1. Introduction
Fruit berries, especially strawberry and wild strawberry (Rosaceae), belong to the most popular horticultural crops. They have delicious taste and aroma, high economic importance, low calorific value, and are of great interest for nutritionists and food technologists because they are often consumed in fresh form when the most bioactive compounds are still active and in the greatest amount [
1]. Poland is an important global producer of strawberries (47.9 thousand hectares), ranking second just after China (111 thousand hectares) [
2]. Octoploid cultivated strawberries (
Fragaria ×
ananassa Duch.) and diploid wild or wood strawberries (
Fragaria vesca L.) are consumed in high quantities (the yield of strawberry is the 7th largest in the world) [
3] and are thus a valuable source of ascorbic acid, potassium, fiber, simple sugars, as well as antioxidants, such as phenolics, anthocyanins, and other flavonoid compounds [
4,
5,
6]. Although yield is commonly the main focus of berries production, consumers demand more and more often fruits with higher phytochemical properties [
7]. Consumption of berries of the Rosaceae family is shown to lower the risk for chronic diseases like inflammation disorders, cancer, cardiovascular and obesity-related disorders, and stroke [
8]. Consequently, numerous studies on the cultivation, physiology, and health-promoting properties of (wild) strawberry are performed.
Two main categories of wild and cultivated strawberry exist: (1) short-day seasonal flowering genotypes which initiate flower buds either under short-day conditions (day length less than 14 h) or at a temperature less than 15 °C and produce only one flush of flowering and fruiting in the spring; and (2) long-day recurrent flowering or everbearing genotypes which, in addition to a spring flush, produce more or less continuous flowering and fruiting throughout the growing season. Although temperature modifies the photoperiodic responses of these plants, they are less sensitive to high temperatures than short-day plants [
9,
10].
Consequently, the cultivation of (wild) strawberry can be performed either outdoors or indoors. Due to the growing demand of developed countries for year-round fresh fruits, many producers are shifting from traditional open-field cultivation to greenhouses [
11,
12]. Therefore, information about the agronomic performance of (wild) strawberry cultivars in greenhouse conditions is widely required to help producers in choosing the best cultivars for this application [
7]. Soilless growing systems, such as gutter cultivation, have several advantages over traditional cultivation in the soil, which is limited due to the ergonomic difficulties of cultivating the plants on the ground surface and the prohibition of chemical fumigants for the control of phytopathogens [
13]. Protected cultivation has provided efficient ways to overcome potentially harmful outdoor conditions such as low and high solar radiation, extreme temperatures, wind, heavy rain, frost, and hail. Moreover, the greenhouse environment can be easily controlled with the required water vapor, CO
2 concentration, and airflow conditions [
14,
15]. Consequently, an extension of the harvesting period and fruit quality improvement are observed [
16]. The technology of soilless cultivation attracts producers due to easier fertility control, the possibility of increasing plant density and yield, a decrease in diseases and pests, and the increment in the product quantity and quality [
17]. Production of plants in closed fertigation systems and coconut fibre substrate is the current trend for the cultivation of (wild) strawberry. When cultivation is carried out on benches above ground level, the work is easier, the use of fumigant soil products is eliminated, and there is a reduction in the occurrence of leaf diseases and, consequently, in the application of pesticides [
13]. To prevent crops from suffering damage caused by the shortage of sunlight, the introduction of supplementary light into the cultivation practice is necessary. Various light sources—such as incandescent, fluorescent, metal halide, or high-pressure sodium lamps (HPS) or light-emitting diodes (LEDs)—were adopted as supplementary lights for the cultivation of strawberry plants [
18].
Environmental factors—such as light, water, CO
2, temperature, and nutrients—are important elements in the production of high-quality berries [
18]. The physiology of (wild) strawberry growth and flowering is complex because of the great variability of the plant behavior in response to climate conditions, and the genetic background of the cultivar [
1,
10]. Those factors directly affect the biosynthesis of essential metabolites in fruits and leaves [
4].
The health-promoting properties of fruits are a result of a unique composition of bioactive compounds, including polyphenols, which are characterized by a high antioxidant activity. The presence and quantity of polyphenols have been used as markers allowing the distinction between cultivars. Fruits may vary in terms of the composition of polyphenols depending on the cultivar, growing conditions, and ripeness stage. The dominant group of polyphenols in (wild) strawberry fruit are anthocyanins with pelargonidin-3-glucoside being the major one [
19].
Chlorophylls and carotenoids fulfill various vital functions in plants. In the chloroplast, chlorophylls are the predominant light-absorbing pigment in photosynthesis, and carotenoids are an important component required for light-harvesting and photoprotection [
20,
21]. A physiological performance, especially
PN and
Gs, are related to the leaf thickness and chlorophyll content as the increase in these parameters enable the plant to absorb more light, particularly in the range of photosynthetically active radiation (PAR), and convert light energy into chemical energy [
22,
23]. Besides the essential function as photoprotectants, carotenoids act as oxidative stress ‘sensor’ and ‘signal’ upon oxidation by reactive oxygen species (ROS) [
24]. Oxidative stress is an integral component of various stress conditions in plants, and this fact mainly determines the substantial overlap in physiological and molecular responses to biotic and abiotic environmental challenges [
25]. Moreover, derivatives of chlorophylls and carotenoids consumed with berry fruits are consistent with cancer prevention in humans including antioxidant and antimutagenic activity [
26,
27]. Photosynthesis is inextricably linked to the physiology of the entire plant through mutual controls. A metabolic signal network including information on the carbon and nitrogen status of tissues interacts with phytohormone signaling and redox signaling to control photosynthetic gene expression and leaf development [
28].
The availability of fresh fruit on the European market throughout the year is varied; i.e., in the autumn and winter season, fresh fruits of (wild) strawberries from local markets are often unavailable, or their taste is worse than that of fruits grown in the spring. Extending the harvest period would allow for a year-round consumption of fresh strawberries from domestic productions and an increase in the producers’ income [
29]. Only the development of greenhouse technologies, including gutter cultivation, gives a chance to lower production costs in colder climatic zones. Therefore, it is desirable to develop research on the physiological, biochemical, and biometrical activities of strawberry and wild strawberry in the autumn–winter gutter cultivation. The selection of the most suitable cultivars for this purpose is also needed.
Each strawberry and wild strawberry cultivar has its growth and yield characteristics [
15]. However, the response of many cultivars to microclimate conditions in gutter cultivation in greenhouses has not been yet determined. Therefore, the objective of this study was to determine the physiological, biochemical, and biometrical responses of two strawberry and two wild strawberry cultivars.
The research hypothesis assumed that among the various cultivars of these two plant species, it is possible to select those that have better biological and physiological attributes for greenhouse gutter cultivation in the autumn–winter season in Poland.
2. Materials and Methods
2.1. Plant Material, Cultivation Conditions, and Evaluation of Crop Productivity
Potted plants of two cultivars of strawberry (everbearing ‘Ostara’ and day-neutral ‘San Andreas’) and two cultivars of wild strawberry (‘Baron von Solemacher’ and ‘Regina’) were used in the study (
Figure 1). The experiment was set in a greenhouse of the Laboratory of Ornamental Plants and Vegetable Crops at the Faculty of Agriculture and Biotechnology of the UTP University of Science and Technology in Bydgoszcz, Poland, (53°07′12.0″ N 18°00′29.4″ E) in the period from 17 September 2020 to 4 February 2021. The experiment was set in two gutters (
Figure 1A), with each gutter used for the separate cultivation of strawberry and wild strawberry. Eight plants of each cultivar studied were planted every 12.5 cm on coconut mats (1 m long). Each cultivar was grown on five mats, with a total of 40 individual plants available. An alternating distribution of cultivars in the gutter was used. The coconut mats were pre-rinsed with water through a fertigation system to remove excess salt. Both the substrate and the filtrate were subjected to chemical analysis of impurities. In the period from 14 October 2020 until the end of the experiment, the plants were additionally irradiated with HPS lamps (LeuTech 600W, Thörigen, Switzerland) for two to six hours from 2 p.m. The mean photosynthetic photon flux density (PPFD) was 100 μmol·m
−2·s
−1. The temperature in the greenhouse was 18.5 °C day/15.5 °C night. Plants were fertigated 2–3 times a week delivering a total of 230 mL medium per two plants per day. Terms of fertigation doses were established according to tensiometer indications. Fertigation was started when the water potential in the soil was −40 kPa. The nutrient solution had an EC of 1.3 mS cm
−1 and a pH of 5.5 during the vegetative growth of plants. When the plants started flowering, the EC was raised to 2.2 mS·cm
−1 at a pH of 5.5–6.0.
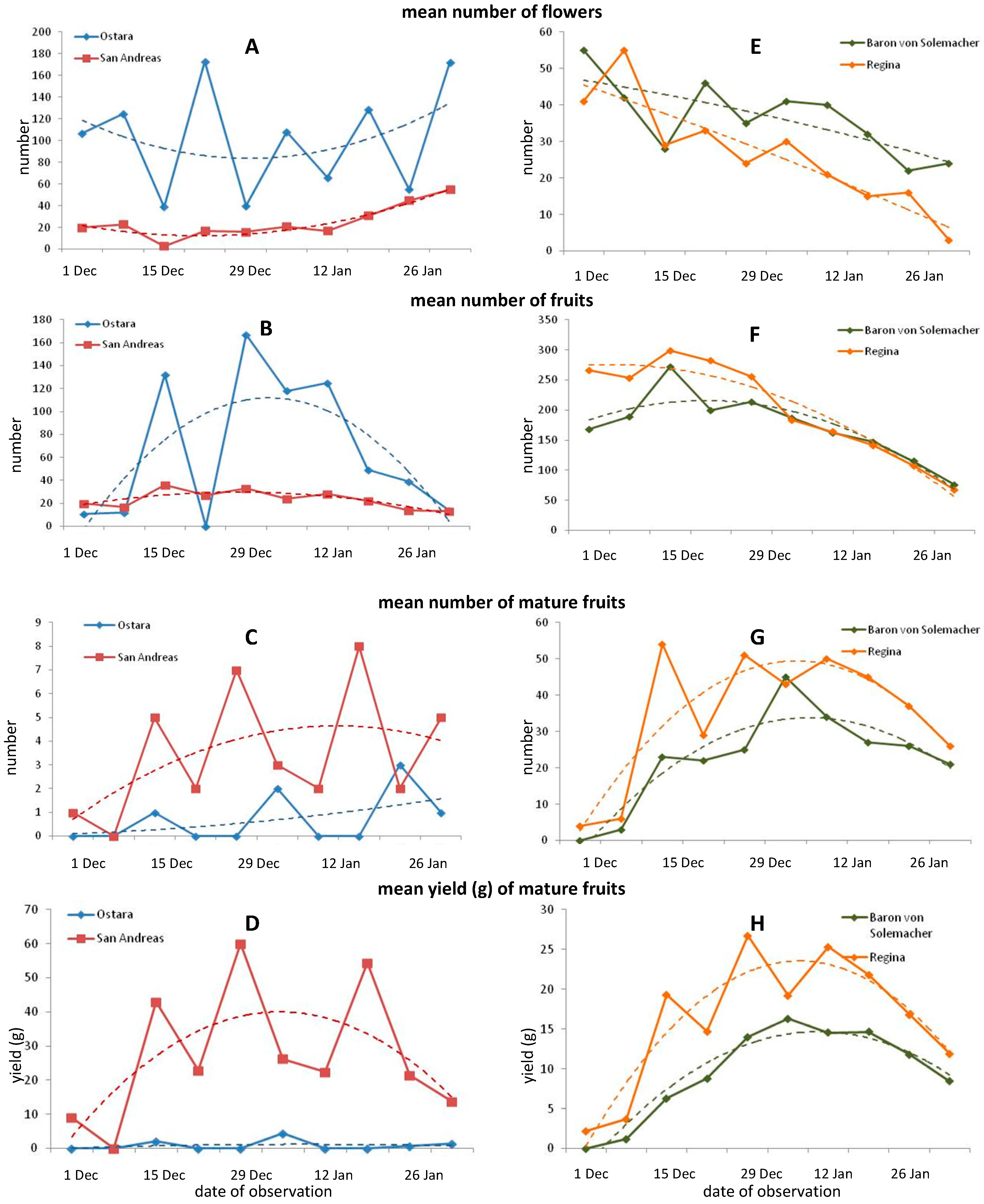
Starting from 1 December, weekly observations and measurements were made regarding the number of produced shoots, flowers, and fruits. Based on weekly measurements of these parameters, graphs were created and trend lines were added according to the polynomial algorithm. The fresh weight (FW) of mature fruit was also considered.
At the end of the experiment, leaves were collected from rosettes and scanned (Epson STD4800 scanner, USA). Next, the obtained pictures were analyzed to measure the mean leaf area (cm2), maximal leaf horizontal width (cm), maximal leaf vertical length (cm), and leaf perimeter (cm) using the imaging software WinFOLIATM (Reagen Instruments, Quebec, QC, Canada). Fresh weight (FW) and dry weight (DW) (g) of leaves were also determined.
2.2. Gas-Exchange Measurements
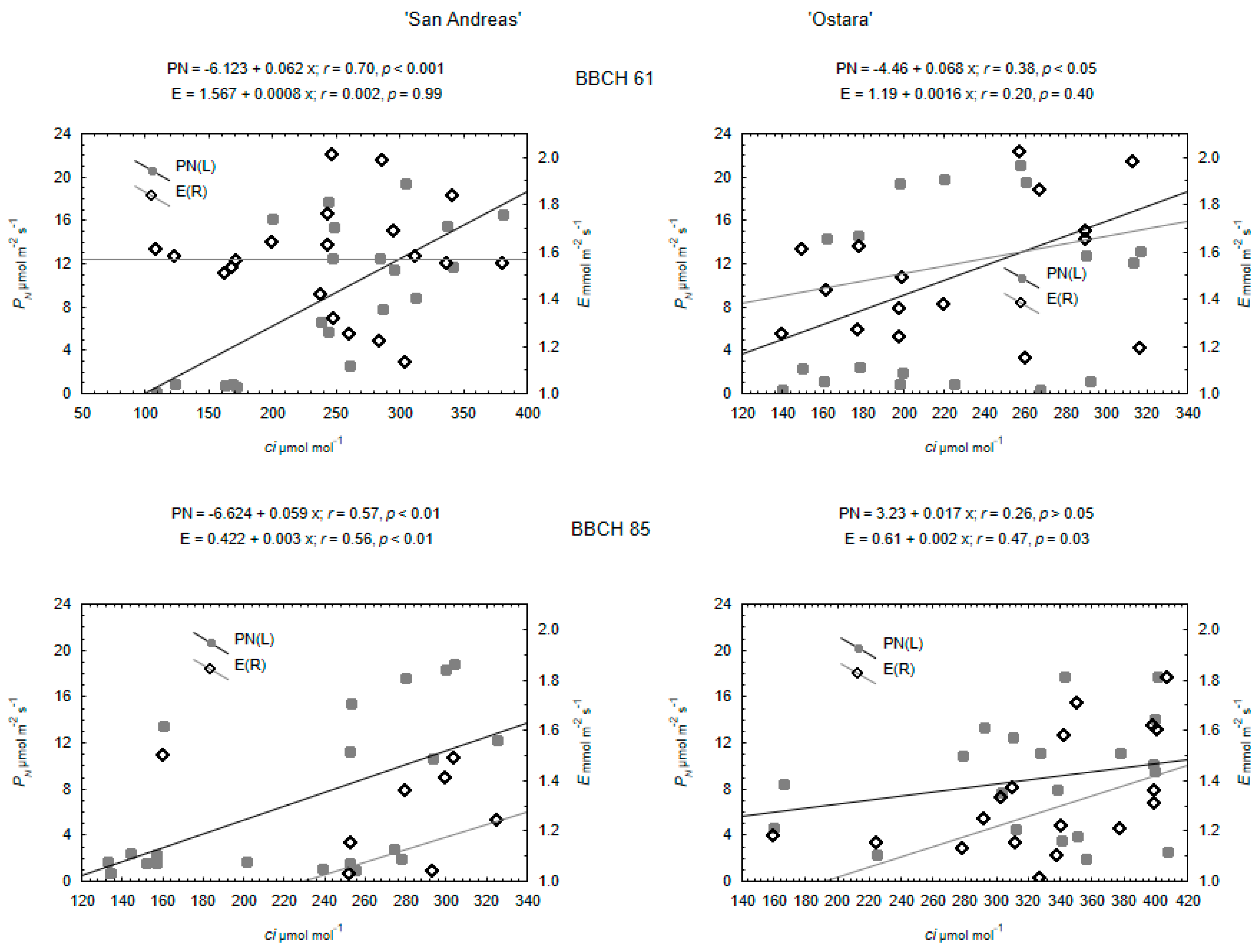
The main gas-exchange parameters, net photosynthetic rate (PN), transpiration rate (E), stomatal conductance (gs), and intercellular carbon dioxide (CO2) concentration (Ci) were measured at the center of the leaflet area, the leaves were fitted into a 6.25 cm2 clamp-on Plant Leaf Chamber (PLC Broad with mixed Red/Blue LED array). Gas exchange measurements lasted 20 min. This was performed using a portable open infrared CO2 gas analyzer (LC-Pro+, ADC BioScientific Ltd., Hoddesdon, UK) between 10.00 a.m. and 2:00 p.m. The system allowed for an automated microclimate control in the PLC. The conditions were stable in PLC and amounted to light 600 μmol·m−2·s−1, CO2 concentration 360 ± 5 ppm; temperature 22 ± 1 °C, water vapor pressure 10 ± 1 mbar (relative humidity approximately 40%). The rate of airflow through the LCpro+ chamber was approximately 200 mL∙min−1. In the growth chamber where the gas-exchange parameters were measured, the oxygen concentration was ambient (21%). Each parameter was measured twice, at stages BBCH 61 (flowering stage) and BBCH 85 (mature fruit stage) at 20 individuals for both species and cultivars.
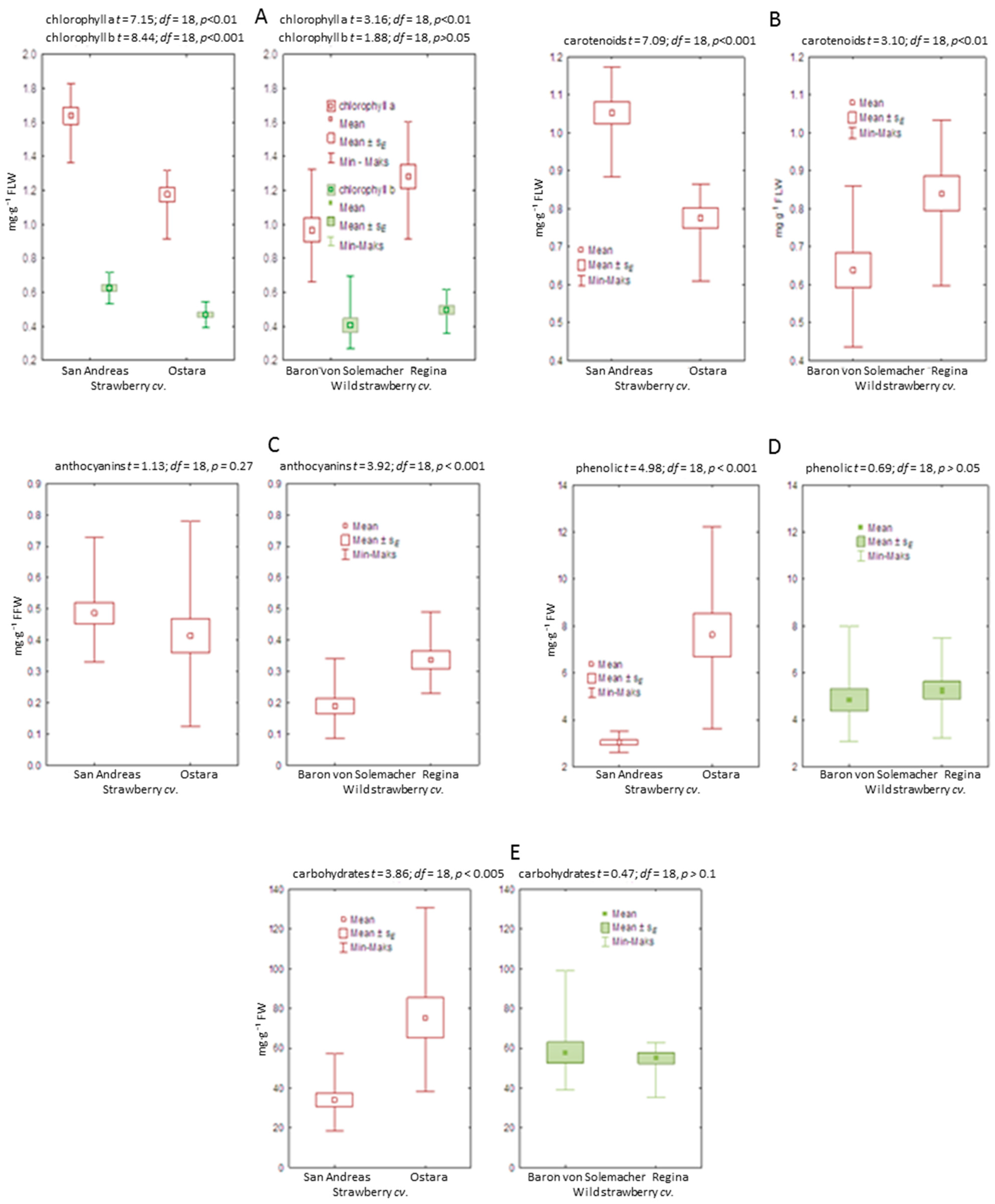
2.3. Biochemical Analysis of Leaves and Fruits
Carotenoids and anthocyanins were extracted from fresh leaves or fruits (100 or 200 mg of fresh tissue, respectively) according to the procedure described by Lema-Rumińska and Zalewska [
30]. The total contents of carotenoids and anthocyanins were calculated with the coefficient obtained from the Wettstein [
31] or Harborne [
32] equation, respectively. Chlorophyll
a and
b were extracted from fresh leaves according to Lichtenthaler and Buschmann [
33] using 100% acetone and 100 mg of fresh tissue samples. Carbohydrates were extracted from 100 mg fresh fruit samples with 2.5 M HCl. The quantification of total carbohydrate content was based on the anthrone test [
34]. The same extract was used to analyze the total phenolic content, according to the Folin-Ciocalteau procedure [
35].
The spectral analysis of extracts was performed in a UV-VIS spectrophotometer–Shimadzu UV 1601-PC (Kioto, Japan) and SmartSpec PlusTM (BioRad, Hercules, CA, USA) in a quartz cuvette (1 cm) at specific wavelengths (λmax): for carotenoids at 440 nm, for anthocyanins at 530 nm, for chlorophyll a and b at 645 and 662 nm, for carbohydrates at 620 nm, and for phenolics at 765 nm. The carbohydrate and phenolic contents were calculated using glucose and gallic acid as the calibration standards, respectively. The content of pigments, carbohydrates, and phenolics was calculated per gram of fresh weight. The biochemical analysis was performed in ten repetitions.
2.4. Statistical Analysis
Data obtained for all the physiological, biochemical, and biometrical characteristics of strawberry and wild strawberry were checked for normality using the Shapiro–Wilk test and the stability of variance with Levene’s test in the groups of cultivars. Verification of null hypotheses regarding the effects of growth stage and cultivars was carried out based on the two-way ANOVA, F test for p < 0.05. For the proven effects of the experimental factors on the physiological parameters in strawberry and wild strawberry, testing of the differences between the mean values was performed with the use of the HSD post-hoc Tukey’s test at the level of p < 0.05. The differences between cultivars in biochemical, and biometrical parameters were tested using the t-Student test, at p = 0.05. r-Pearson’s simple correlation coefficients and linear regressions were estimated between values of Ci and PN/E parameters for both species. The Statistica 13 (StatSoft, Cracow, Poland) software package was used for all the analyses.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}