
Structure of Anther Epidermis and Endothecium, Production of Pollen, and Content of Selected Nutrients in Pollen Grains from Six Rubus idaeus L. Cultivars

Abstract
:1. Introduction
1.1. Anther Epidermis and Andothecium
1.2. Pollen as a Source of Nutrients
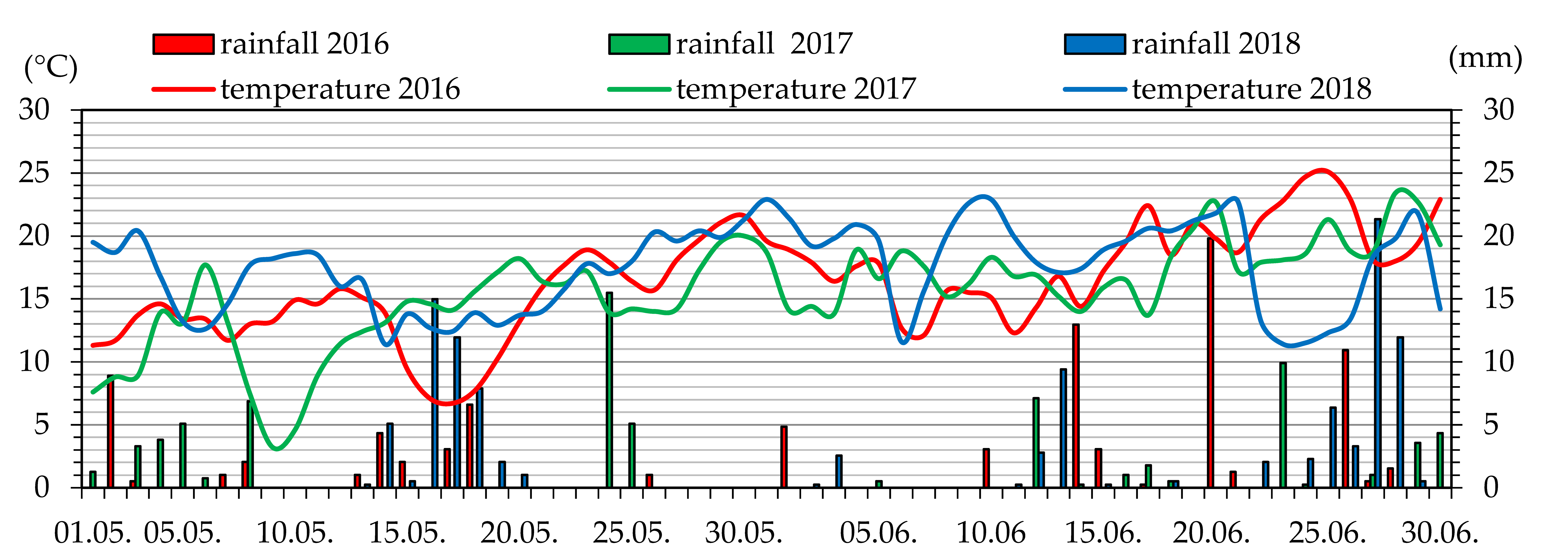
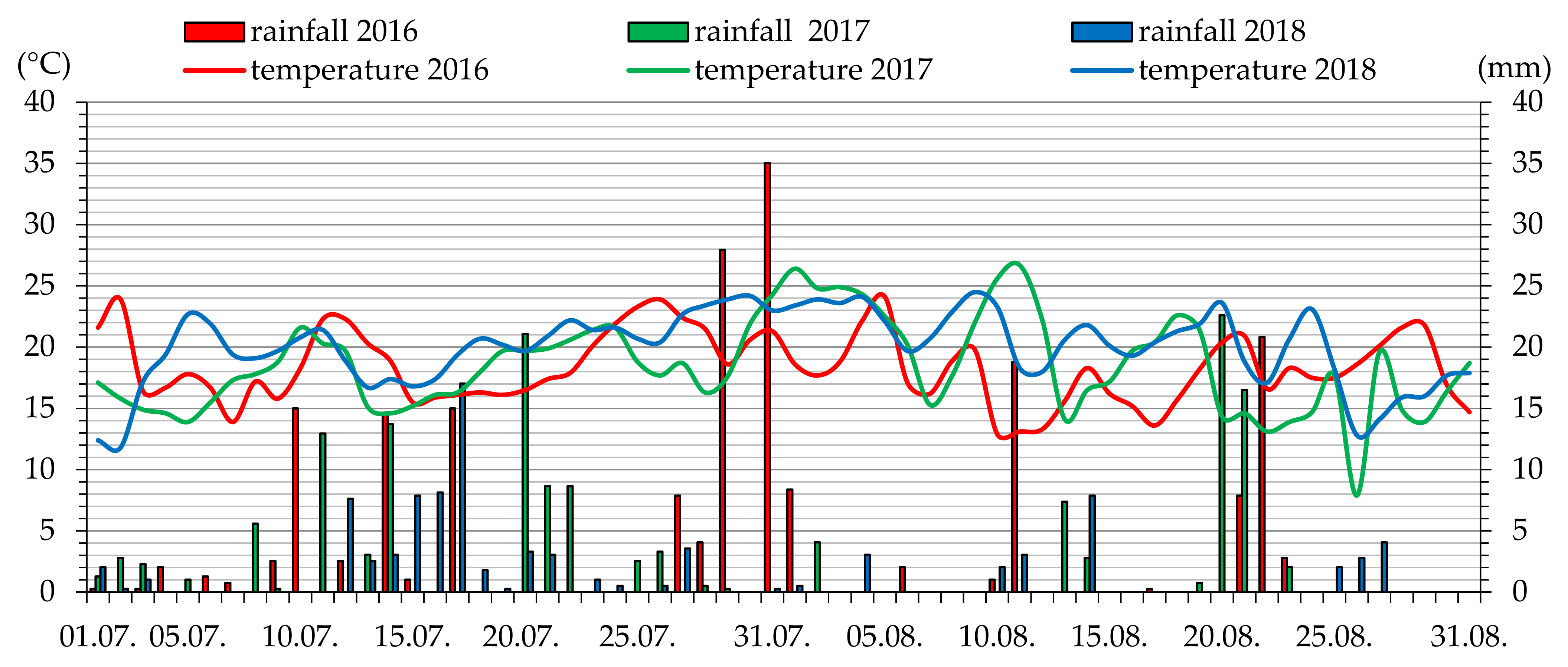
2. Materials and Methods
2.1. Plant Material
2.2. Fixation of Material for Microscopy
2.3. Light Microscopy
2.4. Scanning Electron Microscopy (SEM)
2.5. Transmission Electron Microscopy (TEM)
2.6. Number and Length of Stamens
2.7. Pollen Mass
2.8. Energy Dispersive Spectroscopy (EDS) Microanalysis of Elements
2.9. Total Fat Content and Qualitative and Quantitative Fatty Acid Composition
2.10. Total Protein Content
2.11. Qualitative and Quantitative Amino Acid Composition
2.12. Statistical Analysis of Results
3. Results
3.1. Number and Length of Stamens
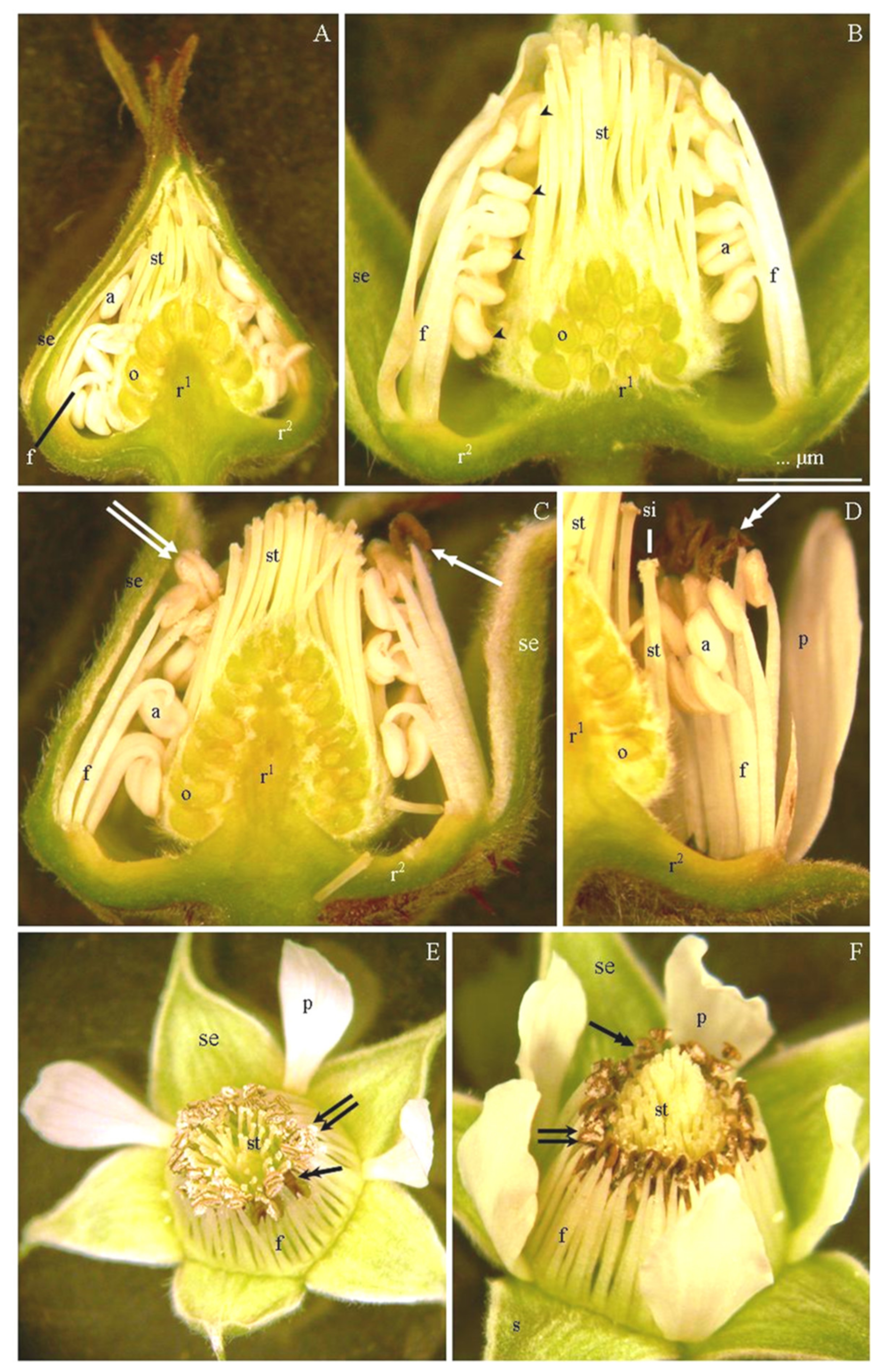
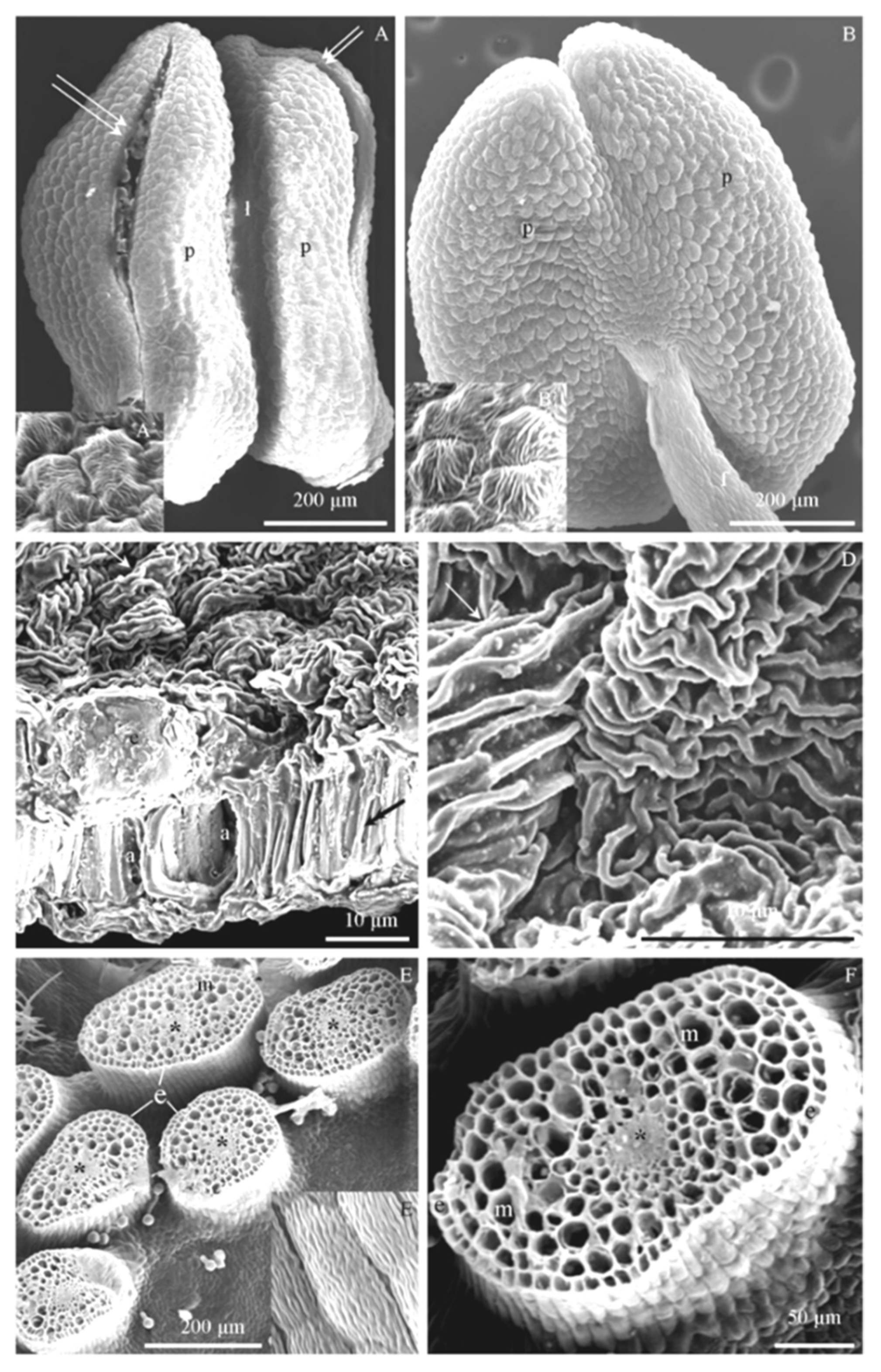
3.2. Micromorphology of Stamen Epidermis
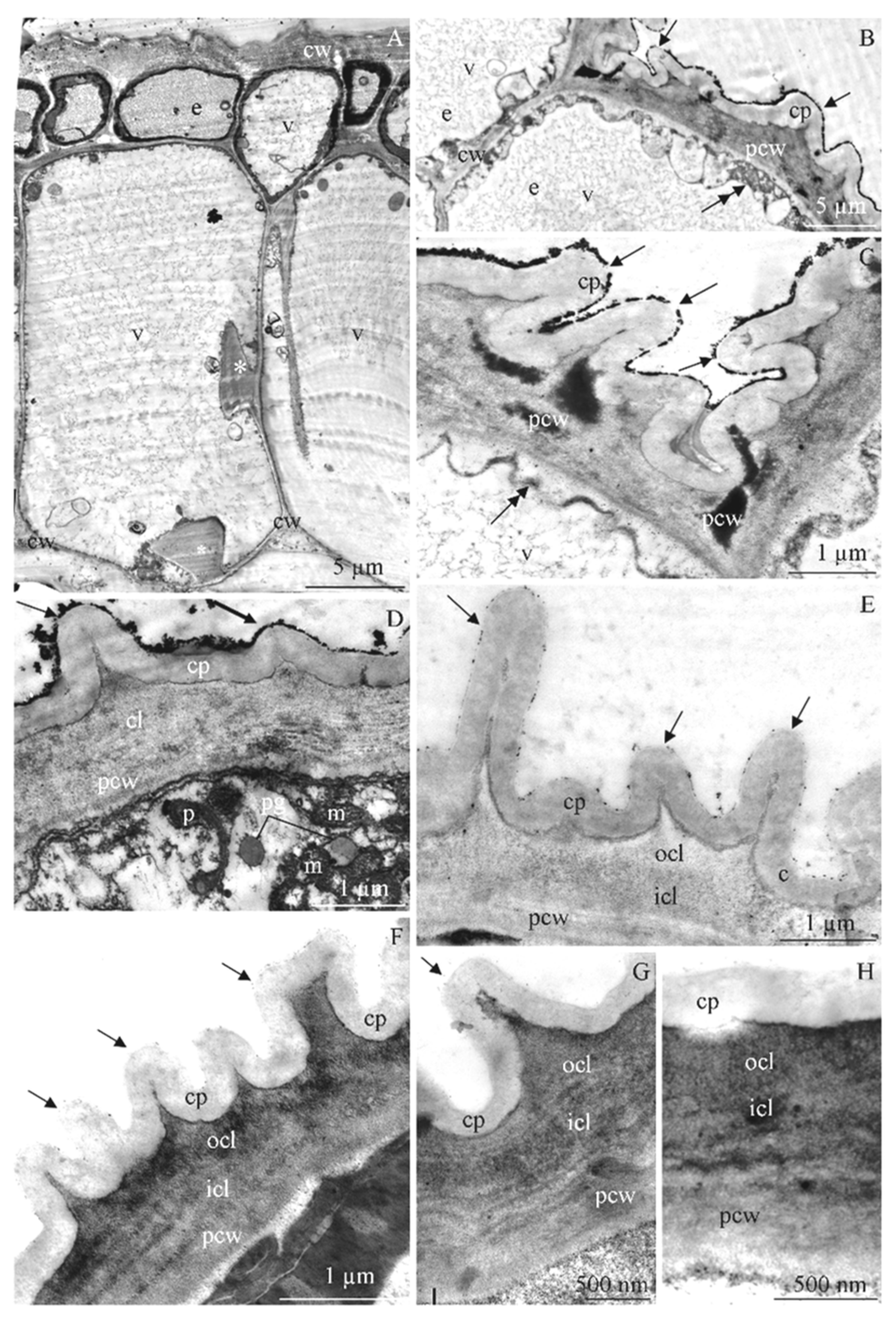
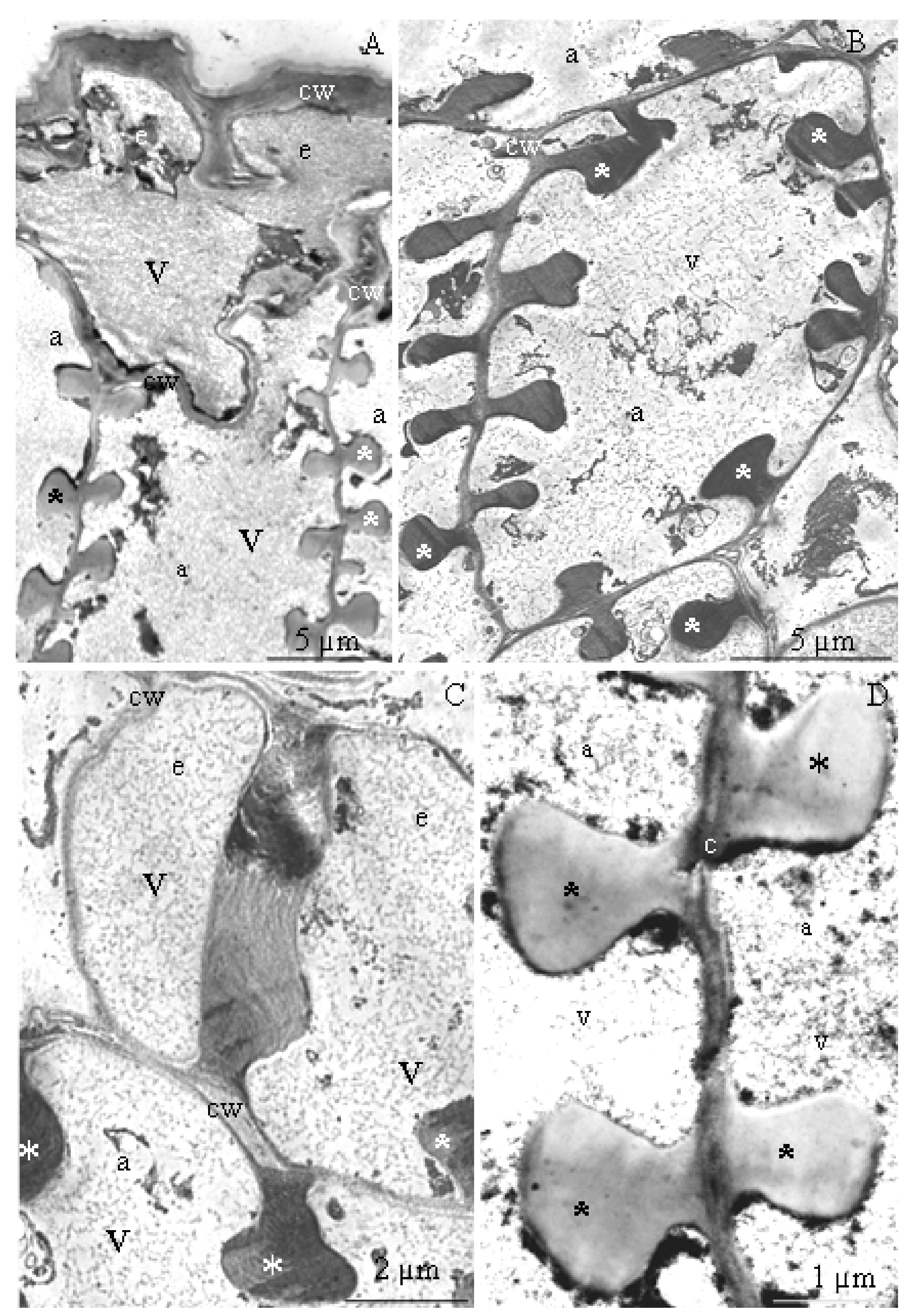
3.3. Anatomy of Stamens
3.4. Ultrastructure of Anther Cells
3.5. The Mass of Pollen Produced
3.6. Total Protein Content
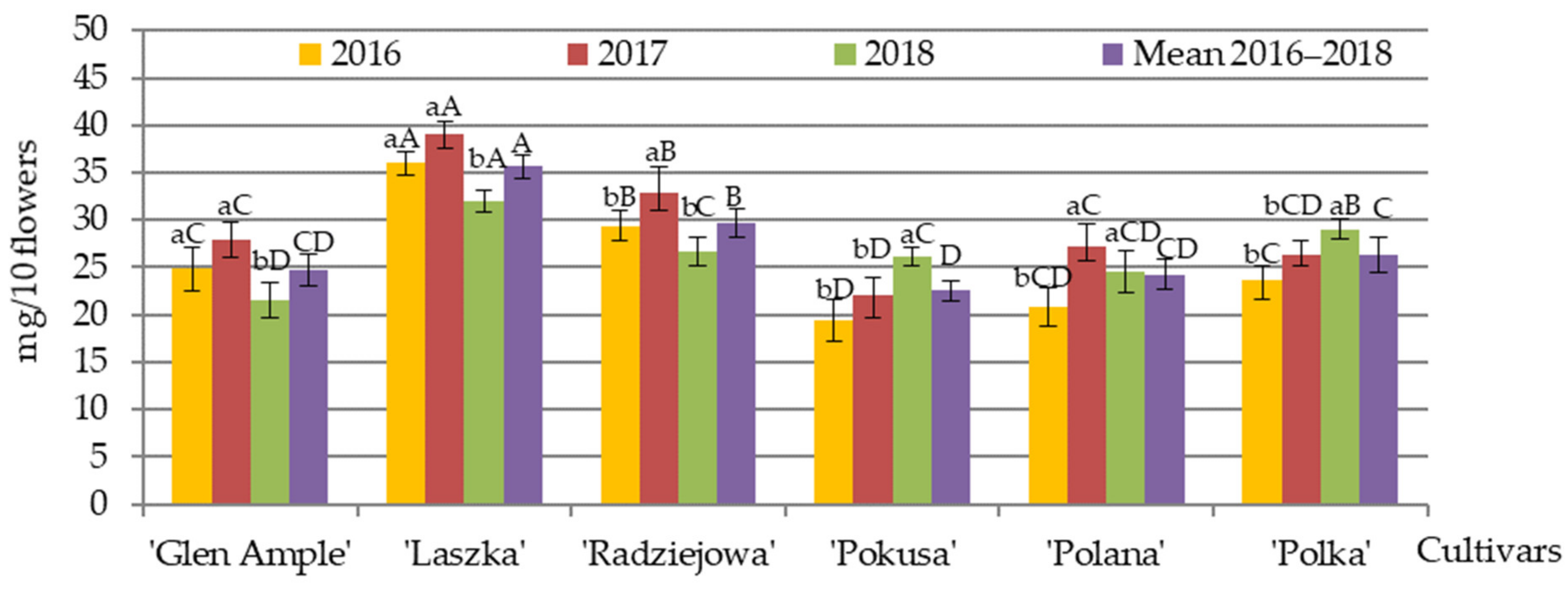
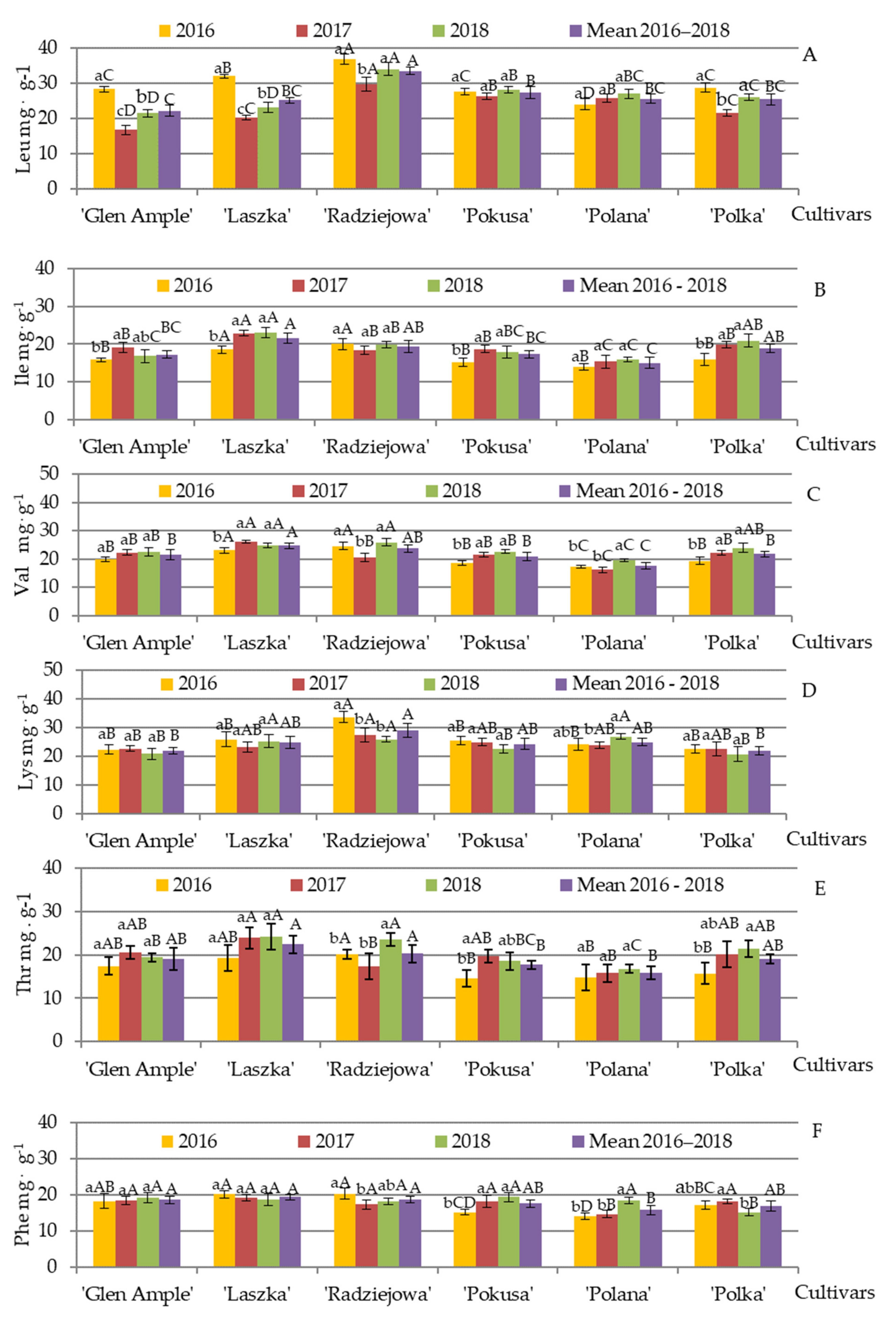
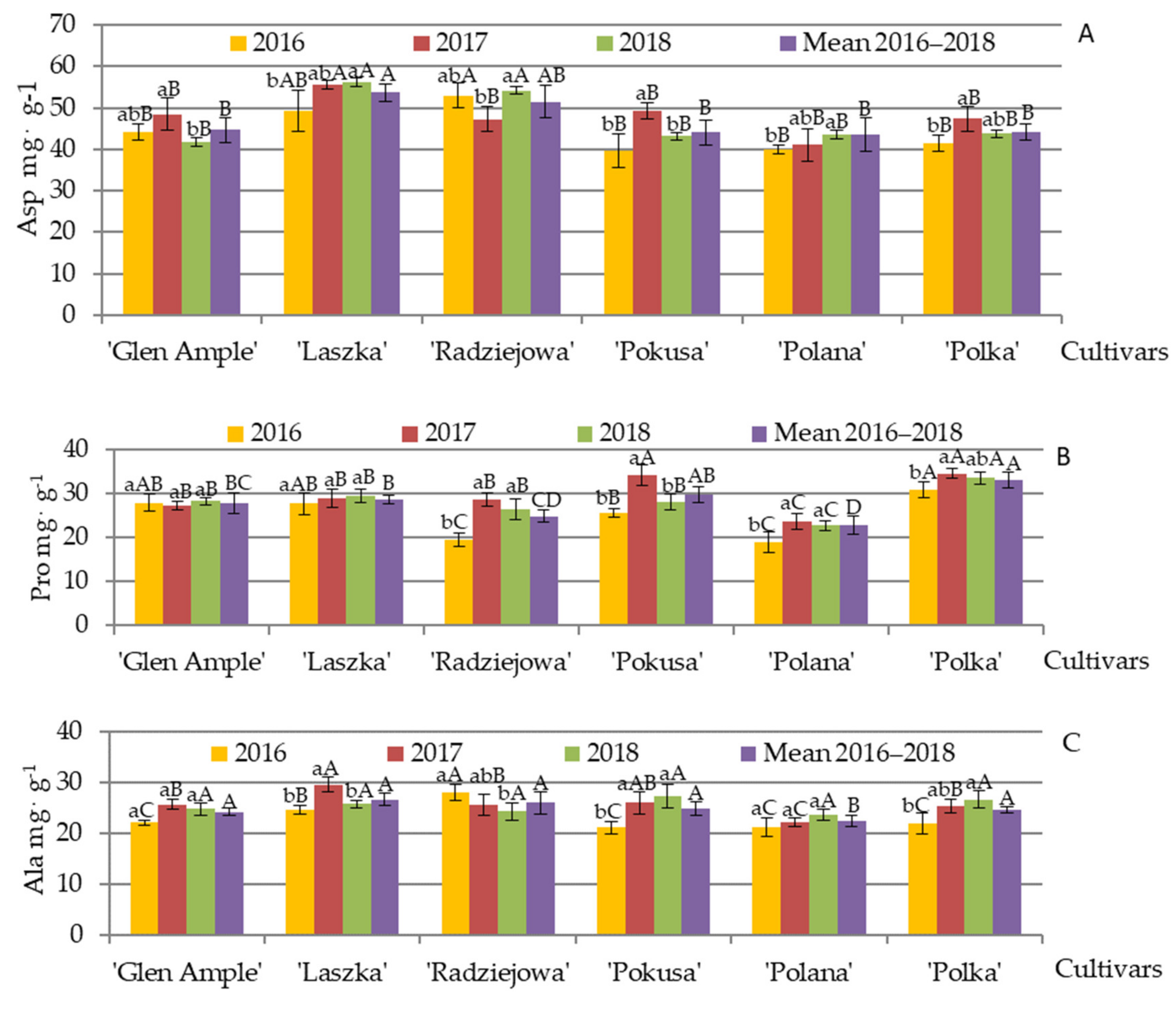
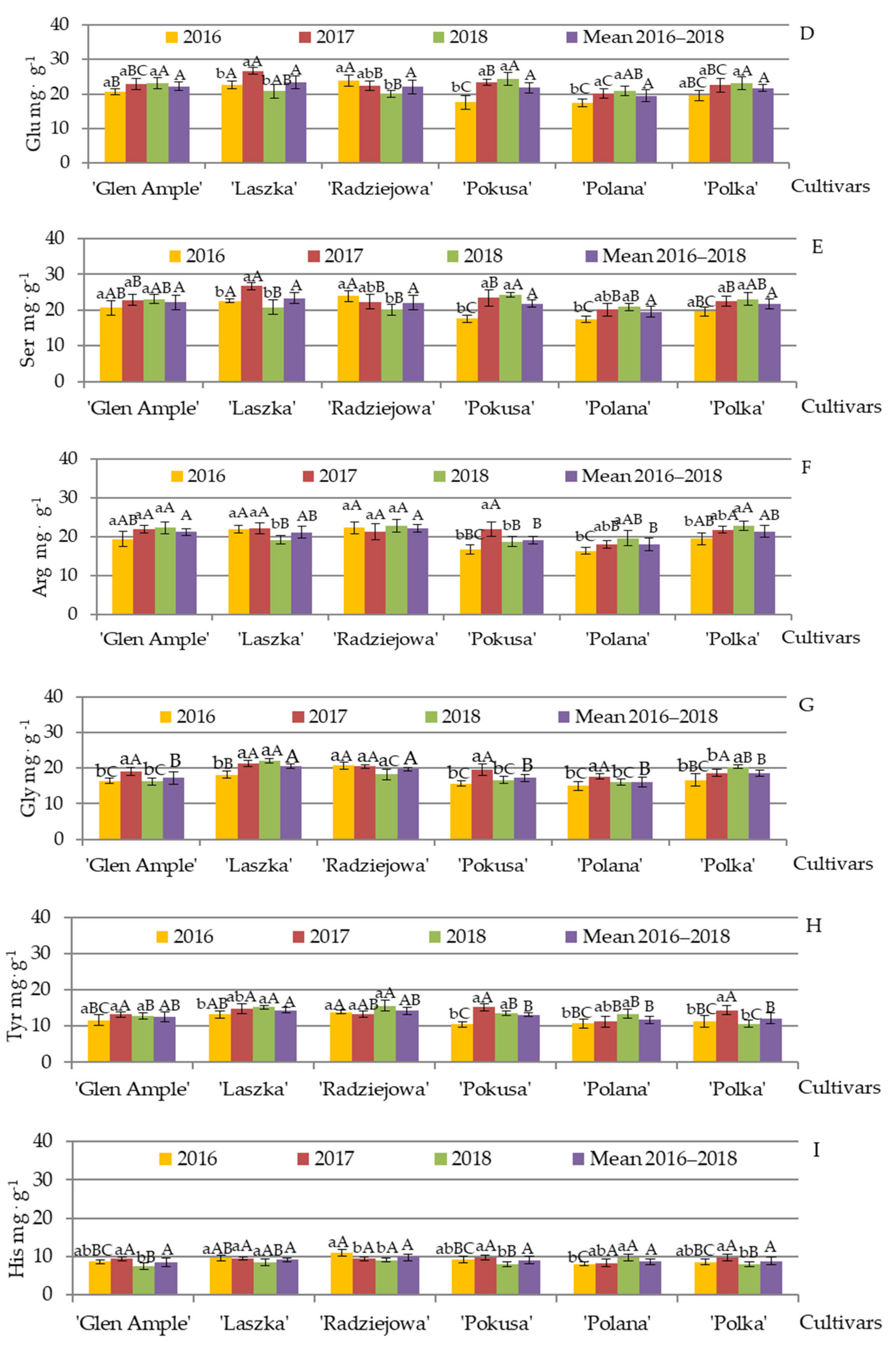
3.7. Qualitative and Quantitative Amino Acid Composition
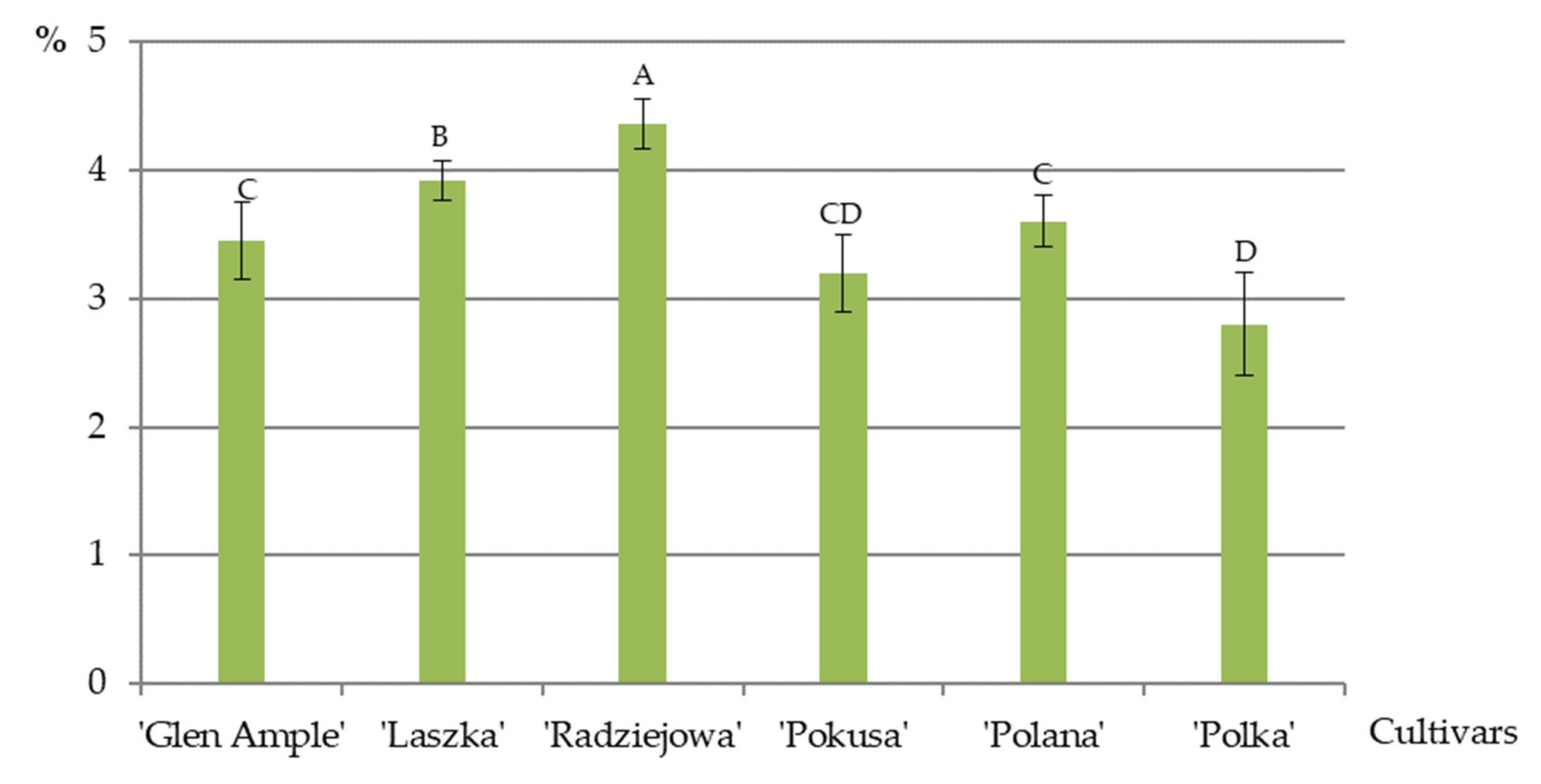
3.8. Total Fat Content
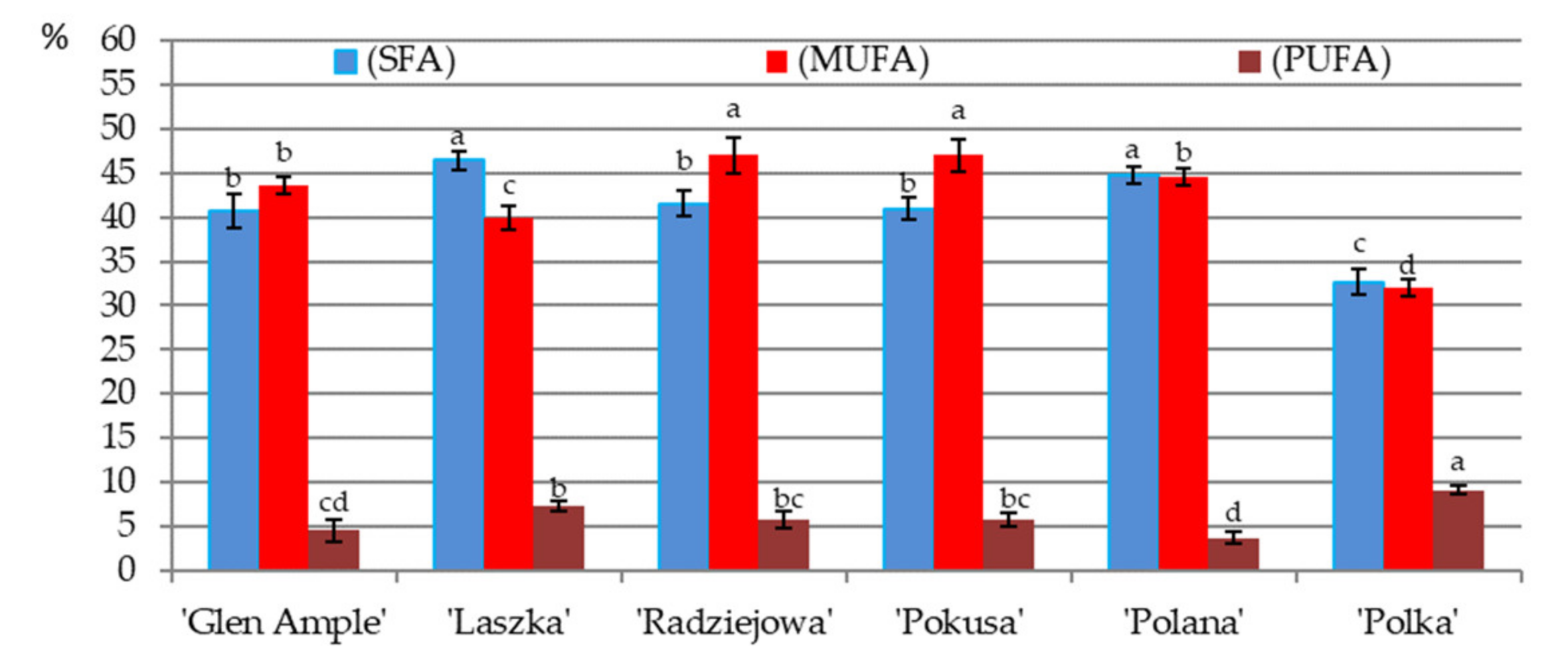
3.9. Fatty Acid Content in Pollen
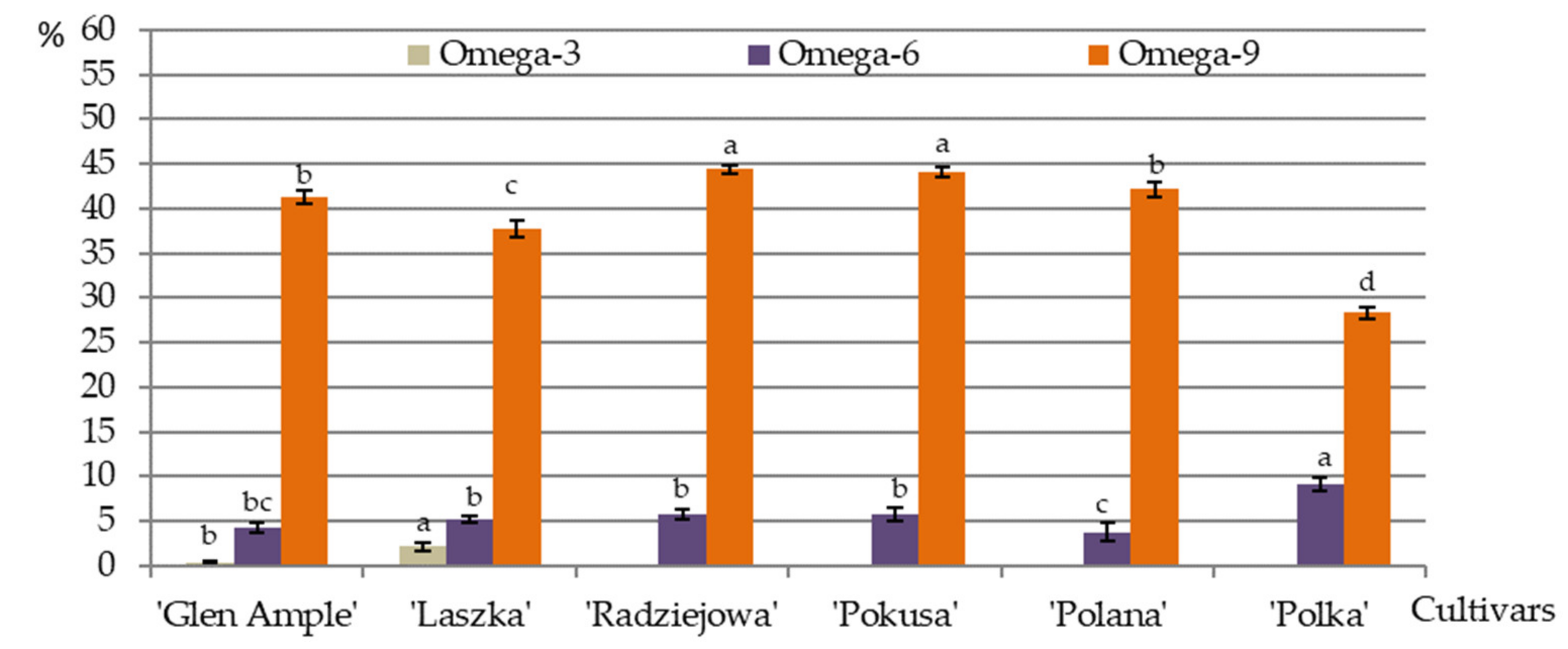
3.10. Omega-3, Omega-6, and Omega-9 Acids
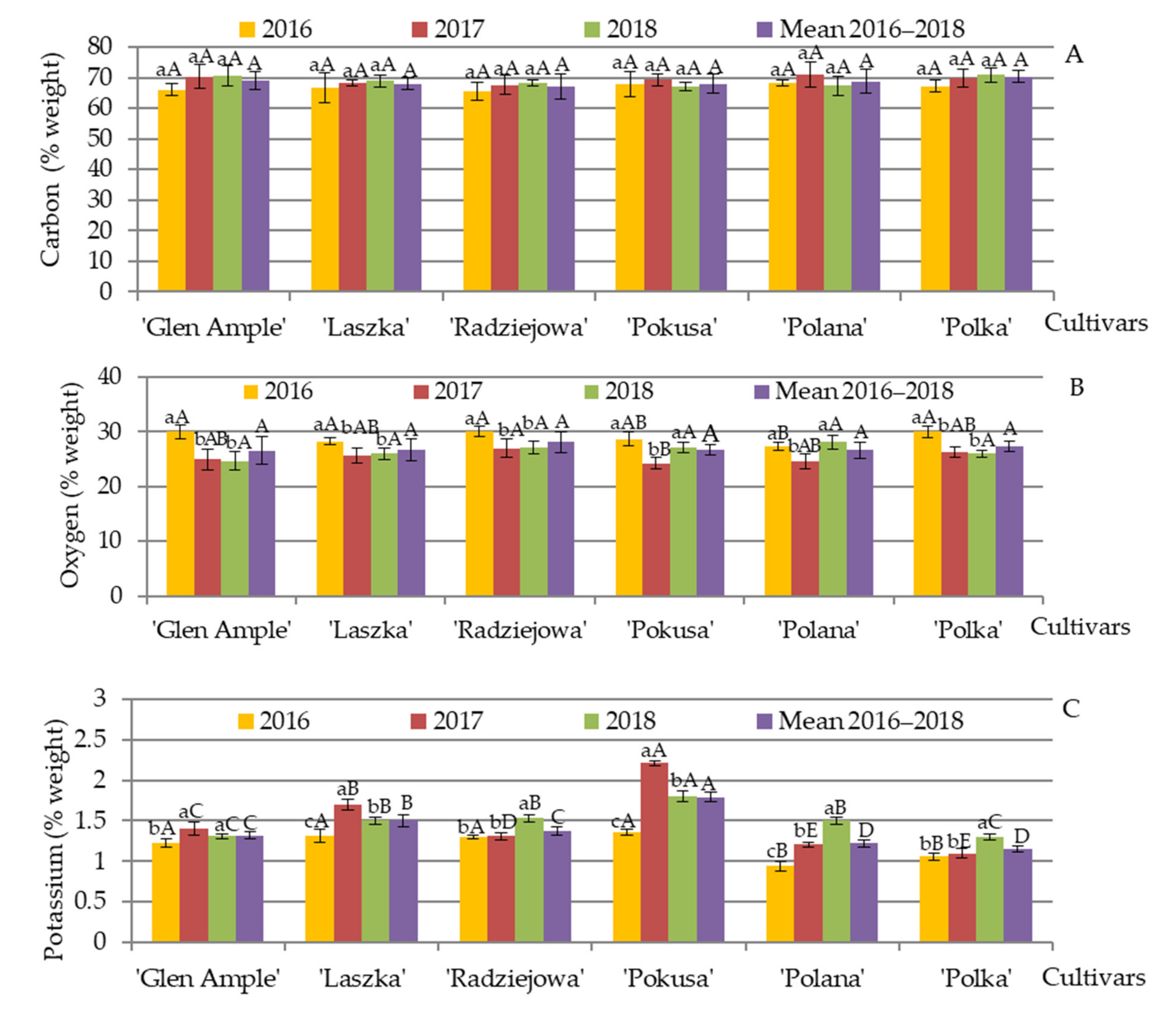
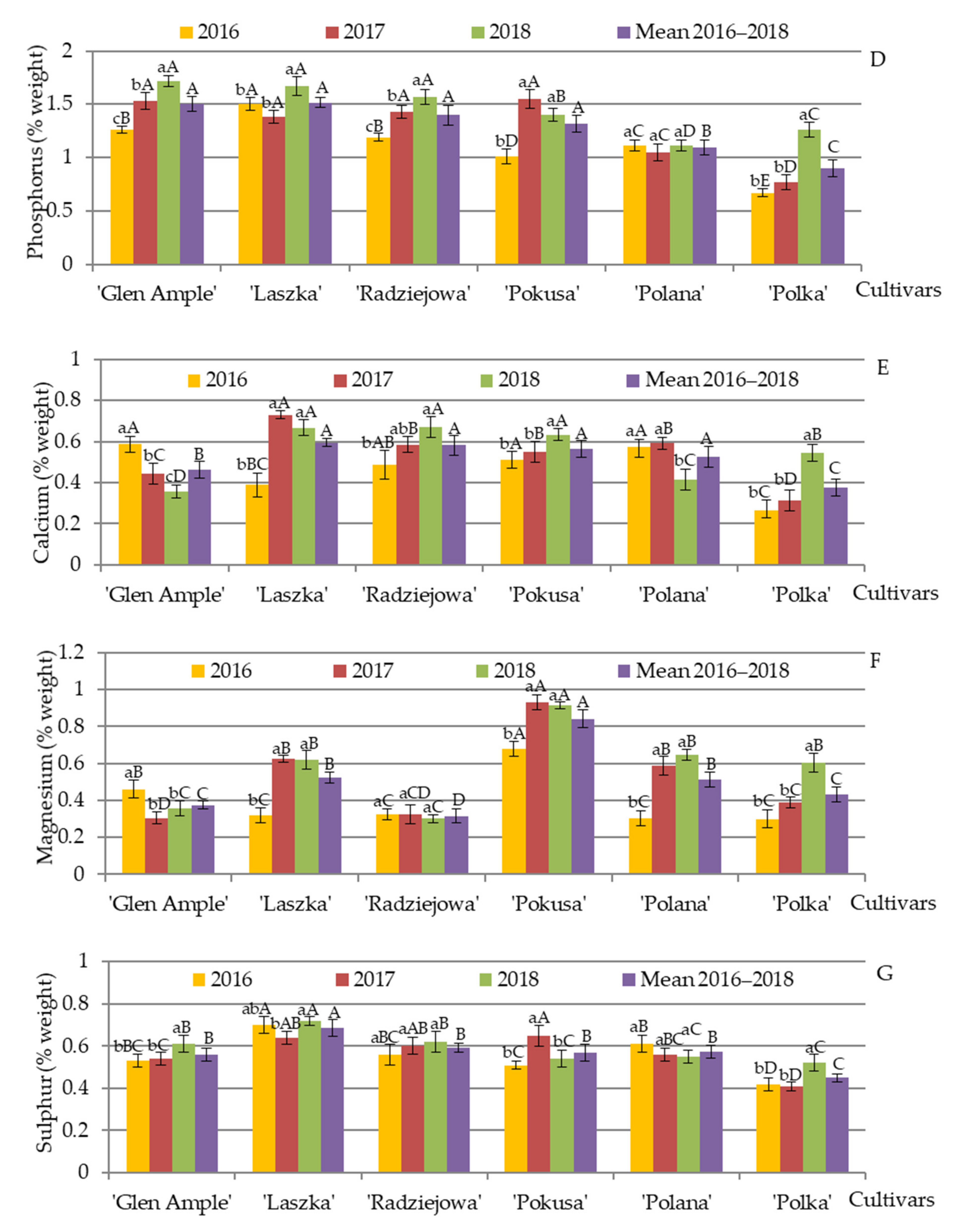
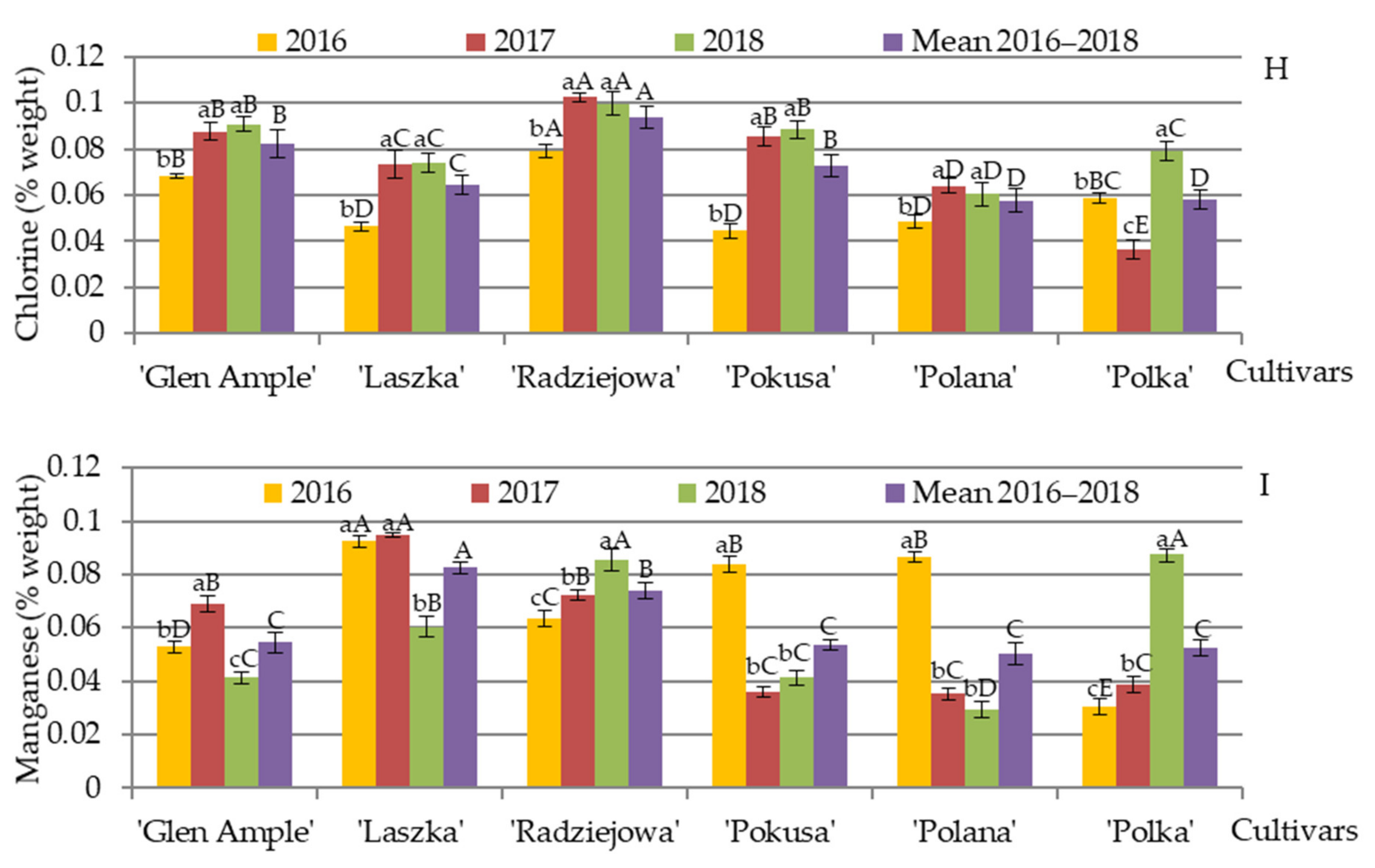
3.11. Content of Selected Minerals in Pollen Exine
4. Discussion
4.1. Micromorphology of Stamen Epidermis
4.2. Number and Size of Stamens
4.3. Structure of Epidermis and Endothecium Cells
4.4. Pollen Mass
4.5. Biologically Active Chemical Compounds
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lebedev, V.G.; Subbotina, N.M.; Maluchenko, O.P.; Krutovsky, K.V.; Shestibratov, K.A. Assessment of genetic diversity in differently colored raspberry cultivars using SSR markers located in flavonoid biosynthesis genes. Agronomy 2019, 9, 518. [Google Scholar] [CrossRef] [Green Version]
- Pinczinger, D.; Reth, M.V.; Keilwagen, J.; Berner, T.; Peil, A.; Flachowsky, H.; Emeriewen, O.F. Mapping of the waxy bloom gene in ‘Black Jewel’ in a parental linkage map of ‘Black Jewel’בGlen Ample’ (Rubus) interspecific population. Agronomy 2020, 10, 1579. [Google Scholar] [CrossRef]
- Khadgi, A.; Weber, C.A. RNA-Seq analysis of prickled and prickle-free epidermis provides insight into the genetics of prickle development in red raspberry (Rubus ideaus L.). Agronomy 2020, 10, 1904. [Google Scholar] [CrossRef]
- Khadgi, A.; Weber, C.A. Genome-wide association study (GWAS) for examining the genomics controlling prickle production in red raspberry (Rubus idaeus L.). Agronomy 2021, 11, 27. [Google Scholar] [CrossRef]
- FAO. Food and Agriculture Organization of the United Nations. 2019. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 24 April 2021).
- Heide, O.M.; Sønsteby, A. Physiology of flowering and dormancy regulation in annual-and biennial-fruiting red raspberry (Rubus idaeus L.)—A review. J. Hortic. Sci. Biotechnol. 2011, 86, 433–442. [Google Scholar] [CrossRef]
- Sønsteby, A.; Stavang, J.A.; Heide, O.M. Production of high-yielding raspberry long canes: The way to 3 kg of fruit per cane. J. Hortic. Sci. Biotechnol. 2013, 88, 591–599. [Google Scholar] [CrossRef]
- Schmidt, K.; Filep, R.; Orosz-Kovács, Z.; Farkas, Á. Patterns of nectar and pollen presentation influence the attractiveness of four raspberry and blackberry cultivars to pollinators. J. Hortic. Sci. Biotech. 2015, 90, 47–56. [Google Scholar] [CrossRef]
- Campos, M.G.R.; Frigerio, C.; Lopes, J.; Bogdanov, S. What is the future of bee-pollen. J. ApiProd. ApiMed. Sci. 2010, 2, 131–144. [Google Scholar] [CrossRef]
- Abouda, Z.; Zerdani, I.; Kalalou, I.; Faid, M.; Ahami, M.T. The antibacterial activity of moroccan bee bread and bee-pollen (fresh and dried) against pathogenic bacteria. Res. J. Microbiol. 2011, 6, 376–384. [Google Scholar]
- Conte, P.; Del Caro, A.; Balestra, F.; Piga, A.; Fadda, C. Bee pollen as a functional ingredient in gluten-free bread: A physical-chemical, technological and sensory approach. LWT Food Sci. Technol. 2018, 90, 1–7. [Google Scholar] [CrossRef]
- Kudrik, B.T.; Tykhonov, O.I.; Koval, V.M.; Shpychak, O.S.; Rudenko, V.V. Development of the composition of foamy washing liquid with an extract of pollen for use in pediatrics. J. Pharm. Sci. Res. 2018, 10, 1017–1021. [Google Scholar]
- Lale, S.V.; Gill, H.S. Pollen grains as a novel microcarrier for oral delivery of proteins. Int. J. Pharm. 2018, 552, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Scott, R.J.; Spielman, M.; Dickinson, H.G. Stamen structure and function. Plant Cell. 2004, 16, S46–S60. [Google Scholar] [CrossRef]
- García, C.C.; Nepi, M.; Pacini, E. It is a matter of timing: Asynchrony during pollen development and its consequences on pollen performance in angiosperms—A review. Protoplasma 2017, 254, 57–73. [Google Scholar] [CrossRef]
- Kostić, A.Ž.; Milinčić, D.D.; Barać, M.B.; Ali Shariati, M.; Tešić, Ž.L.; Pešić, M.B. The application of pollen as a functional food and feed ingredient—The present and perspectives. Biomolecules 2020, 10, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halbritter, H.; Ulrich, S.; Grímsson, F.; Weber, M.; Zetter, R.; Hesse, M.; Buchner, R.; Svojtka, M.; Frosch-Radivo, A. Pollen morphology and ultrastructure. In Illustrated Pollen Terminology, 2nd ed.; Springer: Cham, Switzerland, 2018; pp. 37–65. [Google Scholar] [CrossRef] [Green Version]
- Kostryco, M.; Chwil, M.; Matraszek-Gawron, R. Comparison of the micromorphology and ultrastructure of pollen grains of selected Rubus idaeus L. cultivars grown in commercial plantation. Plants 2020, 9, 1194. [Google Scholar] [CrossRef] [PubMed]
- Rezanejad, F. The structure and ultrastructure of anther epidermis and pollen in Lagerstroemia indica L. (Lythraceae) in response to air pollution. Turk. J. Botany 2008, 32, 35–42. [Google Scholar]
- Pollard, M.; Beisson, F.; Li, Y.; Ohlrogge, J.B. Building lipid barriers: Biosynthesis of cutin and suberin. Trends Plant Sci. 2008, 13, 236–246. [Google Scholar] [CrossRef]
- Samuels, L.; Kunst, L.; Jetter, R. Sealing plant surfaces: Cuticular wax formation by epidermal cells. Annu. Rev. Plant Biol. 2008, 59, 683–707. [Google Scholar] [CrossRef] [Green Version]
- Beisson, F.; Li-Beisson, Y.; Pollard, M. Solving the puzzles of cutin and suberin polymer biosynthesis. Curr. Opin. Plant Biol. 2012, 15, 329–337. [Google Scholar] [CrossRef]
- Tian, Y.; Xiao, S.; Liu, J.; Somaratne, Y.; Zhang, H.; Wang, M.; Zhang, H.; Zhao, L.; Chen, H. Male STERILE6021 (MS6021) is required for the development of anther cuticle and pollen exine in maize. Sci. Rep. 2017, 7, 16736. [Google Scholar] [CrossRef]
- Somaratne, Y.; Tian, Y.; Zhang, H.; Wang, M.; Huo, Y.; Cao, F.; Zhao, L.; Chen, H. Abnormal pollen vacuolation1 (APV1) is required for male fertility by contributing to anther cuticle and pollen exine formation in maize. Plant J. 2017, 90, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Qu, S.; Tucker, M.R.; Zhang, D.; Liang, W.; Shi, J. Ostkpr1 functions in anther cuticle development and pollen wall formation in rice. BMC Plant Biol. 2019, 19, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Z.; Chen, Z.; Yan, W.; Xie, G.; Lu, J.; Wang, N.; Lu, Q.; Yao, N.; Yang, G.; Xia, J.; et al. An ABC transporter, OsABCG26, is required for anther cuticle and pollen exine formation and pollen-pistil interactions in rice. Plant Sci. 2016, 253, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Z.; Jin, M.; Yan, W.; Chen, H.; Qiu, S.; Fu, S.; Xia, J.; Liu, Y.; Chen, Z.; Wu, J.; et al. The ATP-binding cassette (ABC) transporter OsABCG3 is essential for pollen development in rice. Rice 2018, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, H.; Sun, H.; Luo, H.; Zhao, L.; Dong, Z.; Yan, C.Z.; Liu, R.; Chunyan, X.; Li, S.; et al. Irregular pollen exine is a novel factor in anther cuticle and pollen exine formation. Plant Physiol. 2017, 173, 307–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecchetti, V.; Altamura, M.M.; Brunetti, P.; Petrocelli, V.; Falasca, G.; Ljung, K.; Costantino, P.; Cardarelli, M. Auxin controls Arabidopsis anther dehiscence by regulating endothecium lignification and jasmonic acid biosynthesis. Plant J. 2013, 74, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.W.; Kim, Y.Y.; Yoo, K.S.; Ok, S.H.; Cui, M.H.; Jeong, B.C.; Yoo, S.D.; Jeung, J.U.; Shin, J.S. A cystathionine-β-synthase domain-containing protein, CBSX2, regulates endothecial secondary cell wall thickening in anther development. Plant Cell Physiol. 2013, 54, 195–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Xu, Z.; Song, J.; Conner, K.; Vizcay Barrena, G.; Wilson, Z.A. Arabidopsis MYB26/MALE STERILE35 regulates secondary thickening in the endothecium and is essential for anther dehiscence. Plant Cell 2007, 19, 534–548. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Song, J.; Ferguson, A.C.; Klisch, D.; Simpson, K.; Mo, R.; Taylor, B.; Mitsuda, N.; Wilson, Z.A. Transcription factor MYB26 is key to spatial specificity in anther secondary thickening formation. Plant Physiol. 2017, 175, 333–350. [Google Scholar] [CrossRef]
- Jung, K.W.; Oh, S.I.; Kim, Y.Y.; Yoo, K.S.; Cui, M.H.; Shin, J.S. Arabidopsis histidine-containing phosphotransfer factor 4 (AHP4) negatively regulates secondary wall thickening of the anther endothecium during flowering. Mol. Cells 2008, 25, 294–300. [Google Scholar] [PubMed]
- Zhao, S.Q.; Li, W.C.; Zhang, Y.; Tidy, A.C.; Wilson, Z.A. Knockdown of Arabidopsis root UVB SENSITIVE4 disrupts anther dehiscence by suppressing secondary thickening in the endothecium. Plant Cell Physiol. 2019, 60, 2293–2306. [Google Scholar] [CrossRef] [PubMed]
- Pernal, S.F.; Currie, R.W. The influence of pollen quality on foraging behavior in honeybees (Apis mellifera L.). Behav. Ecol. Sociobiol. 2001, 51, 53–68. [Google Scholar] [CrossRef]
- Hanley, M.E.; Franco, M.; Pichon, S.; Darvill, B.; Goulson, D. Breeding system, pollinator choice and variation in pollen quality in British herbaceous plants. Funct. Ecol. 2008, 22, 592–598. [Google Scholar] [CrossRef]
- Weiner, C.N.; Hilpert, A.; Werner, M.; Linsenmair, K.E.; Blüthgen, N. Pollen amino acids and flower specialisation in solitary bees. Apidologie 2010, 41, 476–487. [Google Scholar] [CrossRef] [Green Version]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.L.; Alaux, C. Influence of pollen nutrition on honey bee health: Do pollen quality and diversity matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanderplanck, M.; Leroy, B.; Wathelet, B.; Wattiez, R.; Michez, D. Standardized protocol to evaluate pollen polypeptides as bee food source. Apidologie 2014, 45, 192–204. [Google Scholar] [CrossRef] [Green Version]
- Conti, I.; Medrzycki, P.; Argenti, C.; Meloni, M.; Vecchione, V.; Boi, M.; Mariotti, M.G. Sugar and protein content in different monofloral pollens-building a database. Bull. Insectol. 2016, 69, 318–320. [Google Scholar]
- Liolios, V.; Tananaki, C.; Dimou, M.; Kanelis, D.; Goras, G.; Kazrazafiris, E.; Thrasuvoulou, A. Ranking pollen from bee plants according to their protein contribution to honey bees. J. Apic. Res. 2016, 54, 582–592. [Google Scholar] [CrossRef]
- Roulston, T.A.H.; Cane, J.H.; Buchmann, S.L. What governs protein content of pollen: Pollinator preferences, pollen–pistil interactions, or phylogeny? Ecol. Monogr. 2000, 70, 617–643. [Google Scholar] [CrossRef]
- Heslop-Harrison, J.; Haslop-Harrison, Y.; Knox, R.B.; Howlett, B. Pollen-wall proteins: ‘gametophytic’ and ‘sporophytic’ fractions in the pollen walls of the Malvaceae. Ann. Bot. 1973, 37, 403–412. [Google Scholar] [CrossRef]
- Heslop-Harrison, Y. Pollen germination and pollen tube growth. Int. Rev. Cytol. 1987, 107, 1–78. [Google Scholar] [CrossRef]
- Dickinson, H.G.; Elleman, C.J.; Doughty, J. Pollen coatings—Chimaeric genetics and new functions. Sex. Plant Reprod. 2000, 12, 302–309. [Google Scholar] [CrossRef]
- Obermeyer, G.; Kriechbaumer, R.; Strasser, D.; Maschessing, A.; Bentrup, F.W. Boric acid stymulates the plasma membrane H+ ATPase of ungerminated lily pollen grains. Physiol. Plant. 1996, 98, 281–290. [Google Scholar] [CrossRef]
- Pertl, H.; Himly, M.; Gehwolf, R.; Kriechbaumer, R.; Strasser, D.; Michalke, W.; Obermeyer, G. Molecular and physiological characterisation of a 14-3-3 protein from lily pollen grains regulating the activity of the plasma membrane H+ ATPase during pollen grain germination and tube growth. Planta 2001, 213, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Scolari, L.M.; Hancock, R.D.; Hedley, P.E.; Morris, J.; Smith, K.; Graham, J. Combining QTL mapping and gene expression analysis to elucidate the genetic control of ‘crumbly’ fruit in red raspberry (Rubus idaeus L.). Agronomy 2021, 11, 794. [Google Scholar] [CrossRef]
- Cook, S.M.; Awmack, C.S.; Murray, D.A.; Williams, I.H. Are honey bees’ foraging preferences affected by pollen amino acid composition? Ecol. Entomol. 2003, 28, 622–627. [Google Scholar] [CrossRef]
- Paramás, A.M.; Bárez, J.A.; Marcos, C.C.; García-Villanova, R.J.; Sánchez, J.S. HPLC-fluorimetric method for analysis of amino acids in products of the hive (honey and bee-pollen). Food Chem. 2006, 95, 148–156. [Google Scholar] [CrossRef]
- Saa-Otero, M.P.; Diaz-Losada, E.; Fernandez-Gomez, E. Analysis of fatty acids, proteins and ethereal extract in honeybee pollen-considerations of their floral origin. Grana 2000, 39, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Estevinho, L.M.; Dias, T.; Anjos, O. Influence of the storage conditions (frozen vs. dried) in health-related lipid indexes and antioxidants of bee pollen. Eur. J. Lipid Sci. Technol 2019, 121, 1800393. [Google Scholar] [CrossRef] [Green Version]
- Feás, X.; Vázquez-Tato, M.P.; Estevinho, L.; Seijas, J.A.; Iglesias, A. Organic bee pollen: Botanical origin, nutritional value, bioactive compounds, antioxidant activity and microbiological quality. Molecules 2012, 17, 8359–8377. [Google Scholar] [CrossRef] [PubMed]
- Manning, R. Fatty acids in pollen: A review of their importance for honey bees. Bee World 2001, 82, 60–75. [Google Scholar] [CrossRef]
- Dahl, Å. Pollen lipids can play a role in allergic airway inflammation. Front. Immunol. 2018, 9, 2816. [Google Scholar] [CrossRef]
- Yang, K.; Wu, D.; Ye, X.; Liu, D.; Chen, J.; Sun, P.L. Characterization of chemical composition of bee pollen in China. J. Agric. Food Chem. 2013, 61, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Kostić, A.Ž.; Pešić, M.B.; Mosić, M.D.; Dojčinović, B.P.; Natić, M.M.; Trifković, J.Đ. Mineral content of bee pollen from Serbia. Arh. Hig. Rada Toksikol. 2015, 66, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fratini, F.; Turchi, B.; Michele, G.; Torracca, B.; Giusti, M.; Sagona, S.; Felicioli, A.; Cerri, D. Bee-gathered pollen loads suspension: Preliminary assessment of interaction with microbial growth for a potential employment as a natural food additive. J. Microbiol. Biotech. Food Sci. 2014, 3, 467–469. [Google Scholar]
- Mărgăoan, R.; Mărghitaş, L.A.; Dezmirean, D.S.; Dulf, F.V.; Bunea, A.; Socaci, S.A.; Bobiş, O. Predominant and secondary pollen botanical origins influence the carotenoid and fatty acid profile in fresh honeybee-collected pollen. J. Agric. Food Chem. 2014, 62, 6306–6316. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Q.; Wang, K.; Marcucci, M.C.; Sawaya, A.C.H.F.; Hu, L.; Xue, X.F.; Hu, F.L. Nutrient-rich bee pollen: A treasure trove of active natural metabolites. J. Funct. Foods 2018, 49, 472–484. [Google Scholar] [CrossRef]
- Barbieri, D.; Gabriele, M.; Summa, M.; Colosimo, R.; Leonardi, D.; Domenici, V.; Pucci, L. Antioxidant, nutraceutical properties, and fluorescence spectral profiles of bee pollen samples from different botanical origins. Antioxidants 2020, 9, 1001. [Google Scholar] [CrossRef]
- Reynolds, E.S. The use of lead citrate at high pH as an electron opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warakomska, Z. Badania nad wydajnością pyłkową roślin/Studies on the pollen efficiency of plants (in Polish). Pszczeln. Zesz. Nauk. 1972, 16, 63–90. [Google Scholar]
- Polish Standard PN-EN ISO 12966-1. Animal and Vegetable Fats and Oils. Gas Chromatography of Fatty Acis Methyl Esters. Part 1: Guidelines on Modern Gas Chromatography of Fatty Acid Methyl Esters (ISO 12966-1: 2014); ISO: Geneva, Switzerland, 2014. [Google Scholar]
- Wierciński, J. Przewodnik do Ćwiczeń z Instrumentalnej Analizy Chemicznych Składników Żywności. / Guidebook for Classes in Instrumental Analysis of Food Chemical Components for Students of Food Technology and Human Nutrition, 1st ed.; Wydawnictwo Akademii Rolniczej: Lublin, Poland, 1999; p. 137. (in Polish) [Google Scholar]
- Rabie, A.L.; Wells, J.D.; Dent, L.K. The nitrogen content of pollen protein. J. Apic. Res. 1983, 22, 119–123. [Google Scholar] [CrossRef]
- Davies, M.G.; Thomas, A.J. An investigation of hydrolytic techniques for the amino acid analysis of foodstuffs. J. Sci. Food Agric. 1973, 24, 1525–1540. [Google Scholar] [CrossRef]
- Gupta, C.; Dash, S.S. A new species of Rubus (Rosaceae) from Arunachal Pradesh, India. Blumea 2018, 63, 26–30. [Google Scholar] [CrossRef] [Green Version]
- San, A.M.; Linn, S.S.; Aye, S.M. Taxonomy and pollen morphology of the family Rosaceae found in the southern and nothern Shan state. J. Myanmar Acad. Arts Sci. 2019, 27, 189–210. [Google Scholar]
- Goldberg, R.B.; Beals, T.P.; Sanders, P.M. Anther development: Basic principles and practical applications. Plant Cell 1993, 5, 1217–1229. [Google Scholar] [CrossRef] [Green Version]
- Skrzydeł, J.; Borowska-Wykręt, D.; Kwiatkowska, D. Structure, assembly and function of cuticle from mechanical perspective with special focus on perianth. Int. J. Mol. Sci. 2021, 22, 4160. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wu, D.I.; Shi, J.; He, Y.I.; Pinot, F.; Grausem, B.; Yin, C.; Zhu, L.; Chen, M.; Luo, Z.; et al. Rice CYP703A3, a cytochrome P450 hydroxylase is essential for development of anther cuticle and pollen exine. J. Integr. Plant Biol. 2014, 56, 979–994. [Google Scholar] [CrossRef] [PubMed]
- Endress, P.K.; Stumpf, S. The diversity of stamen structures in ‘lower’ Rosidae (Rosales, Fabales, Proteales, Sapindales). Bot. J. Linn. Soc. 1991, 107, 217–293. [Google Scholar] [CrossRef]
- Nikolić, D.; Radović, A. The morphological properties of the flower and the percent of fertilised pistils of promising yellow fruiting raspberry hybrids. In International Scientific Symposium of Agriculture “AGROSYM Jahorina 2011”, Sarajevo, November 10–12 2011; Kovečević, D., Ed.; Faculty of Agriculture: East Sarajevo, Bosnia and Herzegovina, 2011; pp. 165–171. [Google Scholar]
- Orzeł, A.; Bieniasz, M.; Małodobry, M. Biology of flowering species of the genus Rubus as a preliminary study in primocane fruiting blackberries. In XI International Rubus and Ribes Symposium 1133; Fernandez, G.E., Hummer, K.E., Eds.; ISHS: Leuven, Belgium, 2016; pp. 171–176. [Google Scholar] [CrossRef]
- Kalkman, C. The Genus Rubus (Rosaceae) in Malesia. 2. The subgenus Micranthobatus. Blumea-Biodivers. Evol. Biogeogr. Plants 1984, 29, 319–386. [Google Scholar]
- Kalkman, C. The genus Rubus (Rosaceae) in Malesia. 3. The subgenus Micranthobatus. Blumea-Biodivers. Evol. Biogeogr. Plants 1987, 32, 323–341. [Google Scholar]
- Nybom, H. Pollen-limited seed set in pseudogamous blackberries (Rubus L. subgen. Rubus). Oecologia 1987, 72, 562–568. [Google Scholar] [CrossRef] [PubMed]
- Bean, A.R. Revision of Rubus subgenus Micranthobatus (Fritsch) Kalkman (Rosaceae) in Australia. Austrobaileya 1995, 4, 321–328. [Google Scholar]
- Hiregoudar, H.; Manju, D.; Nautiyal, B.P. Floral biology of wild raspberry (Rubus macilentus C.) species of Garhwal Himalaya, Uttarakhand, India. IJCS 2019, 7, 2200–2206. [Google Scholar]
- Gupta, C.; Dash, S.S. Rubus sengorensis (Rosaceae): A new record to India from Arunachal Pradesh. Nelumbo 2015, 57, 50–55. [Google Scholar] [CrossRef]
- Normasiwi, S.; Salamah, A.; Surya, M.I. Morphological characteristics of Indonesian Rubus flowers. Biodiversitas 2021, 22, 1441–1447. [Google Scholar] [CrossRef]
- Camarda, I. Rubus laconensis and Rubus pignatti (Rosaceae), two new species from Sardinia (Italy). Plant Biosyst. 2013, 147, 849–856. [Google Scholar] [CrossRef]
- Brown, A.O.; McNeil, J.N. Pollination ecology of the high latitude, dioecious cloudberry (Rubus chamaemorus; Rosaceae). Am. J. Bot. 2009, 96, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- Saffari, P.; Majd, A.; Jonoubi, P.; Najafi, F. Study on the reproductive organs development and embryological features of Agrimonia eupatoria L. (Rosaceae). Bot. Lett. 2021, 168, 270–282. [Google Scholar] [CrossRef]
- Raghavan, V. Anther and pollen development in rice (Oryza sativa). Am. J. Bot. 1988, 75, 183–196. [Google Scholar] [CrossRef]
- Jeffree, C.E. The fine structure of the plant cuticle. Annu. Plant Rev. 2006, 23, 11–125. [Google Scholar]
- Evert, R.F. Esau’s Plant Anatomy. Meristems, Cells and Tissues of the Plant Body-their Structure, Function and Development, 3rd ed.; John Wiely and Sons Inc.: Hoboken, NJ, USA, 2007; p. 624. [Google Scholar]
- De Carvalho, J.D.T.D.; Oliveira, J.M.S.D.; Freitas, C.C.D.; Martins, M.S. Stamen 1144 morphoanatomy of Dyckia Schult. f. (Bromeliaceae, Pitcairnioideae) species: New data 1145 for taxonomic use. Acta Bot. Bras. 2016, 30, 389–400. [Google Scholar] [CrossRef] [Green Version]
- Watson, L.; Dallwitz, M.J. The families of angiosperms: Automated descriptions with interactive identification and information retrieval. Aust. Syst. Bot. 1992, 4, 681–695. [Google Scholar] [CrossRef]
- Bhanwra, R.K.; Vij, S.P.; Chandel, V.; Dutt, R.; Dutt, S. Development of pollinium in two epidendroid orchids. Curr. Sci. 2006, 90, 1384–1388. [Google Scholar]
- Nelson, M.R.; Band, L.R.; Dyson, R.J.; Lessinnes, T.; Wells, D.M.; Yang, C.; Everitt, N.M.; Jensen, O.E.; Wilson, Z.A. A biomechanical model of anther opening reveals the roles of dehydration and secondary thickening. New Phytol. 2012, 196, 1030–1037. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.S.; Yao, C.; Xu, Q.L.; Sui, C.X.; Jia, X.L.; Hu, W.J.; Lv, Y.L.; Feng, Y.F.; Peng, Y.J.; Shen, S.Y.; et al. Development of the middle layer in the anther of Arabidopsis. Front. Plant Sci. 2021, 12, 634114. [Google Scholar] [CrossRef] [PubMed]
- Çetinbaş, A.; Ünal, M. Developmental and cytochemical features of male reproductive organ in Crataegus tanacetifolia (Lam). Pers. Not. Bot. Horti Agrobo. 2015, 43, 507–514. [Google Scholar] [CrossRef] [Green Version]
- Hufford, L.D.; Endress, P.K. The diversity of anther structures and dehiscence patterns among Hamamelididae. Bot. J. Linn. Soc. 1989, 99, 301–346. [Google Scholar] [CrossRef]
- D’Arcy, W.G. Anthers and stamens and what they do. In The Anther: Form. Function and Phylogeny, 1st ed.; D’Arcy, W.G., Keating, R.C., Eds.; Cambridge University Press: Cambridge, UK, 1996; pp. 1–24. [Google Scholar]
- Reinoso, H.; Luna, V.; Pharis, R.P.; Bottini, R. Dormancy in peach (Prunus persica) flower buds. V. Anatomy of bud development in relation to phenological stage. Can. J. Bot. 2002, 80, 656–663. [Google Scholar] [CrossRef]
- Manning, J.C. Diversity of endothecial patterns in angiosperms. In The Anther: Form, Function and Phylogeny, 1st ed.; D’Arcy, W.G., Keating, R.C., Eds.; Cambridge University Press: Cambridge, UK, 1996; pp. 136–158. [Google Scholar]
- Mitsuda, N.; Seki, M.; Shinozaki, K.; Ohme-Takagi, M. The NAC transcription 939 factors NST1 and NST2 of Arabidopsis regulate secondary wall thickenings and are 940 required for anther dehiscence. Plant Cell 2005, 17, 2993–3006. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Mao, Y.; Yang, J.; He, Y. TCP24 modulates secondary cell wall thickening and anther endothecium development. Front. Plant Sci. 2015, 6, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonner, L.; Dickinson, H. Anther dehiscence in Lycopersicon esculentum Mill. New Phytol. 1989, 113, 97–115. [Google Scholar] [CrossRef]
- De Craene, L.P.R. The evolutionary significance of homeosis in flowers: A morphological perspective. Int. J. Plant Sci. 2003, 164, S225–S235. [Google Scholar] [CrossRef]
- Percival, M.S. The presentation of pollen in certain angio-sperms and its collection by Apis Mellifera. New Phytol. 1955, 3, 353–368. [Google Scholar] [CrossRef]
- Denisow, B.; Antoń, S.; Szymczak, G. The flowering, pollen production, and insect visitors in the ornamental shrub Potentilla fruticosa L. (Rosaceae). J. Apic. Res. 2013, 57, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Masierowska, M. Early floral resources for urban bees from ornamental shrubs Ribes Aureum, Ribes Sanguineum and Staphylea Pinnata. J. Apic. Sci. 2020, 64, 309–320. [Google Scholar]
- Sawyer, R. Pollen Identification for Beekeepers, 1st ed.; Northern Bee Books: West Yorkshire, UK, 2006; p. 112. [Google Scholar]
- Wignall, V.R.; Arscott, N.A.; Nudds, H.E.; Squire, A.; Green, T.O.; Ratnieks, F.L. Thug life: Bramble (Rubus fruticosus L.) is a valuable foraging resource for honeybees and diverse flower-visiting insects. Insect Conserv. Divers. 2020, 13, 543–557. [Google Scholar] [CrossRef]
- Colbert, S.; De Oliveira, D. Influence of pollen variety on raspberry (Rubus idaeus L.) development. J. Hered. 1990, 81, 434–437. [Google Scholar] [CrossRef]
- Cane, J.H. Pollination potential of the bee Osmia aglaia for cultivated red raspberries and blackberries (Rubus: Rosaceae). HortScience 2005, 40, 1705–1708. [Google Scholar] [CrossRef] [Green Version]
- Tuohimetsä, S.; Hietaranta, T.; Uosukainen, M.; Kukkonen, S.; Karhu, S. Fruit development in artificially self-and cross-pollinated strawberries (Fragaria × ananassa) and raspberries (Rubus idaeus). Acta Agric. Scand. Soil Plant Sci. 2014, 64, 408–415. [Google Scholar] [CrossRef]
- Żurawicz, E.; Studnicki, M.; Kubik, J.; Pruski, K. A careful choice of compatible pollinizers significantly improves the size of fruits in red raspberry (Rubus idaeus L.). Sci. Hortic. 2018, 235, 253–257. [Google Scholar] [CrossRef]
- Lye, G.C.; Jennings, S.N.; Osborne, J.L.; Goulson, D. Impacts of the use of nonnative commercial bumble bees for pollinator supplementation in raspberry. J. Econ. Entomol. 2011, 104, 107–114. [Google Scholar] [CrossRef]
- Andrikopoulos, C.J.; Cane, J.H. Comparative pollination efficacies of five bee species on raspberry. J. Econ. Entomol. 2018, 111, 2513–2519. [Google Scholar] [CrossRef] [PubMed]
- Tasei, J.N.; Aupinel, P. Nutritive value of 15 single pollens and pollen mixes tested on larvae produced by bumblebee workers (Bombus terrestris, Hymenoptera: Apidae). Apidologie 2008, 39, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Gabriele, M.; Parri, E.; Felicioli, A.; Sagona, S.; Pozzo, L.; Domenici, V.; Pucci, L. Phytochemical composition and antioxidant activity of Tuscan bee pollen of different botanic origins. Ital. J. Food Sci. 2015, 27, 248–259. [Google Scholar] [CrossRef]
- Sagona, S.; Pozzo, L.; Peiretti, P.G.; Biondi, C.; Giusti, M.; Gabriele, M.; Pucci, L.; Felicioli, A. Palynological origin, chemical composition, lipid peroxidation and fatty acid profile of organic Tuscanian bee-pollen. J. Apic. Res. 2017, 56, 136–143. [Google Scholar] [CrossRef]
- Kleinschmidt, G.; Kondos, A. The influence of crude protein levels ion colony production. Austral. Beek. 1976, 78, 36–39. [Google Scholar]
- Alaux, C.; Ducloz, F.; Crauser, D.; Le Conte, Y. Wpływ diety na immunokompetencję pszczół. Biol. Łotysz 2010, 6, 562–565. [Google Scholar] [CrossRef] [Green Version]
- DeGrandi-Hoffman, G.; Chen, Y.; Huang, E.; Huang, M.H. The effect of diet on protein concentration, hypopharyngeal gland development and virus load in worker honey bees (Apis mellifera L.). J. Insect Physiol. 2010, 56, 1184–1191. [Google Scholar] [CrossRef]
- Amdam, G.V.; Norberg, K.; Hagen, A.; Omholt, S.W. Social exploitation of vitellogenin. Proc. Natl. Acad. Sci. USA 2003, 100, 1799–1802. [Google Scholar] [CrossRef] [Green Version]
- Koywiwattrakul, P.; Sittipraneed, S. Expression of vitellogenin and transferrin in activated ovaries of worker honey bees. Apis mellifera. Biochem. Genet. 2009, 47, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Bitondi, M.M.G.; Simões, Z.L.P. The relationship between level of pollen in the diet, vitellogenin and juvenile hormone titers in africanized Apis mellifera workers. J. Apic. Res. 1996, 35, 27–36. [Google Scholar] [CrossRef]
- Amdam, G.V.; Omholt, S.W. The regulatory anatomy of honeybee lifespan. J. Theor. Biol. 2002, 216, 209–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-García, C.; Heerman, M.C.; Cook, S.C.; Evans, J.D.; DeGrandi-Hoffman, G.; Banmeke, O.; Zhang, Y.; Huang, S.; Hamilton, M.; Chen, Y.P. Transferrin-mediated iron sequestration suggests a novel therapeutic strategy for controlling Nosema disease in the honey bee, Apis mellifera. PLoS Pathog. 2021, 17, e1009270. [Google Scholar] [CrossRef] [PubMed]
- Corona, M.; Velarde, R.A.; Remolina, S.; Moran-Lauter, A.; Wang, Y.; Hughes, K.A.; Robinson, G.E. Vitellogenin, juvenile hormone, insulin signaling, and queen honey bee longevity. Proc. Natl. Acad. Sci. USA 2007, 104, 7128–7133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, C.M.; Ihle, K.E.; Fondrk, M.K.; Page Jr, R.E.; Amdam, G.V. The gene vitellogenin has multiple coordinating effects on social organization. PLoS Biol. 2007, 5, 62. [Google Scholar] [CrossRef] [PubMed]
- Szczesna, T. Protein content and amino acids composition of bee-collected pollen originating from Poland, South Korea and China. J. Apic. Sci. 2006, 50, 91–99. [Google Scholar]
- Chwil, M. Struktura nektarników kwiatowych i wartość pszczelarska wybranych gatunków roślin z podrodziny Prunoideae (Rosaceae)/Structure of flower nectaries and beekeeping value of plant factors from the subfamily Prunoideae (Rosaceae). Habilitation Thesis, Wydawnictwo Uniwersytetu Przyrodniczego, Lublin, Poland, 2013; p. 108. (in Polish). [Google Scholar]
- Al-Kahtani, S.N.; Taha, E.K.; Khan, K.A.; Ansari, M.J.; Farag, S.A.; Shawer, D.; Elnabawy, E.M. Effect of harvest season on the nutritional value of bee pollen protein. PLoS ONE 2020, 15, e0241393. [Google Scholar] [CrossRef] [PubMed]
- Human, H.; Nicolson, S.W. Nutritional content of fresh, bee-collected and stored pollen of Aloe greatheadii var. davyana (Asphodelaceae). Phytochemistry 2006, 67, 1486–1492. [Google Scholar] [CrossRef] [Green Version]
- Block, K.P. Interactions among leucine, isoleucine, and valine with special reference to the branched-chain amino acid antagonism. In Absorption and Utilization of Amino Acids, 1st ed.; Friedman, M., Ed.; CRC Press: Boca Raton, FL, USA, 2019; pp. 229–244. [Google Scholar] [CrossRef]
- Green, C.R.; Wallace, M.; Divakaruni, A.S.; Phillips, S.A.; Murphy, A.N.; Ciaraldi, T.P.; Metallo, C.M. Branched-chain amino acid catabolism fuels adipocyte differentiation and lipogenesis. Nat. Chem. Biol. 2016, 12, 15–21. [Google Scholar] [CrossRef] [Green Version]
- McCormack, S.E.; Shaham, O.; McCarthy, M.A.; Deik, A.A.; Wang, T.J.; Gerszten, R.E.; Clish, C.B.; Mootha, V.K.; Grinspoon, S.K.; Fleischman, A. Circulating branched chain amino acid concentrations are associated with obesity and future insulin resistance in children and adolescents. Pediatr. Obes. 2013, 8, 52–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, D.; Richardson, N.E.; Green, C.L.; Spicer, A.B.; Murphy, M.E.; Flores, V.; Jang, C.; Kasza, I.; Nikodemova, M.; Wakai, M.H.; et al. The adverse metabolic effects of branched-chain amino acids are mediated by isoleucine and valine. Cell Metab. 2021, 33, 905–922. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.; Shafir, S.; Yehonatan, L.; Palmer, R.G.; Thornburg, R. A novel role for proline in plant floral nectars. Naturwissenschaffen 2006, 93, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Gualano, A.B.; Bozza, T.; Lopes De Campos, P.; Roschel, H.; Dos Santos Costa, A.; Luiz Marquezi, M.; Benatti, F.; Herbert Lancha Junior, A. Branched-chain amino acids supplementation enhances exercise capacity and lipid oxidation during endurance exercise after muscle glycogen depletion. J. Sports Med. Phys. Fitness. 2011, 51, 82–88. [Google Scholar] [PubMed]
- Melville, G.W.; Siegler, J.C.; Marshall, P.W. The effects of D-aspartic acid supplementation in resistance-trained men over a three month training period: A randomised controlled trial. PLoS ONE 2017, 12, e0182630. [Google Scholar] [CrossRef] [Green Version]
- Roshanzamir, F.; Safavi, S.M. The putative effects of D-aspartic acid on blood testosterone levels: A systematic review. Int. J. Reprod. Biomed. 2017, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Patri, M. Synaptic transmission and amino acid neurotransmitters. In Neurochemical Basis of Brain Function and Dysfunction, 1st ed.; Heinbocker, T., Tsoka, A.B., Eds.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Bauer, D.; Hamacher, K.; Bröer, S.; Pauleit, D.; Palm, C.; Zilles, K.; Coenen, H.; Langen, K.J. Preferred stereoselective brain uptake of d-serine-a modulator of glutamatergic neurotransmission. Nucl. Med. Biol. 2005, 32, 793–797. [Google Scholar] [CrossRef]
- Li, P.; Yin, Y.L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Bastings, J.J.; van Eijk, H.M.; Olde Damink, S.W.; Rensen, S.S. D-amino acids in health and disease: A focus on cancer. Nutrients 2019, 11, 2205. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, A.; Nishikawa, T.; Hayashi, T.; Fujii, N.; Harada, K.; Oka, T.; Takahashi, K. The presence of free D-serine in rat brain. Fed. Eur. Dioch. Soc. 1992, 296, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Mori, H.; Inoue, R. Serine racemase knockout mice. Chem. Biodivers. 2010, 7, 1573–1578. [Google Scholar] [CrossRef]
- Biancucci, M.; Mattioli, R.; Forlani, G.; Funck, D.; Costantino, P.; Trovato, M. Role of proline and GABA in sexual reproduction of angiosperms. Front. Plant Sci. 2015, 6, 680. [Google Scholar] [CrossRef] [Green Version]
- Mattioli, R.; Biancucci, M.; El Shall, A.; Mosca, L.; Costantino, P.; Funck, D.; Trovato, M. Proline synthesis in developing microspores is required for pollen development and fertility. BMC Plant Biol. 2018, 18, 356. [Google Scholar] [CrossRef] [Green Version]
- Kaur, G.; Asthir, B.J.B.P. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Mardirossian, M.; Sola, R.; Beckert, B.; Collis, D.W.; Di Stasi, A.; Armas, F.; Hilpert, H.; Wilson, D.N.; Scocchi, M. Proline rich peptides with improved antimicrobial activity against E. coli, K. pneumoniae, and A. baumannii. ChemMedChem 2019, 14, 2025–2033. [Google Scholar] [CrossRef] [Green Version]
- Bayram, N.E.; Gercek, Y.C.; Çelik, S.; Mayda, N.; Kostić, A.Ž.; Dramićanin, A.M.; Özkök, A. Phenolic and free amino acid profiles of bee bread and bee pollen with the same botanical origin-similarities and differences. Arab. J. Chem. 2021, 14, 103004. [Google Scholar] [CrossRef]
- Petersen, K.F.; Dufour, S.; Cline, G.W.; Shulman, G.I. Regulation of hepatic mitochondrial oxidation by glucose-alanine cycling during starvation in humans. J. Clin. Investig. 2019, 129, 4671–4675. [Google Scholar] [CrossRef] [PubMed]
- Leij-Halfwerk, S.; Dagnelie, P.C.; van den Berg, J.W.O.; Wattimena, J.D.L.; Hordijk-Luijk, C.H.; Wilson, J.P. Weight loss and elevated gluconeogenesis from alanine in lung cancer patients. Am. J. Clin. Nutr. 2000, 71, 583–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyake, Y.; Iwasaki, Y.; Terada, R.; Okamaoto, R.; Ikeda, H.; Makino, Y.; Kobashi, H.; Takaguchi, K.; Sakaguchi, K.; Shiratori, Y. Persistent elevation of serum alanine aminotransferase levels leads to poor survival and hepatocellular carcinoma development in type 1 autoimmune hepatitis. Aliment. Pharmacol. Ther. 2006, 24, 1197–1205. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.L.; Gong, N. Glycine and glycine receptor signaling in hippocampal neurons: Diversity, function and regulation. Prog. Neurobiol. 2010, 91, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torres, I.; María Zuniga-Munoz, A.; Guarner-Lans, V. Beneficial effects of the amino acid glycine. Mini. Rev. Med. Chem. 2017, 17, 15–32. [Google Scholar] [CrossRef]
- Razak, M.A.; Begum, P.S.; Viswanath, B.; Rajagopal, S. Multifarious beneficial effect of nonessential amino acid, glycine: A review. Oxid. Med. Cell. Longev. 2017, 718, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Brosnan, M.E.; Brosnan, J.T. Histidine metabolism and function. J. Nutr. 2020, 150, 2570S–2575S. [Google Scholar] [CrossRef] [PubMed]
- Ma, E.H.; Bantug, G.; Griss, T.; Condotta, S.; Johnson, R.M.; Samborska, B.; Mainolfi, N.; Suri, V.; Guak, H.; Balmer, M.L.; et al. Serine is an essential metabolite for effector T cell expansion. Cell Metab. 2017, 25, 345–357. [Google Scholar] [CrossRef]
- Xu, N.; Chen, G.; Liu, H. Antioxidative categorization of twenty amino acids based on experimental evaluation. Molecules 2017, 22, 2066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, M.L.; Lima-Cabello, E.; Alché, J.D.D.; Martínez-Rivas, J.M.; Castro, A.J. Lipid composition and associated gene expression patterns during pollen germination and pollen tube growth in olive (Olea europaea L.). Plant Cell. Physiol. 2020, 61, 1348–1364. [Google Scholar] [CrossRef]
- Anjos, O.; Paula, V.; Delgado, T.; Estevinho, L. Influence of the storage conditions on the quality of bee pollen. Zemdirbyste 2019, 106, 87–94. [Google Scholar] [CrossRef]
- Farah, J.; Choël, M.; De Nadaї, P.; Gosselin, S.; Petitprez, D.; Baroudi, M.; Visez, N. Extractable lipids from Phleum pratense pollen grains and their modifications by ozone exposure. Aerobiologia 2020, 36, 171–182. [Google Scholar] [CrossRef]
- Bashir, M.E.H.; Lui, J.H.; Palnivelu, R.; Naclerio, R.M.; Preuss, D. Pollen lipidomics: Lipid profiling exposes a notable diversity in 22 allergenic pollen and potential biomarkers of the allergic immune response. PLoS ONE 2013, 8, e57566. [Google Scholar] [CrossRef]
- Shakya, R.; Bhatla, S.C. A comparative analysis of the distribution and composition of lipidic constituents and associated enzymes in pollen and stigma of sunflower. Sex. Plant Reprod. 2010, 23, 163–172. [Google Scholar] [CrossRef]
- Manning, R.; Harvey, M. Fatty acids in honeybee-collected pollens from six endemic Western Australian eucalypts and the possible significance to the Western Australian beekeeping industry. Aust. J. Exp. Agric. 2002, 42, 217–223. [Google Scholar] [CrossRef]
- Leaf, A. The electrophysiologic basis for the antiarrhythmic and anticonvulsant effects of n−3 polyunsaturated fatty acids: Heart and brain. Lipids 2001, 36, S107–S110. [Google Scholar] [CrossRef] [PubMed]
- Morais, M.; Moreira, L.; Feás, X.; Estevinho, L.M. Honeybee-collected pollen from five Portuguese Natural Parks: Palynological origin, phenolic content, antioxidant properties and antimicrobial activity. Food Chem. Toxicol. 2011, 49, 1096–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, R.H.; Su, S.D.; Xiao, H.; Tian, H.Q. Calcium: A critical factor in pollen germination and tube elongation. Int. J. Mol. Sci. 2019, 20, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinhorst, L.; Kudla, J. Calcium-a central regulator of pollen germination and tube growth. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 1573–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, T.C.; Stephenson, A.G. Effects of soil phosphorus on pollen production, pollen size, pollen phosphorus content, and the ability to sire seeds in Cucurbita pepo (Cucurbitaceae). Sex. Plant Reprod. 1994, 7, 215–220. [Google Scholar] [CrossRef]
- Lankinen, Å. Effects of soil pH and phosphorus on in vitro pollen competitive ability and sporophytic traits in clones of Viola tricolor. Int. J. Plant Sci. 2000, 161, 885–893. [Google Scholar] [CrossRef]
- Zonia, L.; Cordeiro, S.; Tupý, J.; Feijó, J.A. Oscillatory chloride efflux at the pollen tube apex has a role in growth and cell volume regulation and is targeted by inositol 3, 4, 5, 6-tetrakisphosphate. Plant Cell 2002, 14, 2233–2249. [Google Scholar] [CrossRef] [Green Version]
- Matveyeva, N.P.; Andreyuk, D.S.; Yermakov, I.P. Transport of Cl– across the plasma membrane during pollen grain germination in tobacco. Biochemistry 2003, 68, 1247–1251. [Google Scholar] [CrossRef] [PubMed]
- Breygina, M.A.; Matveyeva, N.P.; Andreyuk, D.S.; Yermakov, I.P. Transmembrane transport of K+ and Cl− during pollen grain activation in vivo and in vitro. Russ. J. Dev. Biol. 2012, 43, 85–93. [Google Scholar] [CrossRef]
- Sawidis, T.; Baycu, G.; Weryszko-Chmielewska, E.; Sulborska, A. Impact of manganese on pollen germination and tube growth in lily. Acta Agrobot. 2021, 74, 1–17. [Google Scholar] [CrossRef]
- Li, L.G.; Sokolov, L.N.; Yang, Y.H.; Li, D.P.; Ting, J.; Pandy, G.K.; Luan, S. A mitochondrial magnesium transporter functions in Arabidopsis pollen development. Mol. Plant 2008, 1, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, L.G.; Liu, Z.H.; Yuan, Y.J.; Guo, L.L.; Mao, D.D.; Tian, L.F.; Chen, B.D.; Luan, S.; Li, D.P. Magnesium transporter AtMGT9 is essential for pollen development in Arabidopsis. Cell Res. 2009, 19, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.F.; Wang, B.; Lou, Y.; Han, W.J.; Lu, J.Y.; Li, D.D.; Li, L.G.; Zhu, J.; Yang, Z.N. Magnesium transporter 5 plays an important role in Mg transport for male gametophyte development in Arabidopsis. Plant J. 2015, 84, 925–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Huang, Y.; Tan, H.; Yang, X.; Tian, L.; Luan, S.; Chen, L.; Li, D. An endoplasmic reticulum magnesium transporter is essential for pollen development in Arabidopsis. Plant Sci. 2015, 231, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.F.; Qian, X.X.; Wang, K.Q.; Yu, Y.H.; Guo, Y.Y.; Zhao, X.; Wang, B.; Yang, N.Y.; Huang, J.R.; Yang, Z.N. Slowing development facilitates Arabidopsis mgt mutants to accumulate enough magnesium for pollen formation and fertility restoration. Front. Plant Sci. 2021, 11, 2299. [Google Scholar] [CrossRef]
- Morgano, M.A.; Martins, M.C.T.; Rabonato, L.C.; Milani, R.F.; Yotsuyanagi, K.; Rodriguez-Amaya, D.B. A comprehensive investigation of the mineral composition of brazilian bee pollen: Geographic and seasonal variations and contribution to human diet. J. Braz. Chem. Soc. 2012, 23, 727–736. [Google Scholar] [CrossRef] [Green Version]
- Brodschneider, R.; Crailsheim, K. Nutrition and health in honey bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Taha, E.K.A. Chemical composition and amounts of mineral elements in honeybee-collected pollen in relation to botanical origin. J. Apic. Sci. 2015, 59, 75–81. [Google Scholar] [CrossRef]
- Wright, G.A.; Nicolson, S.W.; Shafir, S. Nutritional physiology and ecology of honey bees. Annu. Rev. Entomol. 2018, 63, 327–344. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivars | Year | ||||||
|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | Mean for 2016–2018 | ||||
| Min.–Max. | Mean | Min.–Max. | Mean | Min.–Max. | Mean | ||
| Biennial fruiting cultivars | |||||||
| ‘Glen Ample’ | 89–107 | 97.56 ± 4.70 aBC | 92–111 | 102.38 ± 4.69 aAB | 99–125 | 107.31 ± 6.04 aAB | 102.42 ± 6.47 AB |
| ‘Laszka’ | 91–116 | 109.00 ± 6.75 aAB | 97–123 | 112.56 ± 7.77 aA | 107–127 | 117.94 ± 6.51 aA | 113.17 ± 7.82 A |
| ‘Radziejowa’ | 92–121 | 106.38 ± 8.39 aAB | 89–120 | 104.63 ± 7.75 aAB | 105–122 | 113.69 ± 5.10 aA | 108.23 ± 8.10 AB |
| Repeated fruiting cultivars | |||||||
| ‘Pokusa’ | 75–113 | 87.56 ± 10.01 bC | 96–113 | 104.75 ± 4.89 aAB | 96–110 | 103.31 ± 3.36 aB | 98.29 ± 10.01 AB |
| ‘Polana’ | 98–124 | 111.31 ± 7.50 aA | 101–126 | 109.63 ± 7.16 aAB | 100–123 | 115.81 ± 6.41 aA | 112.25 ± 7.38 A |
| ‘Polka’ | 88–97 | 92.56 ± 3.27 bC | 97–103 | 100.13 ± 2.00 aAB | 97–121 | 105.31 ± 8.44 aAB | 93.33 ± 7.44 B |
| Cultivars | Year | ||||||
|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | Mean for 2016–2018 | ||||
| Min.–Max. | Mean | Min.–Max. | Mean | Min.–Max. | Mean | ||
| Biennial fruiting cultivars | |||||||
| ‘Glen Ample’ | 4–6 | 4.97 ± 0.67 aA | 3–6 | 4.53 ± 0.69 aA | 3–6 | 4.77 ± 0.77 aA | 4.76 ± 0.73 A |
| ‘Laszka’ | 2–6 | 3.64 ± 0.79 aA | 2–5 | 3.27 ± 0.62 aA | 2–5 | 3.92 ± 0.78 aA | 3.5 ± 0.89 A |
| ‘Radziejowa’ | 2–6 | 3.86 ± 0.72 aA | 2–5 | 3.39 ± 0.79 aA | 2–6 | 3.97 ± 0.73 aA | 3.65 ± 0.89 A |
| Repeated fruiting cultivars | |||||||
| ‘Pokusa’ | 2–6 | 3.89 ± 0.75 aA | 2–5 | 3.50 ± 0.66 aA | 3–6 | 4.47 ± 0.82 aA | 3.92 ± 0.75 A |
| ‘Polana’ | 2–5 | 3.83 ± 0.67 aA | 2–5 | 3.69 ± 0.64 aA | 2–5 | 3.68 ± 0.75 aA | 3.73 ± 0.73 A |
| ‘Polka’ | 2–5 | 3.78 ± 0.63 aA | 2–5 | 4.04 ± 0.61 aA | 3–7 | 4.67 ± 0.66 aA | 4.06 ± 0.72 A |
| Cultivars | Study Year | ||||||
|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | Mean for 2016–2018 | ||||
| Min.–Max. | Mean | Min.–Max. | Mean | Min.–Max. | Mean | ||
| Biennial fruiting cultivars | |||||||
| ‘Glen Ample’ | 23.43–28.13 | 25.53 ± 2.39 aB | 26.37–31.97 | 29.45 ± 2.97 aAB | 26.08–32.16 | 28.96 ± 3.05 aAB | 28.08 ± 3.12 AB |
| ‘Laszka’ | 26.13–29.87 | 27.94 ± 1.87 bB | 30.05–33.84 | 32.27 ± 1.98 aA | 30.71–35.91 | 33.67 ± 2.67 aA | 31.29 ± 3.21 A |
| ‘Radziejowa’ | 29.86–32.63 | 31.27 ± 1.39 aA | 26.02–28.35 | 27.43 ± 1.24 bBC | 27.81–32.91 | 30.49 ± 2.56 abAB | 29.73 ± 2.36 AB |
| Repeated fruiting cultivars | |||||||
| ‘Pokusa’ | 25.89–29.91 | 27.90 ± 2.01 aAB | 27.31–33.92 | 30.65 ± 3.31 aAB | 26.04–31.34 | 29.45 ± 2.96 aAB | 29.34 ± 2.71 AB |
| ‘Polana’ | 23.53–26.93 | 24.93 ± 1.78 aB | 25.31–27.08 | 26.13 ± 0.89 aC | 26.31–28.02 | 27.32 ± 0.90 aB | 26.13 ± 1.67 B |
| ‘Polka’ | 25.42–27.03 | 26.46 ± 0.90 bB | 27.92–31.51 | 29.58 ± 1.81 aA | 24.96–28.32 | 27.05 ± 1.82 abB | 27.70 ± 1.98 AB |
| Fatty Acids | Biennial Fruiting Cultivars | Repeated Fruiting Cultivars | ||||
|---|---|---|---|---|---|---|
| ‘Glen Ample’ | ‘Laszka’ | ‘Radziejowa’ | ‘Pokusa’ | ‘Polana’ | ‘Polka’ | |
| Saturated fatty acids (SFA) | ||||||
| hexanoic acid (caproic acid) C6:0 | 0.18 ± 0.01 b | nd | nd | nd | nd | 0.89 ± 0.07 a |
| octanoic acid (caprylic acid) C8:0 | 0.15 ± 0.01 b | 0.64 ± 0.04 a | 0.07 ± 0.00 c | nd | nd | 0.03 ± 0.00 d |
| decanoic acid (capric acid) C10:0 | 0.10 ± 0.02 a | 0.06 ± 0.01 a | nd | nd | nd | nd |
| n-dodecanoic acid (lauric acid) C12:0 | 0.17 ± 0.02 c | 0.60 ± 0.08 a | 0.14 ± 0.02 c | 0.16 ± 0.02 c | 0.30 ± 0.04 b | nd |
| tetradecanoic acid (myristic acid) C14:0 | 0.88 ± 0.17 c | 1.73 ± 0.14 a | 0.73 ± 0.10c | 0.76 ± 0.08 c | 1.02 ± 0.20 b | 0.37 ± 0.05 d |
| pentadecanoic acid (pentadecylic acid) C15:0 | 0.22 ± 0.05 b | 0.33 ± 0.07 a | nd | nd | nd | nd |
| hexadecanoic acid (palmitic acid) C16:0 | 28.06 ± 0.84 d | 30.42 ± 1.29 cd | 31.50 ± 0.97 ab | 31.16 ± 1.08 bc | 32.98 ± 1.14 a | 26.25 ± 0.63 e |
| octadecanoic acid (stearic acid) C18:0 | 8.33 ± 0.36 a | 8.94 ± 0.25 a | 8.37 ± 0.54 a | 8.22 ± 0.23 a | 8.95 ± 0.47 a | 5.12 ± 0.28 d |
| eicosanoic a cid (arachidic acid) C20:0 | 0.43 ± 0.04 b | 1.00 ± 0.13 a | nd | nd | 0.57 | nd |
| docosanoic acid (behenic acid) C22:0 | 1.73 ± 0.24 b | 2.05 ± 0.41 a | 0.38 ± 0.08 d | 0.46 ± 0.06 d | 0.93 ± 0.04 c | nd |
| tetracosanoic acid (lignoceric acid) C24:0 | 0.44 ± 0.05 b | 0.61 ± 0.09 a | 0.36 ± 0.04 b | 0.19 ± 0.03 c | nd | nd |
| Monounsaturated fatty acid (MUFA) | ||||||
| cis -9-hexadecenoic acid (palmitoleic acid) C16:1n7 | 2.34 ± 0.23 b | 2.19 ± 0.15 b | 2.62 ± 0.55 b | 2.86 ± 0.67 b | 2.41 ± 0.30 b | 3.72 ± 0.45 a |
| is -9- octadecenoic acid (oleic acid) C18:1n9c + trans -9-octadecenoic acid (elaidic acid) C18:1n9t | 41.24 ± 1.52 a | 37.77 ± 1.74 b | 44.39 ± 2.07 a | 44.10 ± 2.94 a | 42.12 ± 2.38 a | 28.30 ± 1.90 c |
| Polyunsaturated fatty acid (PUFA) | ||||||
| cis -9,12-octadecadienoic acid (linoleic acid) C18:2n6c + trans- 9,12-octadecadienoic acid (linoelaidic acid) C18:2n6t | 3.81 ± 0.33 c | 4.56 ± 0.27 b | 5.32 ± 0.29 a | 5.52 ± 0.30 a | 3.24 ± 0.26 c | 3.40 ± 0.26 c |
| cis-9,12,15-octadecatrienoic acid (α-linolenic acid) C18:3n3(alpha) | 0.36 ± 0.07 b | 2.13 ± 0.35 a | nd | nd | nd | nd |
| cis -13,16-docosadienoic acid (docosadienoate) C22:2n6 | 0.44 ± 0.03 b | 0.61 ± 0.05 a | 0.44 ± 0.02 b | 0.24 ± 0.03 c | 0.48 ± 0.04 b | 0.71 ± 0.06 a |
| No. | Species/Cultivar | Number of Stamens/Flower | Author | No. | Species/Cultivar | Number of Stamens/Flower | Author |
|---|---|---|---|---|---|---|---|
| 1. | R. alceifolius | 160–230 | [76] | 28. | R. malvaceus | 100–200 | [76] |
| 2. | R. becardii | 115 | 29. | R. mearrsii | 60–90 | ||
| 3. | R. benguetensis | 50–80 | 30. | R. megacarpus | 30–185 | [77] | |
| 4. | R. chrysophyllus | 50–100 | 31. | R. moluccaanus | 30–185 | [76] | |
| 5. | R. cumingii | 60 | 32. | R. moorei | 40–50 | [79] | |
| 6. | R. diclinis | 18–40 | [77] | 33. | R. nebulosus | 45–60 | |
| 7. | R. divaricatus | 120 | [78] | 34. | R. nessensis | 77 | [78] |
| 8. | R. elongatus | 80–125 | [76] | 35. | R. novoguineensis | 14–20 | [77] |
| 9. | R. glomeratus | 24–120 | 36. | R. pedemontanus | 193 | [78] | |
| 10. | R. grabowskii | 171 | [78] | 37. | R. plicatus | 106 | |
| 11. | R. hartmanii | 105 | 38. | R. polyanthemus | 200 | ||
| 12. | R. heterosephalus | 70–90 | [76] | 39. | R. pyramidalis | 172 | |
| 13 | R. idaeus ‘Glen Ample’ | 102 | [PS] | 40. | R. pyrifolius | 40–100 | [76] |
| 14. | R. idaeus ‘Laszka’ | 113 | 41. | R. radula | 134 | [78] | |
| 15. | R. idaeus ‘Meeker’ | 88–93 | [74] | 42. | R. rolfei | 120–140 | [76] |
| 43. | R. royenii | 40–60 | [77] | ||||
| 16. | R. idaeus ‘Pokusa’ | 98 | [PS] | 44. | R. royenii ‘Hispidus’ | 14–20 | |
| 17. | R. idaeus ‘Polana’ | 112 | 45. | R. royenii ‘Ikilimbu’ | 45–60 | ||
| 18. | R. idaeus ‘Polka’ | 93 | 46. | R. royenii ‘Royenii’ | 20–40 | ||
| 83–88 | [75] | ||||||
| 19. | R. idaeus ‘Radziejowa’ | 108 | [PS] | 47. | R. scissus | 47 | |
| 20. | R. infestus | 102 | [78] | 48. | R. sengorensis | 20 | [81] |
| 21. | R. insularis | 205 | 49. | R. sosogenensis | 100 | [76] | |
| 22. | R. keleterios | 85–125 | [76] | 50. | R. sprengelii | 125 | [78] |
| 23 | R. laciniatus | 152 | [78] | 51. | R. sundaicus | 65 | [76] |
| 24. | R. lidebergii | 172 | 52. | R. vestitus | 217 | [78] | |
| 25. | R. lineatus | 50–150 | [76] | 53. | R. vigorosus | 132 | |
| 26. | R. luzonenesis | 50–100 | 54. | R. wahlbergii | 170 | ||
| 27. | R. macilentus | 60–90 | [80] | 55. | Rubus subgenus Rubus Watson ‘Gazda’ | 103–125 | [75] |
| No. | Genus/Species/Cultivar | Length | Height | Author | |
|---|---|---|---|---|---|
| Filament | Anther | Stamen | |||
| 1. | R. alcelifolius | nd | nd | 4.0 | [82] |
| 2. | R. benguetensis | 5 | 0.5–0.8 | 5.5–5.8 | [76] |
| 3. | R. chamaemorus | nd | nd | 4.0 | [84] |
| 4. | R. chrysophyllus | nd | nd | 2.2 | [82] |
| 5. | R. clementis | 2.5 | 1–1.5 | 3.5–4.0 | [77] |
| 6. | R. cordiformis | 7 | 1–1.5 | 8–8.5 | |
| 7. | R. diclinis | 4.5 | 0.6–1.0 | 5.1–5.5 | |
| 8. | R. ellipticus | nd | nd | 2.7 | [82] |
| 9. | R. elongatus | 2–2.5 | 0.8–2.3 | 2.8–4.8 | [76] |
| 10. | R. fraxinifolius | nd | nd | 1.9 | [82] |
| 11. | R. idaeus ‘Polka’ | nd | nd | 3.6–6.9 | [75] |
| 12. | R. laconensis | nd | nd | 6–10 | [83] |
| 13. | R. lineatus | 4 | 0.8–1 | 4.8–5.0 | [76] |
| nd | nd | 1.4 | [83] | ||
| 14. | R. luzoniensis | 4.5 | 0.5 | 5.0 | [76] |
| 15. | R. macilentus | nd | nd | 3.2 | [80] |
| 16. | R. megacarpus | 10 | 2 | 12 | [77] |
| 17. | R. moluccanus | nd | nd | 3.0 | [82] |
| 18. | R. moorei | 2.5–3.5 | 0.5–1.5 | 3.0–5.0 | [79] |
| 19. | R. nebulosus | 2.5–3.5 | 0.5–1.5 | 3.0–5.0 | |
| 20. | R. novoguineensis | 2.5 | 1 | 3.5 | [77] |
| 21. | R. pyrifolius | 6.5 | 0.5–0.8 | 7.0–7.3 | [76] |
| nd | nd | 4.8 | [82] | ||
| 22. | R. rosifolius | nd | nd | 2.5 | |
| 23. | R. royenii ‘Hispidus’ | 1.5 | 1 | 2.5 | [77] |
| 24. | R. royenii ‘Ikilimbu | 2.5 | 1.2-1.5 | 3.7–4.0 | |
| 25. | R. royenii ‘Royenii’ | 2.5 | 1 | 3.5 | |
| 26. | R. saxatilis ‘Gazda’ | nd | nd | 5.9–6.9 | [75] |
| 27. | R. sengorensis | nd | nd | 1–2 | [81] |
| 28. | R. smithii | 3 | 1.5–2.0 | 4.5–5.0 | [76] |
| 29 | R. sorsogonensis | 8 | 0.8 | 8.8 | |
| 30. | R. sundaicus | 4.5 | 0.3–0.8 | 4.8–5.3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kostryco, M.; Chwil, M. Structure of Anther Epidermis and Endothecium, Production of Pollen, and Content of Selected Nutrients in Pollen Grains from Six Rubus idaeus L. Cultivars. Agronomy 2021, 11, 1723. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091723
Kostryco M, Chwil M. Structure of Anther Epidermis and Endothecium, Production of Pollen, and Content of Selected Nutrients in Pollen Grains from Six Rubus idaeus L. Cultivars. Agronomy. 2021; 11(9):1723. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091723
Chicago/Turabian StyleKostryco, Mikołaj, and Mirosława Chwil. 2021. "Structure of Anther Epidermis and Endothecium, Production of Pollen, and Content of Selected Nutrients in Pollen Grains from Six Rubus idaeus L. Cultivars" Agronomy 11, no. 9: 1723. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091723