
Effect of Nitrogen Fertilization and Fungicide Application at Heading on the Gluten Protein Composition and Rheological Quality of Wheat


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Treatments
- late-season N fertilization:
- ○
- Unfertilized control, with a total of 130 kg N ha−1 top-dressed applied as a granular ammonium nitrate (AN) fertilizer, split into 50 kg N ha−1 at tillering (GS23) and 80 kg N ha−1 at the beginning of stem elongation (GS32), without any late fertilization at the middle of heading (GS55);
- ○
- N application at heading (GS55), with the addition of 30 kg N ha−1 top-dressed, applied as AN, to the previously reported fertilization during the vegetative stages; the total N rate was 160 kg N ha−1.
- fungicide application:
- ○
- Untreated control;
- ○
- Heading application (GS55), with the application of a mixture of triazole fungicide prothioconazole + tebuconazole applied as 0.125 kg + 0.125 kg of the active ingredient (AI) ha−1 (Prosaro®, Bayer, Italy).
2.2. Crop Assessments
2.2.1. Grain Yield
2.2.2. Kernel Quality Traits and Rheological Properties
2.2.3. Gluten Protein Quantification
2.2.4. Gliadin Separation by Means of SDS-PAGE and Densitometric Analyses
2.2.5. Statistical Analysis
3. Results
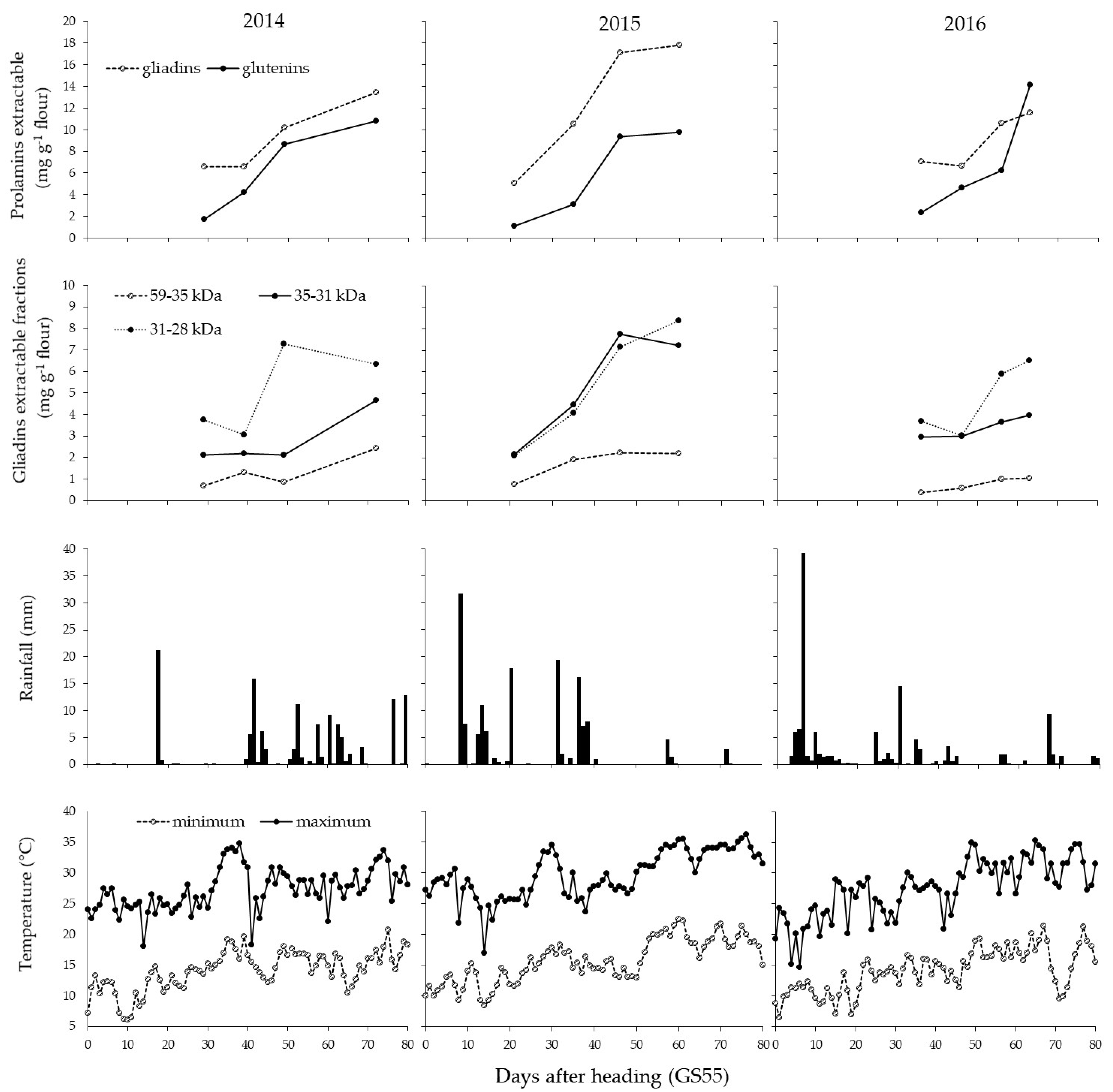
3.1. Weather Conditions
3.2. Grain Yield and Kernel Traits
3.3. Gluten Fractions and Alveographic Parameters of the Refined Flour at Harvest
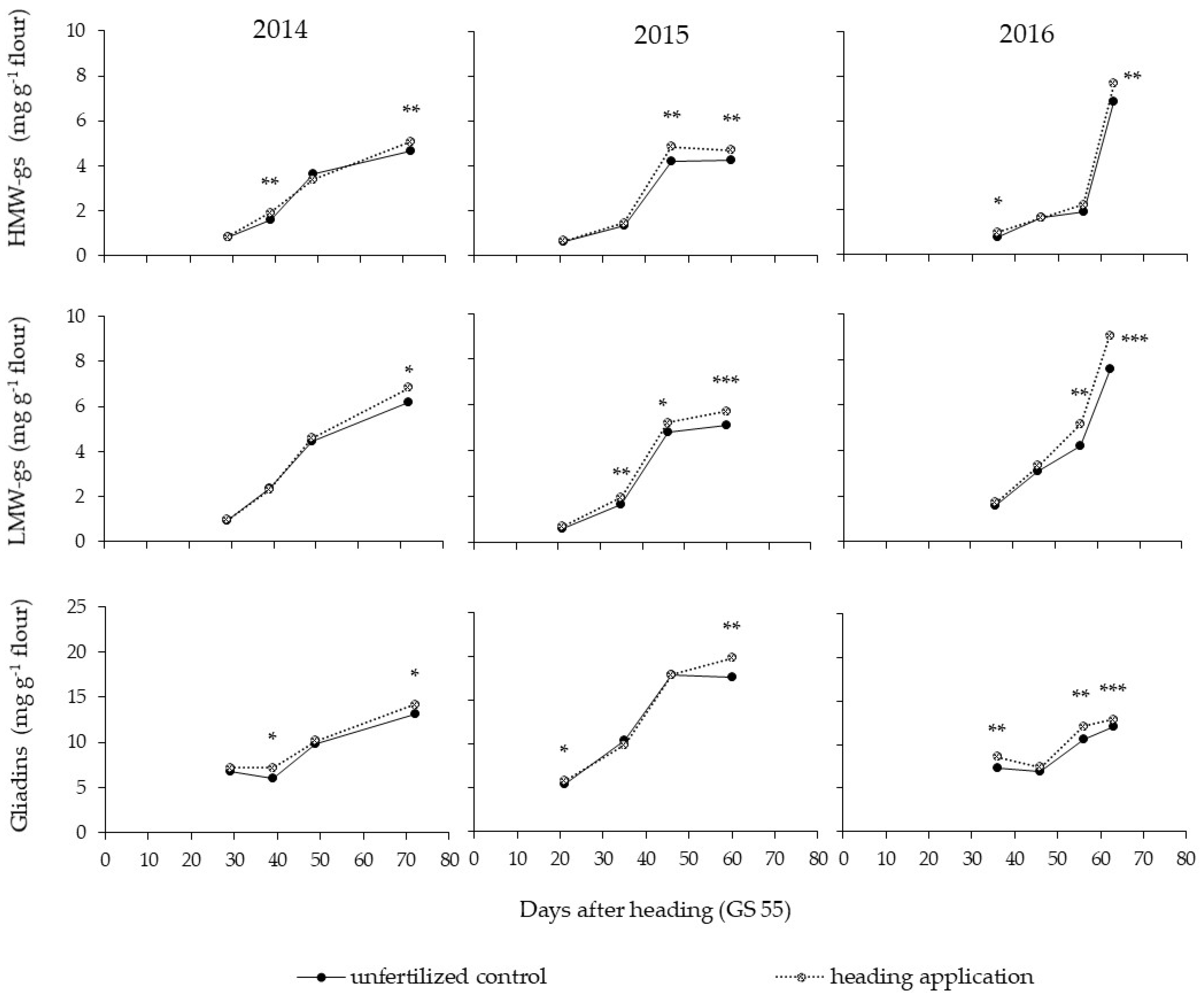
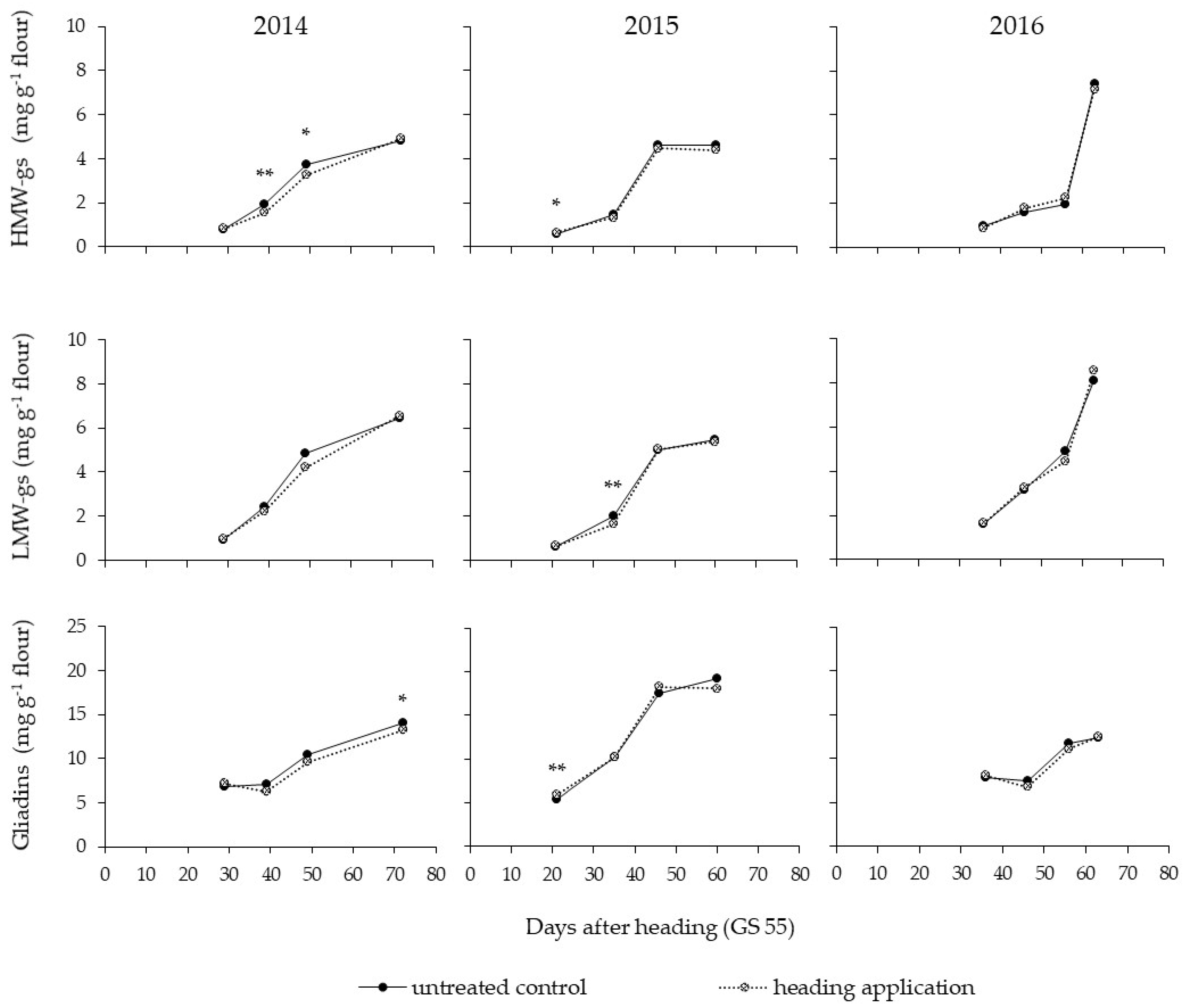
3.4. Accumulation of the Gluten Fractions during Ripening
4. Discussion
4.1. Effect of Late-Season N Fertilization
4.2. Effect of a Fungicide Application
4.3. Effect of the Year on Gluten Protein Accumulation during Ripening
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Ma, W. Wheat Gluten Protein and Its Impacts on Wheat Processing Quality. Front. Agric. Sci. Eng. 2019, 6, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R.; Halford, N.G. Cereal Seed Storage Proteins: Structures, Properties and Role in Grain Utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uthayakumaran, S.; Newberry, M.; Keentok, M.; Stoddard, F.L.; Bekes, F. Basic Rheology of Bread Dough with Modified Protein Content and Glutenin-to-Gliadin Ratios. Cereal Chem. 2000, 77, 744–749. [Google Scholar] [CrossRef]
- Denčić, S.; Mladenov, N.; Kobiljski, B. Effects of Genotype and Environment on Breadmaking Quality in Wheat. Int. J. Plant Prod. 2011, 5, 71–82. [Google Scholar]
- Blandino, M.; Vaccino, P.; Reyneri, A. Late-Season N Increases Improver Common and Durum Wheat Quality. Agron. J. 2015, 107, 680–690. [Google Scholar] [CrossRef] [Green Version]
- Blandino, M.; Visioli, G.; Marando, S.; Marti, A.; Reyneri, A. Impact of Late-Season N Fertilisation Strategies on the Gluten Content and Composition of High Protein Wheat Grown under Humid Mediterranean Conditions. J. Cereal Sci. 2020, 94, 102995. [Google Scholar] [CrossRef]
- Martre, P.; Porter, J.R.; Jamieson, P.D.; Triboï, E. Modeling Grain Nitrogen Accumulation and Protein Composition to Understand the Sink/Source Regulations of Nitrogen Remobilization for Wheat. Plant Physiol. 2003, 133, 1959–1967. [Google Scholar] [CrossRef] [Green Version]
- Fuertes-Mendizábal, T.; González-Torralba, J.; Arregui, L.M.; González-Murua, C.; González-Moro, M.B.; Estavillo, J.M. Ammonium as Sole N Source Improves Grain Quality in Wheat. J. Sci. Food Agric. 2013, 93, 2162–2171. [Google Scholar] [CrossRef]
- Foca, G.; Ulrici, A.; Corbellini, M.; Pagani, M.A.; Lucisano, M.; Franchini, G.C.; Tassi, L. Reproducibility of the Italian ISQ Method for Quality Classification of Bread Wheats: An Evaluation by Expert Assessors. J. Sci. Food Agric. 2007, 87, 839–846. [Google Scholar] [CrossRef]
- Fuertes-Mendizábal, T.; Estavillo, J.M.; Duñabeitia, M.K.; Huérfano, X.; Castellón, A.; González-Murua, C.; Aizpurua, A.; González-Moro, M.B. 15N Natural Abundance Evidences a Better Use of N Sources by Late Nitrogen Application in Bread Wheat. Front. Plant Sci. 2018, 9, 853. [Google Scholar] [CrossRef]
- Bogard, M.; Allard, V.; Brancourt-Hulmel, M.; Heumez, E.; Machet, J.-M.; Jeuffroy, M.-H.; Gate, P.; Martre, P.; Le Gouis, J. Deviation from the Grain Protein Concentration–Grain Yield Negative Relationship Is Highly Correlated to Post-Anthesis N Uptake in Winter Wheat. J. Exp. Bot. 2010, 61, 4303–4312. [Google Scholar] [CrossRef] [Green Version]
- Rossmann, A.; Scherf, K.A.; Rühl, G.; Greef, J.M.; Mühling, K.H. Effects of a Late N Fertiliser Dose on Storage Protein Composition and Bread Volume of Two Wheat Varieties Differing in Quality. J. Cereal Sci. 2020, 93, 102944. [Google Scholar] [CrossRef]
- Xue, C.; auf’m Erley, G.S.; Rücker, S.; Koehler, P.; Obenauf, U.; Mühling, K.H. Late Nitrogen Application Increased Protein Concentration but Not Baking Quality of Wheat. J. Plant Nutr. Soil Sci. 2016, 179, 591–601. [Google Scholar] [CrossRef]
- Rekowski, A.; Wimmer, M.A.; Henkelmann, G.; Zörb, C. Is a Change of Protein Composition after Late Application of Nitrogen Sufficient to Improve the Baking Quality of Winter Wheat? Agriculture 2019, 9, 101. [Google Scholar] [CrossRef] [Green Version]
- Triboï, E.; Martre, P.; Triboï-Blondel, A. Environmentally-induced Changes in Protein Composition in Developing Grains of Wheat Are Related to Changes in Total Protein Content. J. Exp. Bot. 2003, 54, 1731–1742. [Google Scholar] [CrossRef]
- Song, L.; Li, L.; Zhao, L.; Liu, Z.; Li, X. Effects of Nitrogen Application in the Wheat Booting Stage on Glutenin Polymerization and Structural–Thermal Properties of Gluten with Variations in HMW-GS at the Glu-D1 Locus. Foods 2020, 9, 353. [Google Scholar] [CrossRef]
- Castro, A.C.; Fleitas, M.C.; Schierenbeck, M.; Gerard, G.S.; Simón, M.R. Evaluation of Different Fungicides and Nitrogen Rates on Grain Yield and Bread-Making Quality in Wheat Affected by Septoria tritici Blotch and Yellow Spot. J. Cereal Sci. 2018, 83, 49–57. [Google Scholar] [CrossRef]
- Blandino, M.; Reyneri, A. Effect of Fungicide and Foliar Fertilizer Application to Winter Wheat at Anthesis on Flag Leaf Senescence, Grain Yield, Flour Bread-Making Quality and DON Contamination. Eur. J. Agron. 2009, 30, 275–282. [Google Scholar] [CrossRef]
- Gaju, O.; Allard, V.; Martre, P.; Le Gouis, J.; Moreau, D.; Bogard, M.; Hubbart, S.; Foulkes, M.J. Nitrogen Partitioning and Remobilization in Relation to Leaf Senescence, Grain Yield and Grain Nitrogen Concentration in Wheat Cultivars. Field Crops Res. 2014, 155, 213–223. [Google Scholar] [CrossRef]
- Blandino, M.; Pascale, M.; Haidukowski, M.; Reyneri, A. Influence of Agronomic Conditions on the Efficacy of Different Fungicides Applied to Wheat at Heading: Effect on Flag Leaf Senescence, Fusarium Head Blight Attack, Grain Yield and Deoxynivalenol Contamination. Ital. J. Agron. 2011, 6, e32. [Google Scholar] [CrossRef]
- Gooding, M.J.; Gregory, P.J.; Ford, K.E.; Pepler, S. Fungicide and Cultivar Affect Post-Anthesis Patterns of Nitrogen Uptake, Remobilization and Utilization Efficiency in Wheat. J. Agric. Sci. 2005, 143, 503–518. [Google Scholar] [CrossRef]
- Ruske, R.E.; Gooding, M.J.; Jones, S.A. The Effects of Triazole and Strobilurin Fungicide Programmes on Nitrogen Uptake, Partitioning, Remobilization and Grain N Accumulation in Winter Wheat Cultivars. J. Agric. Sci. 2003, 140, 395–407. [Google Scholar] [CrossRef]
- Motta-Romero, H.; Niyongira, F.; Boehm, J.D.B., Jr.; Rose, D.J. Effects of Foliar Fungicide on Yield, Micronutrients, and Cadmium in Grains from Historical and Modern Hard Winter Wheat Genotypes. PLoS ONE 2021, 16, e0247809. [Google Scholar] [CrossRef] [PubMed]
- Dimmock, J.P.R.E.; Gooding, M.J. The Influence of Foliar Diseases, and Their Control by Fungicides, on the Protein Concentration in Wheat Grain: A Review. J. Agric. Sci. 2002, 138, 349–366. [Google Scholar] [CrossRef]
- Matzen, N.; Ravn Jørgensen, J.; Holst, N.; Nistrup Jørgensen, L. Grain Quality in Wheat—Impact of Disease Management. Eur. J. Agron. 2019, 103, 152–164. [Google Scholar] [CrossRef]
- Fleitas, M.C.; Schierenbeck, M.; Gerard, G.S.; Dietz, J.I.; Golik, S.I.; Campos, P.E.; Simón, M.R. How Leaf Rust Disease and Its Control with Fungicides Affect Dough Properties, Gluten Quality and Loaf Volume under Different N Rates in Wheat. J. Cereal Sci. 2018, 80, 119–127. [Google Scholar] [CrossRef]
- Malik, A.H.; Prieto-Linde, M.L.; Kuktaite, R.; Andersson, A.; Johansson, E. Individual and Interactive Effects of Cultivar Maturation Time, Nitrogen Regime and Temperature Level on Accumulation of Wheat Grain Proteins. J. Sci. Food Agric. 2011, 91, 2192–2200. [Google Scholar] [CrossRef]
- Garrido-Lestache, E.; López-Bellido, R.J.; López-Bellido, L. Effect of N Rate, Timing and Splitting and N Type on Bread-Making Quality in Hard Red Spring Wheat under Rainfed Mediterranean Conditions. Field Crops Res. 2004, 85, 213–236. [Google Scholar] [CrossRef]
- Aranguren, M.; Castellón, A.; Aizpurua, A. Wheat Grain Protein Content under Mediterranean Conditions Measured with Chlorophyll Meter. Plants 2021, 10, 374. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal Code for the Growth Stages of Cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- The International Association for Cereal Science and Technology ICC. Standard Methods of the International Association for Cereal Chemistry; The International Association for Cereal Science and Technology ICC: Vienna, Austria, 1992. [Google Scholar]
- Visioli, G.; Comastri, A.; Imperiale, D.; Paredi, G.; Faccini, A.; Marmiroli, N. Gel-Based and Gel-Free Analytical Methods for the Detection of HMW-GS and LMW-GS in Wheat Flour. Food Anal. Methods 2016, 9, 469–476. [Google Scholar] [CrossRef]
- Santiveri, F.; Royo, C.; Romagosa, I. Patterns of grain filling of spring and winter hexaploid triticales. Eur. J. Agron. 2002, 16, 219–230. [Google Scholar] [CrossRef]
- Wheat Protein Market Industry Growth, Trends, and Forecasts|COVID-19 Impact Analysis|Markets and Markets. 2021. Available online: https://www.marketsandmarkets.com/Market-Reports/wheat-protein-market-67845768.html (accessed on 19 April 2021).
- Yu, Z.; Islam, S.; She, M.; Diepeveen, D.; Zhang, Y.; Tang, G.; Zhang, J.; Juhasz, A.; Yang, R.; Ma, W. Wheat Grain Protein Accumulation and Polymerization Mechanisms Driven by Nitrogen Fertilization. Plant J. 2018, 96, 1160–1177. [Google Scholar] [CrossRef]
- Bashir, M.; Ali, S.; Izni, A.; Harun, R. Impact of Excessive Nitrogen Fertilizers on the Environment and Associated Mitigation Strategies. Asian J. Microbiol. Biotechnol. Environ. Sci. 2013, 15, 213–221. [Google Scholar]
- Saint Pierre, C.; Peterson, C.J.; Ross, A.S.; Ohm, J.B.; Verhoeven, M.C.; Larson, M.; Hoefer, B. Winter Wheat Genotypes under Different Levels of Nitrogen and Water Stress: Changes in Grain Protein Composition. J. Cereal Sci. 2008, 47, 407–416. [Google Scholar] [CrossRef]
- Peña, E.; Bernardo, A.; Soler, C.; Jouve, N. Relationship between Common Wheat (Triticum aestivum L.) Gluten Proteins and Dough Rheological Properties. Euphytica 2005, 143, 169–177. [Google Scholar] [CrossRef]
- Zheng, T.; Qi, P.-F.; Cao, Y.-L.; Han, Y.-N.; Ma, H.-L.; Guo, Z.-R.; Wang, Y.; Qiao, Y.-Y.; Hua, S.-Y.; Yu, H.-Y.; et al. Mechanisms of Wheat (Triticum aestivum) Grain Storage Proteins in Response to Nitrogen Application and Its Impacts on Processing Quality. Sci. Rep. 2018, 8, 11928. [Google Scholar] [CrossRef] [PubMed]
- Zhen, S.; Deng, X.; Xu, X.; Liu, N.; Zhu, D.; Wang, Z.; Yan, Y. Effect of High-Nitrogen Fertilizer on Gliadin and Glutenin Subproteomes during Kernel Development in Wheat (Triticum aestivum L.). Crop. J. 2020, 8, 38–52. [Google Scholar] [CrossRef]
- Wang, J.; Pawelzik, E.; Weinert, J.; Zhao, Q.; Wolf, G.A. Effect of Fungicide Treatment on the Quality of Wheat Flour and Breadmaking. J. Agric. Food Chem. 2004, 52, 7593–7600. [Google Scholar] [CrossRef]
- Shewry, P.R.; Underwood, C.; Wan, Y.; Lovegrove, A.; Bhandari, D.; Toole, G.; Mills, E.N.C.; Denyer, K.; Mitchell, R.A.C. Storage Product Synthesis and Accumulation in Developing Grains of Wheat. J. Cereal Sci. 2009, 50, 106–112. [Google Scholar] [CrossRef]
- Blumenthal, C.S.; Barlow, E.W.R.; Wrigley, C.W. Growth Environment and Wheat Quality: The Effect of Heat Stress on Dough Properties and Gluten Proteins. J. Cereal Sci. 1993, 18, 3–21. [Google Scholar] [CrossRef]
- Hernández-Espinosa, N.; Mondal, S.; Autrique, E.; Gonzalez-Santoyo, H.; Crossa, J.; Huerta-Espino, J.; Singh, R.P.; Guzmán, C. Milling, Processing and End-Use Quality Traits of CIMMYT Spring Bread Wheat Germplasm under Drought and Heat Stress. Field Crops Res. 2018, 215, 104–112. [Google Scholar] [CrossRef]
- Malik, A.H.; Kuktaite, R.; Johansson, E. Combined Effect of Genetic and Environmental Factors on the Accumulation of Proteins in the Wheat Grain and Their Relationship to Bread-Making Quality. J. Cereal Sci. 2013, 57, 170–174. [Google Scholar] [CrossRef]
- Souza, E.J.; Martin, J.M.; Guttieri, M.J.; O’Brien, K.M.; Habernicht, D.K.; Lanning, S.P.; McLean, R.; Carlson, G.R.; Talbert, L.E. Influence of Genotype, Environment, and Nitrogen Management on Spring Wheat Quality. Crop Sci. 2004, 44, 425–432. [Google Scholar] [CrossRef]
- Peterson, C.J.; Graybosch, R.A.; Baenziger, P.S.; Grombacher, A.W. Genotype and environment effects on quality characteristics of hard red winter wheat. Crop Sci. 1992, 32, 98–103. [Google Scholar] [CrossRef]
- Panozzo, J.F.; Eagles, H.A. Cultivar and environmental effects on quality characters in wheat. II. Protein. Aust. J. Agric. Res. 2000, 51, 629–636. [Google Scholar] [CrossRef]
- Wang, X.; Liu, F. Effects of Elevated CO2 and Heat on Wheat Grain Quality. Plants 2021, 10, 1027. [Google Scholar] [CrossRef] [PubMed]
- Abonyi, T.; Király, I.; Tömösközi, S.; Baticz, O.; Guóth, A.; Gergely, S.; Scholz, É.; Lásztity, D.; Lásztity, R. Synthesis of Gluten-Forming Polypeptides. 1. Biosynthesis of Gliadins and Glutenin Subunits. J. Agric. Food Chem. 2007, 55, 3655–3660. [Google Scholar] [CrossRef]
- Johansson, E.; Malik, A.H.; Hussain, A.; Rasheed, F.; Newson, W.R.; Plivelic, T.; Hedenqvist, M.S.; Gällstedt, M.; Kuktaite, R. Wheat Gluten Polymer Structures: The Impact of Genotype, Environment, and Processing on Their Functionality in Various Applications. Cereal Chem. 2013, 90, 367–376. [Google Scholar] [CrossRef]
- Lafiandra, D.; Masci, S.; Blumenthal, C.; Wrigley, C.W. The Formation of Glutenin Polymer in Practice. In Cereal Foods World; Symposium American Association of Cereal Chemists: Minneapolis, MI, USA, 1999; pp. 572–578. [Google Scholar]
- Blumenthal, C.S.; Batey, I.L.; Bekes, F.; Wrigley, C.W.; Barlow, E.W.R. Gliadin Genes Contain Heat-Shock Elements: Possible Relevance to Heat-Induced Changes in Grain Quality. J. Cereal Sci. 1990, 11, 185–188. [Google Scholar] [CrossRef]
- Hurkman, W.J.; Tanaka, C.K.; Vensel, W.H.; Thilmony, R.; Altenbach, S.B. Comparative Proteomic Analysis of the Effect of Temperature and Fertilizer on Gliadin and Glutenin Accumulation in the Developing Endosperm and Flour from Triticum aestivum L. Cv. Butte 86. Proteome Sci. 2013, 11, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Year | 0–30 cm | 30–60 cm |
|---|---|---|
| 2014 | 0.109% | 0.086% |
| 2015 | 0.102% | 0.082% |
| 2016 | 0.113% | 0.092% |
| Crop | ||||
|---|---|---|---|---|
| Techniques | Growth Stage a | 2014 | 2015 | 2016 |
| Sowing | 25 October 2013 | 23 October 2014 | 23 October 2015 | |
| N fertilization | tillering (GS23) | 10 March 2014 | 6 March 2015 | 25 February 2016 |
| stem elong. (GS32) | 11 April 2014 | 9 April 2015 | 29 March 2016 | |
| heading (GS55) b | 5 May 2014 | 7 May 2015 | 5 May 2016 | |
| Harvest | 16 July 2014 | 6 July 2015 | 8 July 2016 |
| Milk | Soft Dough | Physiological | Commercial | |
|---|---|---|---|---|
| Years | Stage | Stage | Maturity | Maturity |
| GS75 | GS85 | GS91 | GS99 | |
| 2014 | 3 June 2014 | 13 June 2014 | 23 June 2014 | 16 July 2014 |
| 2015 | 28 May 2015 | 11 June 2015 | 22 June 2015 | 6 July 2015 |
| 2016 | 10 June 2016 | 20 June 2016 | 30 June 2016 | 7 July 2016 |
| Month | 2014 | 2015 | 2016 | |||
|---|---|---|---|---|---|---|
| Rainfall | GDDs a | Rainfall | GDDs | Rainfall | GDDs | |
| (mm) | (°C-Day) | (mm) | (°C-Day) | (mm) | (°C-Day) | |
| November | 81 | 244 | 271 | 289 | 3 | 284 |
| December | 64 | 133 | 92 | 185 | 2 | 182 |
| January | 67 | 158 | 36 | 139 | 4 | 159 |
| February | 87 | 181 | 206 | 143 | 128 | 190 |
| March | 89 | 328 | 188 | 300 | 71 | 286 |
| April | 75 | 442 | 67 | 416 | 80 | 428 |
| May | 49 | 538 | 86 | 581 | 112 | 517 |
| June | 50 | 659 | 55 | 666 | 37 | 642 |
| November–June | 562 | 2683 | 1000 | 2720 | 436 | 2689 |
| November–April | 463 | 1485 | 859 | 1473 | 288 | 1530 |
| May–June | 99 | 1198 | 141 | 1247 | 149 | 1159 |
| Factor | Source of Variation | TKW | GPC | Gluten Extractable Fractions (mg g−1 Flour) | ||
|---|---|---|---|---|---|---|
| g | % | HMW-gs | LMW-gs | Gliadins | ||
| Late N | unfertilized control | 34.60 a | 14.41 b | 5.27 b | 6.26 b | 14.23 b |
| fertilization (N) | heading application | 34.85 a | 15.64 a | 5.83 a | 7.21 a | 15.79 a |
| p-value | 0.682 | <0.001 | <0.001 | <0.001 | <0.001 | |
| Fungicide | untreated control | 33.88 b | 15.19 a | 5.60 a | 6.67 a | 15.32 a |
| application (F) | heading application | 35.61 a | 14.86 b | 5.50 a | 6.80 a | 14.66 b |
| p-value | <0.001 | 0.034 | 0.281 | 0.205 | 0.014 | |
| Year (Y) | 2014 | 34.55 b | 14.96 b | 4.86 b | 6.50 b | 13.60 b |
| 2015 | 33.87 c | 15.43 a | 4.50 c | 5.42 c | 18.69 a | |
| 2016 | 36.01 a | 14.69 b | 7.29 a | 8.34 a | 12.43 c | |
| p-value | <0.001 | 0.003 | <0.001 | <0.001 | <0.001 | |
| N * F | p-value | 0.953 | 0.096 | 0.006 | 0.087 | 0.034 |
| N * Y | p-value | 0.925 | 0.204 | 0.181 | 0.019 | 0.124 |
| F * Y | p-value | 0.907 | 0.235 | 0.277 | 0.258 | 0.170 |
| N * F * Y | p-value | 0.996 | 0.136 | 0.047 | 0.443 | 0.797 |
| Factor | Source of Variation | Gluten Protein Fraction Ratio | Alveographic Parameters | |||
|---|---|---|---|---|---|---|
| gs/glia | H/L | Y/X (H) | W | P/L | ||
| J * 10−4 | ||||||
| Late N | unfertilized control | 0.85 a | 0.84 a | 0.58 a | 337 b | 1.99 a |
| fertilization (N) | heading application | 0.88 a | 0.80 a | 0.58 a | 414 a | 1.74 b |
| p-value | 0.210 | 0.103 | 0.956 | <0.001 | 0.004 | |
| Fungicide | untreated control | 0.86 a | 0.84 a | 0.59 a | 383 a | 1.89 a |
| application (F) | heading application | 0.87 a | 0.80 a | 0.57 a | 365 b | 1.85 a |
| p-value | 0.071 | 0.046 | 0.576 | 0.045 | 0.574 | |
| Year (Y) | 2014 | 0.83 b | 0.75 b | 0.41 c | 390 a | 1.75 b |
| 2015 | 0.53 c | 0.84 a | 0.61 b | 322 b | 1.64 b | |
| 2016 | 1.26 a | 0.88 a | 0.72 a | 409 a | 2.24 a | |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | |
| N * F | p-value | 0.006 | <0.001 | 0.027 | 0.099 | 0.338 |
| N * Y | p-value | 0.084 | 0.356 | 0.823 | 0.156 | 0.039 |
| F * Y | p-value | 0.404 | 0.214 | 0.571 | 0.640 | 0.754 |
| N * F * Y | p-value | 0.373 | 0.163 | 0.580 | 0.754 | 0.742 |
| Fungicide | Late N | Gluten Fractions | Gluten Fraction Ratio | ||||
|---|---|---|---|---|---|---|---|
| Application | Fertilization | HMW-gs | LMW-gs | Gliadins | gs/glia | H/L | Y/X (H) |
| untreated control | unfertilized control | 5.46 b | 6.12 b | 14.28 bc | 0.87 ab | 0.91 a | 0.62 a |
| heading application | 5.74 ab | 7.23 a | 16.48 a | 0.85 ab | 0.78 b | 0.55 a | |
| heading application | unfertilized control | 5.08 c | 6.41 b | 14.19 c | 0.84 b | 0.76 b | 0.53 a |
| heading application | 5.91 a | 7.19 a | 15.15 b | 0.90 a | 0.83 b | 0.61 a | |
| p-value | <0.001 | <0.001 | <0.001 | 0.009 | <0.001 | 0.137 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Landolfi, V.; Visioli, G.; Blandino, M. Effect of Nitrogen Fertilization and Fungicide Application at Heading on the Gluten Protein Composition and Rheological Quality of Wheat. Agronomy 2021, 11, 1687. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091687
Landolfi V, Visioli G, Blandino M. Effect of Nitrogen Fertilization and Fungicide Application at Heading on the Gluten Protein Composition and Rheological Quality of Wheat. Agronomy. 2021; 11(9):1687. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091687
Chicago/Turabian StyleLandolfi, Viola, Giovanna Visioli, and Massimo Blandino. 2021. "Effect of Nitrogen Fertilization and Fungicide Application at Heading on the Gluten Protein Composition and Rheological Quality of Wheat" Agronomy 11, no. 9: 1687. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091687