 72–79. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. Ecophysiology of secondary dormancy in seeds of Ambrosia artemisiifolia. Ecology 1980, 61, 475–480. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Temperature requirements for after-ripening in buried seeds of four summer annual weeds. Weed Res. 1987, 27, 385–389. [Google Scholar] [CrossRef]
- Coble, H.D.; Williams, F.M.; Ritter, R.L. Common ragweed (Ambrosia artemisiifolia) interference in soybean (Glycine max). Weed Sci. 1981, 29, 339–342. [Google Scholar] [CrossRef]
- Gibson, D.J.; Millar, K.M.; Delong, M.; Connolly, J.; Kirwan, L.; Wood, A.J.; Young, B.G. The weed community affects yield and quality of soybean (Glycine max Merr.). J. Sci. Food Agric. 2007, 88, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Brückner, D.; Lepossa, A.; Herpai, Z. Inhibitory effect of ragweed (Ambrosia artemisiifolia L.)-inflorescense extract on the germination of Amaranthus hypochondriacus L. and growth of two soil algae. Chemosphere 2003, 51, 515–519. [Google Scholar] [CrossRef]
- Wang, P.; Liang, W.J.; Kong, C.H.; Jiang, Y. Allelopathic potentials of volatile allelochemicals of Ambrosia trifida L. on other plants. Allelopath. J. 2005, 15, 131–136. [Google Scholar]
- Pajević, S.; Borišev, M.; Orčić, D.; Boža, P.; Nikolić, N. Photosynthetic and biochemical characteristics of invasive species (Ambrosia artemisiifolia L., Ambrosia trifida L. and Iva xanthifolia Nutt.) depending on soil humidity and phenological phase. Russ. J. Ecol. 2010, 41, 498–505. [Google Scholar] [CrossRef]
- Vidotto, F.; Tesio, F.; Ferrero, A. Allelopathic effects of Ambrosia artemisiifolia L. in the invasion process. Crop Prot. 2013, 54, 161–167. [Google Scholar] [CrossRef]
- Mutch, D.R.; Martin, T.E.; Kolola, K.R. Red clover (Trifolium pratense) suppression of common ragweed (Ambrosia artemisiifolia) in winter wheat (Triticum aestivum). Weed Technol. 2003, 17, 181–185. [Google Scholar] [CrossRef]
- Bonea, D.; Bonciu, E.; Niculescu, M.; Olaru, A.L. The allelopathic, cytotoxic and genotoxic effect of Ambrosia artemisiifolia on the germination and root meristeme of Zea mays. Caryologia 2018, 71, 24–48. [Google Scholar] [CrossRef]
- Molinaro, F.; Mozzetti, C.; Ferrero, A.; Tabasso, S.; Negra, M. Bioherbicidal activity of a germacranolide sesquiterpene dilactone from Ambrosia artemisiifolia L. J. Environ. Sci. Health B 2016, 51, 847–852. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Green-Tracewicz, E.; Page, E.R.; Swanton, C.J. Light quality and the critical period for weed control in soybean. Weed Sci. 2012, 60, 86–91. [Google Scholar] [CrossRef]
- Gal, J.; Afifi, M.; Lee, E.; Lewis, L.; Swanton, C.J. Detection of neighbouring weeds alter soybean seeling roots and nodulation. Weed Sci. 2015, 63, 888–900. [Google Scholar] [CrossRef]
- Devine, T.E.; Kuykendall, L.D. Host genetic control of symbiosis in soybean (Glycine max L.). Plant Soil 1996, 186, 173–187. [Google Scholar] [CrossRef]
- Blumenthal, J.M.; Russelle, M.P.; Vance, C.P. Localized and internal effect of nitrate on symbiotic dinitrogen fixation. Physiol. Plant. 1997, 101, 59–66. [Google Scholar] [CrossRef]
- Kiers, E.T.; Hutton, M.G.; Denison, R.F. Human selection and the relaxation of legume defences against ineffective rhizobia. Proc. R Soc. B 2014, 274, 3119–3126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sessitsch, A.; Ramirez-Saas, H.; Hardarson, G.; Akkermans, A.D.; de Vos, M. Classification of an Austrian rhizobia and the Mexican isolate FL27 obtained from Phaseolus vulgaris L. as Rhizobium gallicum. Int. J. Syst. Bacteriol. 1997, 47, 1097–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, R.; Causton, D.R.; Shipley, B.; Askew, A.P. A modern tool for classical plant growth analysis. Ann. Bot. 2002, 90, 485–488. [Google Scholar] [CrossRef] [Green Version]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of Akaike Information Criterion and Bayesian Approaches over Likelihood Ratio Tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Burnham, K.P.; Anderson, D.R. Model Selection and Inference: A Practical Information-Theoretic Approach, 1st ed.; Springer: New York, NY, USA, 2002; pp. 192–196. [Google Scholar]
- Bussan, A.J.; Burnside, O.C.; Orf, J.H.; Ristau, E.A.; Puettmann, K.J. Field evaluation of soybean (Glycine max) genotypes for weed competitiveness. Weed Sci. 1997, 45, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Bensch, C.N.; Horak, M.J.; Peterson, D. Intereference of redroot pigweed (Amaranthus retroflexus), Palmer amaranth (A. palmeri), and common waterhemp (A. rudis) in soybean. Weed Sci. 2003, 51, 37–43. [Google Scholar] [CrossRef]
- Zimdahl, R.L. Weed-Crop Competition: A Review, 2nd ed.; Blackwell Publishing: Oxford, UK, 2007. [Google Scholar]
- Monks, D.W.; Oliver, L.R. Interactions between soybean (Glycine max) cultivars and selected weeds. Weed Sci. 1988, 36, 770–774. [Google Scholar] [CrossRef]
- Essl, F.; Biró, K.; Brandes, D.; Broennimann, O.; Bullock, J.M.; Chapman, D.S.; Chauvel, B.; Dullinger, S.; Fumanal, B.; Guisaon, A.; et al. Biological Flora of the British Isles: Ambrosia artemisiifolia. J. Ecol. 2015, 103, 1069–1098. [Google Scholar] [CrossRef] [Green Version]
- Barrentine, W.L. Common cocklebur competition in soybeans. Weed Sci 1974, 22, 600–603. [Google Scholar] [CrossRef]
- Datta, A.; Ullah, H.; Tursun, N.; Pornprom, T.; Knezevic, S.Z.; Chauhan, B.S. Managing weeds using crop competition in soybean [Glycine max (L.) Merr.]. J. Crop Prot. 2017, 95, 60–68. [Google Scholar] [CrossRef]
- Berti, A.; Dunan, C.; Sattin, M.; Zanin, G.; Westra, P. A new approach to determine when to control weeds. Weed Sci. 1996, 44, 496–503. [Google Scholar] [CrossRef]
- Baysinger, J.A.; Sims, B.D. Giant ragweed (Ambrosia trifida) interference in soybeans (Glycine max). Weed Sci. 1991, 39, 358–362. [Google Scholar] [CrossRef]
- Mosier, D.G.; Oliver, L.R. Soybean (Glycine max) interference on common cocklebur (Xanthium strumarium) and entireleaf morningglory (Ipomoea hederacea var. integruscula). Weed Sci. 1995, 43, 402–409. [Google Scholar] [CrossRef]
- Weston, L.A.; Putnam, A.R. Inhibition of growth, nodulatoin, and nitrogen fixation of legumes by Quackgrass. Crop Sci. 1985, 25, 561–565. [Google Scholar] [CrossRef]
- Puppo, A.; Pauly, N.; Boscari, A.; Mandon, K.; Brouquisse, R. Hydrogen peroxide and nitric oxide: Key regulators of the Legume-Rhizobium and mycorrhizal symbiosis. Antioxid. Redox Signal. 2013, 18, 2202–2219. [Google Scholar] [CrossRef] [PubMed]
- Pierik, R.; Mommer, L.; Voesenek, L.A.C.J. Molecular mechanisms of plant competition: Neighbour detection and response strategies. Funct. Ecol. 2013, 27, 841–853. [Google Scholar] [CrossRef]
- Swamy, P.M.; Smith, B.N. Role of abscisic acid in plant stress tolerance. Curr. Sci. 1999, 76, 1220–1227. [Google Scholar]
- Finkelstein, R.R.; Rock, C.D. Abscisic acid biosynthesis and repsonse. In The Arabidopsis Book/American Society of Plant Biologists; ASPB Member Center: Rockville, MD, USA, 2002; p. e0058. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, A.; Akune, M.; Kogiso, M.; Imagama, Y.; Osuki, K.-I.; Uchiumi, T.; Higashi, S.; Han, S.-Y.; Yoshida, S.; Asami, T.; et al. Control of nodule number by the phytohormone abscisic acid in the roots of two leguminous species. Plant Cell Physiol. 2004, 45, 914–922. [Google Scholar] [CrossRef] [Green Version]
- Farooq, U.Z.M.A.; Bano, A. Effect of abscisic acid and chlorochloine chloride on nodulation and biochemical content of Vigna radiata L. under water stress. Pak. J. Bot. 2006, 38, 1511–1518. [Google Scholar]
- Cho, M.J.; Harper, J.E. Effect if abscisic acid application on root isoflavonoid concentration and nodulation of wild-type and nodulation-mutant soybean plants. Plant Soil 1993, 153, 145–149. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, C.; Yang, J.; Yu, N.; Wang, E. Hormone modulation of legume-rhizobial symbiosis. J. Integr. Plant Biol. 2018, 60, 632–648. [Google Scholar] [CrossRef] [PubMed]
- Long, S.R. Genes and signals in the rhizobium-legume symbiosis. Plant Physiol. 2001, 125, 69–72. [Google Scholar] [CrossRef] [Green Version]
- Makarova, L.E.; Latysheva, S.E.; Putilina, T.E. The effect of the phenolic compounds exuded by pea roots in darkness on the reproduction of Rhizobium. Appl. Biochem. Microbiol. 2007, 43, 429–434. [Google Scholar] [CrossRef]
- Makarova, L.E.; Smirnov, V.I.; Klyba, L.V.; Petrova, I.G.; Dudareva, L.V. Role of allelopathic compounds in the regulation and development of legume-rhizobial symbiosis. Appl. Biochem. Microbiol. 2012, 48, 355–362. [Google Scholar] [CrossRef]
- Wardle, D.A.; Nicholson, K.S.; Ahmed, M.; Rahman, A. Interference effects of the invasive plant Carduus nutans L. against the nitrogen fixation ability of Trifolium repens L. Plant Soil 1994, 163, 287–297. [Google Scholar] [CrossRef]
- Batish, D.R.; Levanya, K.; Singh, H.P.; Kohli, R.K. Phenolic allelochemicals released by Chenopodium murale affect the growth, nodulation and macromolecule content in chickpea and pea. Plant Growth Regul. 2007, 51, 119–128. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy—An Overview. In Chemically Mediated Interactions between Plants and Other Organisms; Cooper-Driver, G.A., Swain, T., Conn, E.E., Eds.; Springer: Boston, MA, USA, 1984; Volume 19. [Google Scholar] [CrossRef]
- Mizutani, J. Selected Allelochemicals. Crit. Rev. Plant Sci. 1999, 18, 653–671. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Dates | Greenhouse Trial 2018 | DAS | BBCH | Field Trial 2017 | DAS | BBCH | Field Trial 2018 | DAS | BBCH |
|---|---|---|---|---|---|---|---|---|---|
| Seeding | 13 February | 21 | 16–18 | 23 May | 17 May | ||||
| T1 | 6 March | 39 | 21–29 | 12 June | 20 | 21–29 | 6 June | 20 | 15–25 |
| T2 | 14 April | 60 | 51–65 | 3 July | 41 | 51–55 | 26 June | 40 | 25–29 |
| T3 | 4 May | 80 | 69–79 | 27 July | 65 | 59–65 | 20 July | 64 | 51–59 |
| T4 | 1 June | 108 | 96–99 | 21 August | 90 | 69–79 | 16 August | 91 | 65–79 |
| T5 | 22 September | 122 | 93–99 | 18 September | 124 | 96–99 | |||
| runner bean: | 19 October | 155 | 89–97 | ||||||
| Soybean | Ragweed | ||||||
|---|---|---|---|---|---|---|---|
| T1–T2 | T2–T3 | T3–T4 | T4–T5 | T2–T3 | T3–T4 | T4–T5 | |
| Greenhouse trial | |||||||
| Cultivar Albenga | |||||||
| R0 | 0.21 ± 0.01 | 0.47 ± 0.13 | 0.02 ± 0.20 | 0.01 ± 0.01 | n.a. | n.a. | n.a. |
| R1 | 0.18 ± 0.02 | 0.31 ± 0.14 | 0.07 ± 0.81 | 0.04 ± 0.34 | 0.02 ± 0.01 | 0.07 ± 0.03 | −0.01 ± 0.16 |
| R3 | 0.17 ± 0.02 | −0.04 ± 0.05 | 0.10 ± 0.18 | 0.15 ± 0.16 | 0.01 ± 0.00 | 0.05 ± 0.00 | −0.01 ± 0.01 |
| R5 | 0.14 ± 0.03 | 0.06 ± 0.04 | 0.01 ± 0.06 | 0.12 ± 0.07 | 0.01 ± 0.00 | 0.02 ± 0.00 | −0.00 ± 0.02 |
| Cultivar Mentor | |||||||
| R0 | 0.23 ± 0.05 | 0.51 ± 0.06 | 0.54 ± 0.25 | 0.01 ± 0.06 | n.a. | n.a. | n.a. |
| R1 | 0.20 ± 0.02 | 0.60 ± 0.07 | 0.28 ± 0.51 | −0.05 ± 0.14 | 0.01 ± 0.01 | 0.04 ± 0.00 | −0.05 ± 0.03 |
| R3 | 0.13 ± 0.03 | 0.49 ± 0.05 | 0.03 ± 0.32 | −0.00 ± 0.04 | 0.01 ± 0.00 | 0.04 ± 0.00 | 0.00 ± 0.03 |
| R5 | 0.09 ± 0.06 | 0.61 ± 0.04 | 0.02 ± 0.57 | 0.21 ± 0.24 | 0.01 ± 0.01 | 0.01 ± 0.01 | 0.00 ± 0.01 |
| R | *** | *** | n.s. | n.s. | *** | n.s. | n.s. |
| V | *** | ** | n.s. | n.s. | *** | n.s. | n.s. |
| R * V | * | *** | n.s. | n.s. | n.s. | n.s. | n.s. |
| Field trial 2017 | |||||||
| Cultivar Albenga | |||||||
| R0 | 0.45 ± 0.24 | 0.09 ± 0.20 | 0.25 ± 0.60 | 0.12 ± 0.53 | n.a. | n.a. | n.a. |
| R1 | 0.40 ± 0.21 | 0.25 ± 0.80 | 0.15 ± 1.07 | −0.10 ± 0.38 | 2.79 ± 4.01 | 1.55 ± 2.11 | 0.34 ± 0.66 |
| R3 | 0.42 ± 0.10 | 0.32 ± 0.38 | −0.53 ± 0.50 | 0.05 ± 0.02 | 1.13 ± 1.42 | 0.56 ± 0.64 | −0.16 ± 1.10 |
| R5 | 0.23 ± 0.27 | −0.01 ± 0.15 | 0.15 ± 0.23 | −0.11 ± 0.19 | 0.40 ± 0.47 | 1.93 ± 2.46 | −0.13 ± 0.18 |
| Cultivar Mentor | |||||||
| R0 | 0.31 ± 0.05 | 0.78 ± 0.17 | 0.38 ± 0.15 | 0.13± 0.28 | n.a. | n.a. | n.a. |
| R1 | 0.17 ± 0.13 | 0.65 ± 0.47 | 0.12 ± 0.18 | 0.05 ± 0.24 | 3.26 ± 1.84 | 1.65 ± 1.74 | 2.11 ± 0.93 |
| R3 | 0.22 ± 0.14 | 0.45 ± 0.26 | 0.01 ± 0.20 | −0.06 ± 0.32 | 1.30 ± 1.01 | 1.05 ± 0.79 | 0.47 ± 1.26 |
| R5 | 0.18 ± 0.16 | 0.23 ± 0.20 | 0.07 ± 0.30 | 0.03 ± 0.16 | 1.00 ± 1.52 | 0.79 ± 0.69 | 0.24 ± 1.42 |
| R | * | ** | n.s. | n.s. | n.s. | n.s. | n.s. |
| V | * | *** | n.s. | n.s. | ** | n.s. | n.s. |
| R * V | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
| Field trial 2018 | |||||||
| Cultivar Albenga | |||||||
| R0 | 0.32 ± 0.05 | 0.43 ± 0.5 | 0.30 ± 0.12 | 0.12 ± 0.20 | n.a. | n.a. | n.a. |
| R1 | 0.28 ± 0.06 | 0.45 ± 0.37 | 0.18 ± 0.13 | −0.03 ± 0.19 | 4.17 ± 2.90 | 1.84 ± 2.29 | 2.88 ± 4.29 |
| R3 | 0.22 ± 0.06 | 0.38 ± 0.17 | −0.05 ± 0.03 | −0.01 ± 0.16 | 2.86 ± 1.13 | 1.33 ± 0.61 | 0.14 ± 0.85 |
| R5 | 0.14 ± 0.10 | 0.11 ± 0.14 | −0.03 ± 0.04 | −0.04 ± 0.10 | 2.15 ± 1.81 | 2.55 ± 1.87 | 0.37 ± 1.60 |
| R | ** | *** | ** | n.s. | * | n.s. | n.s. |
| Parameter | Best Explanatory Model | AICc | ΔAICc | R2m (1) | R2c (2) |
|---|---|---|---|---|---|
| Soybean dry matter | Null model: trial | 616.5 | -- | ||
| ragweed dry matter × mean weight of nodules | 524.4 | 0.0 | 0.5 | 0.9 | |
| ragweed dry matter + mean weight of nodules | 527.3 | 2.9 | |||
| Soybean yield | Null model: trial | 540.7 | -- | ||
| ragweed dry matter × mean weight of nodules | 446.2 | 0.0 | 0.6 | 0.7 | |
| ragweed dry matter + mean weight of nodules | 452.7 | 6.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hall, R.M.; Urban, B.; Wagentristl, H.; Karrer, G.; Winter, A.; Czerny, R.; Kaul, H.-P. Common Ragweed (Ambrosia artemisiifolia L.) Causes Severe Yield Losses in Soybean and Impairs Bradyrhizobium japonicum Infection. Agronomy 2021, 11, 1616. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081616
Hall RM, Urban B, Wagentristl H, Karrer G, Winter A, Czerny R, Kaul H-P. Common Ragweed (Ambrosia artemisiifolia L.) Causes Severe Yield Losses in Soybean and Impairs Bradyrhizobium japonicum Infection. Agronomy. 2021; 11(8):1616. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081616
Chicago/Turabian StyleHall, Rea Maria, Bernhard Urban, Helmut Wagentristl, Gerhard Karrer, Anna Winter, Robert Czerny, and Hans-Peter Kaul. 2021. "Common Ragweed (Ambrosia artemisiifolia L.) Causes Severe Yield Losses in Soybean and Impairs Bradyrhizobium japonicum Infection" Agronomy 11, no. 8: 1616. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11081616