
The Distribution of Soil Micro-Nutrients and the Effects on Herbage Micro-Nutrient Uptake and Yield in Three Different Pasture Systems

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
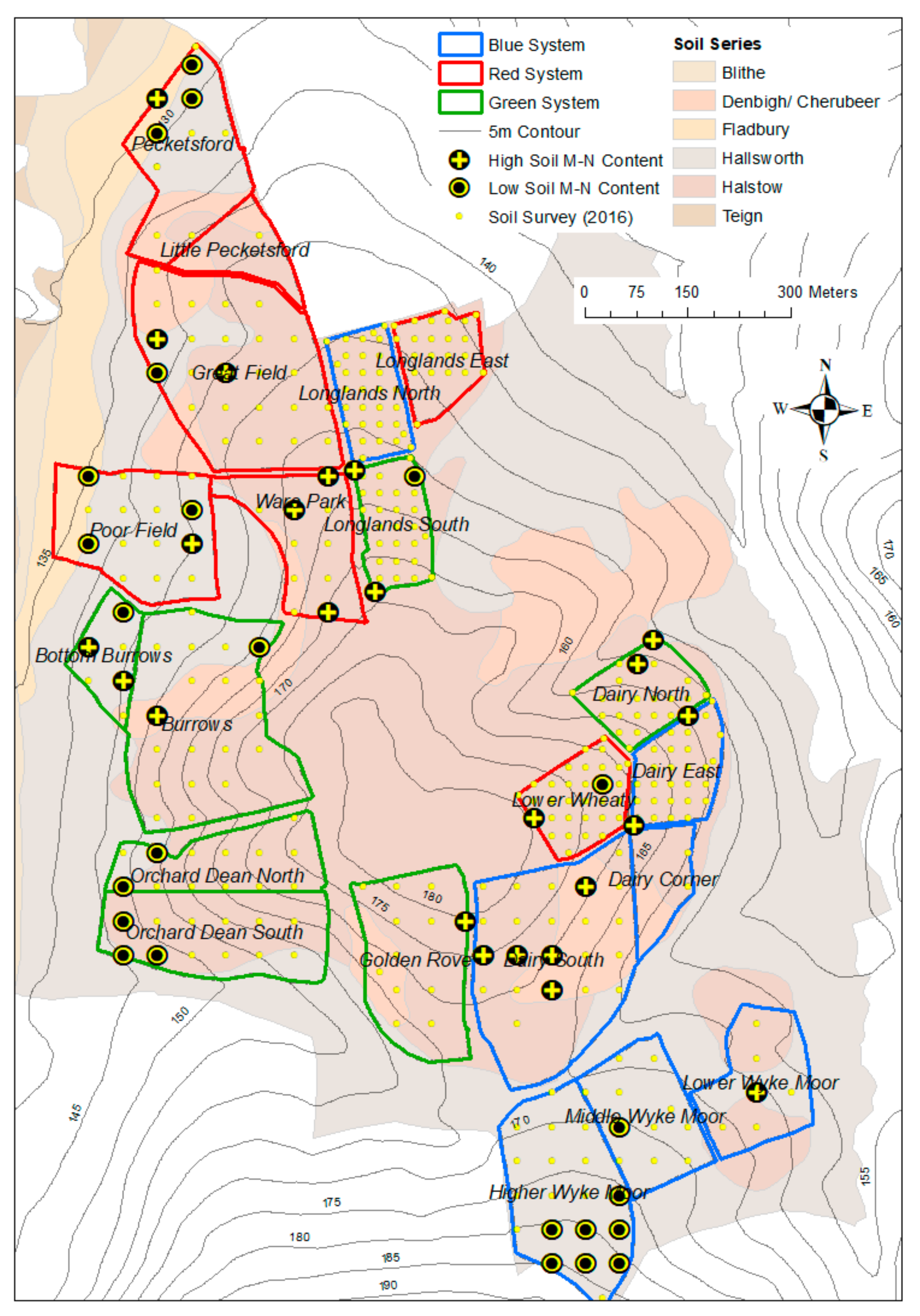
2.1. Site Description
2.2. Pasture System Treatments and Flora
- The green/P + N system continues to represent permanent pasture, predominantly composed of perennial ryegrass (Lolium perenne) with some unsown grass, legume and forb species, containing on average 64% Lolium perenne, 38% Agrostis stolonifera, L., 2% Holcus lanatus (L.) and 1% Alopecurus geniculatus (L.) as the main constituents. This system receives N fertilizer at a standard rate. None of the seven fields have been ploughed for at least 20 years.
- The blue/HSG + L system represents a sward improved with a perennial ryegrass of an innovative high-sugar variety and white clover mix (Lolium perenne cv. AberMagic + cv. AberHerald or Festulolium cv. Prior + AberHerald), targeting 30% ground cover by the latter. No N fertilizer is used (except rarely, described below) due to clover’s atmospheric N fixation.
- The red/HSG + N system represents a sward improved with a perennial ryegrass of an innovative high-sugar variety (Lolium perenne cv. AberMagic). This system receives N fertilizer at a standard rate.
2.3. Soil Survey and Soil Sample Analysis
2.4. Herbage Sampling
2.5. Herbage Analysis
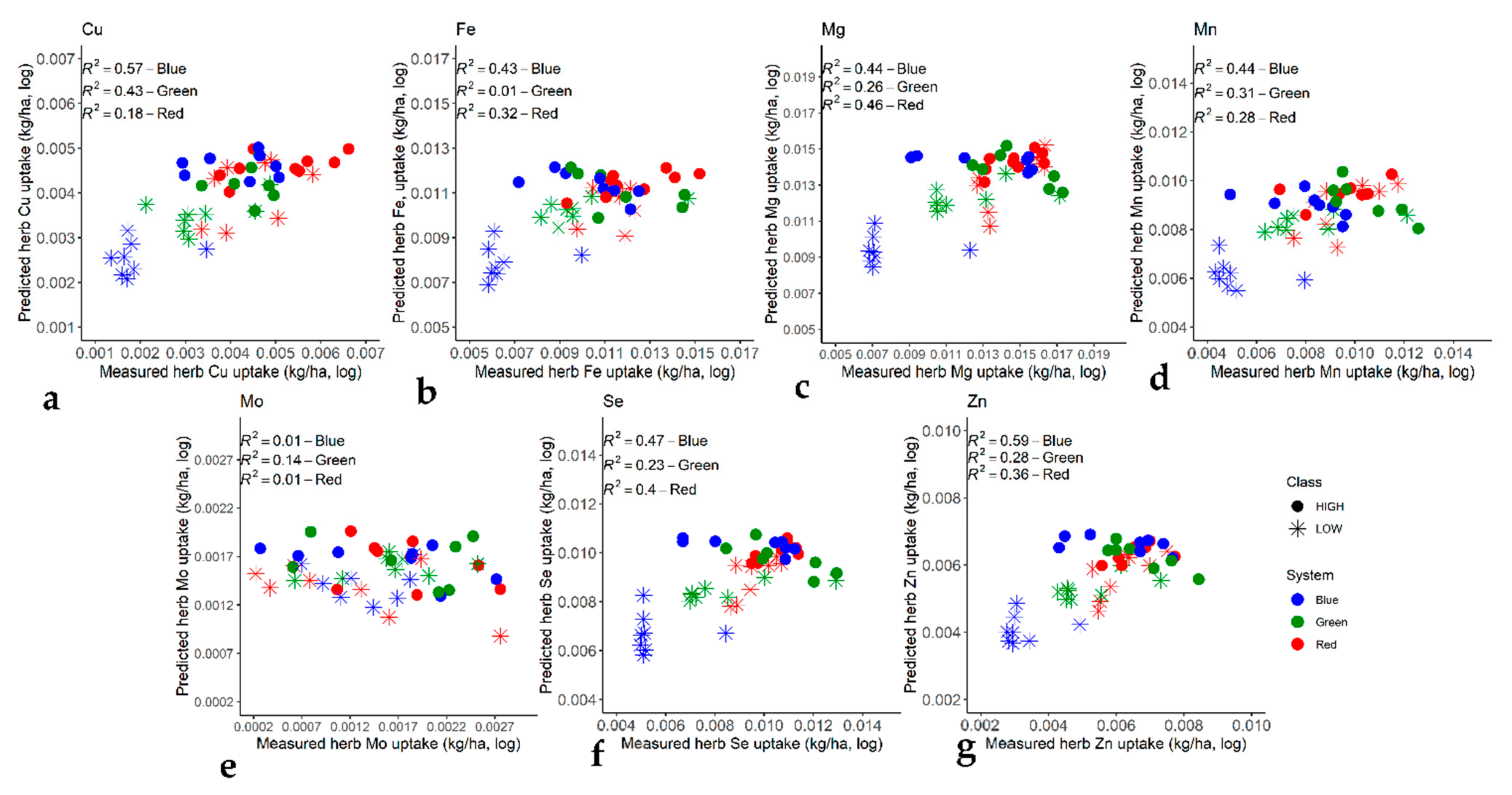
2.6. Prediction of Micro-Nutrient Content with Machine Learning and Statistical Analyses
3. Results
3.1. Site and Soil Properties in the Pasture Systems and at Locations Classed as High and Low Soil Micro-Nutrient Content
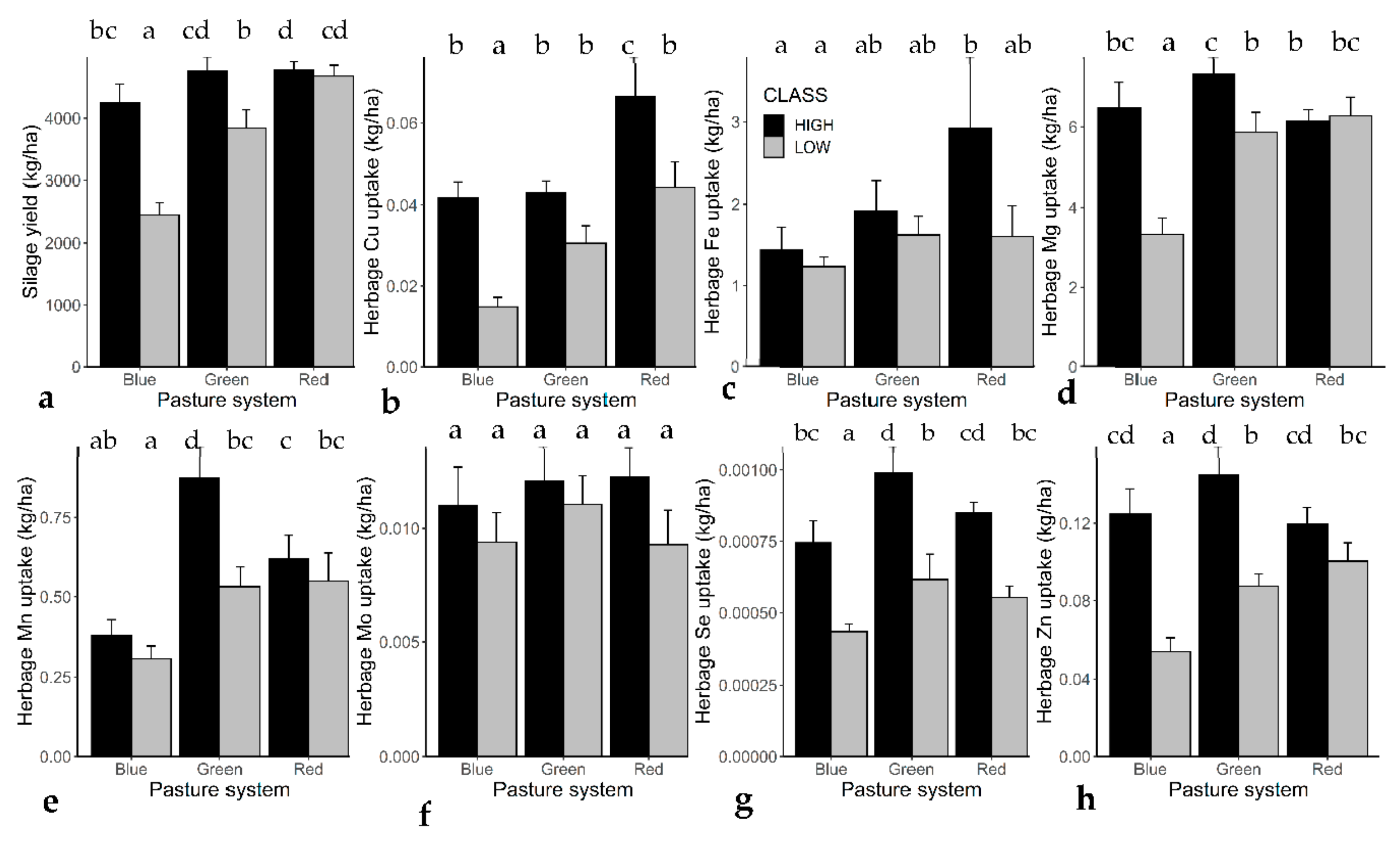
3.2. Herbage Micro-Nutrient Concentrations
3.3. Herbage Yield and Micro-Nutrient Uptake
4. Discussion
4.1. Site and Soil Properties Explaining Soil Micro-Nutrient Concentrations
4.2. Herbage Micro-Nutrient Concentrations
4.3. Effects of Soil Micro-Nutrient Concentrations on Yield
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arnott, G.; Ferris, C.; O’Connell, N. A Comparison of Confinement and Pasture Systems for Dairy Cows: What Does the Science Say? AgriSearch: Hillsborough, UK, 2015. [Google Scholar]
- Wilkinson, J.M.; Lee, M.R.F.; Rivero, M.J.; Chamberlain, A.T. Some challenges and opportunities for grazing dairy cows on temperate pastures. Grass Forage Sci. 2020, 75, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.B.; Tracy, B.F. Evaluating seasonal variation in mineral concentration of cool-season pasture herbage. Grass Forage Sci. 2013, 70, 94–101. [Google Scholar] [CrossRef]
- Orjales, I.; Latorre, C.H.; Miranda, M.; Rey-Crespo, F.; Rodríguez-Bermúdez, R.; López-Alonso, M. Evaluation of trace element status of organic dairy cattle. Animal 2018, 12, 1296–1305. [Google Scholar] [CrossRef]
- Govasmark, E.; Steen, A.; Bakken, A.K.; Strøm, T.; Hansen, S. Factors affecting the concentration of Zn, Fe and Mn in herbage from organic farms and in relation to dietary requirements of ruminants. Acta Agric. Scand. Sect. B Plant Soil Sci. 2005, 55, 131–142. [Google Scholar] [CrossRef]
- Khan, Z.I.; Ashraf, M.; Hussain, A. Evaluation of macro mineral contents of forages: Influence of pasture and seasonal variation. Asian-Australas. J. Anim. Sci. 2007, 20, 908–913. [Google Scholar] [CrossRef]
- Kingston-Smith, A.; Marshall, A.; Moorby, J. Breeding for genetic improvement of forage plants in relation to increasing animal production with reduced environmental footprint. Animal 2013, 7, 79–88. [Google Scholar] [CrossRef]
- Medeiros, R.B.; Steiner, J.J. White clover seed production: III. Cultivar differences under contrasting management practices. Crop. Sci. 2000, 40, 1317–1324. [Google Scholar] [CrossRef]
- Edmeades, D.C. The magnesium requirements of pastures in New Zealand: A review. N. Z. J. Agric. Res. 2004, 47, 363–380. [Google Scholar] [CrossRef]
- Sherrell, C.; Metherell, A. Diagnosis and treatment of molybdenum deficiency in pastures. Proc. N. Z. Grassl. Assoc. 1986, 47, 203–209. [Google Scholar] [CrossRef]
- Zaman, M.; Zaman, S.; Quin, B.; Kurepin, L.; Shaheen, S.; Nawaz, S.; Dawar, K. Improving pasture growth and urea efficiency using N inhibitor, molybdenum and elemental sulphur. J. Soil Sci. Plant Nutr. 2014, 14, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Rengel, Z. Availability of Mn, Zn and Fe in the rhizosphere. J. Soil Sci. Plant Nutr. 2015, 15, 397–409. [Google Scholar] [CrossRef] [Green Version]
- Kao, P.; Darch, T.; McGrath, S.; Kendall, N.; Buss, H.; Warren, H.; Lee, M. Factors influencing elemental micronutrient supply from pasture systems for grazing ruminants. Adv. Agron. 2020, 164, 161–229. [Google Scholar] [CrossRef]
- Stevenson, F.J. Organic Matter-Micronutrient Reactions in Soil. In Micronutrients in Agriculture; Mortvedt, J., Ed.; Soil Science Society of America American Society of Agronomy: Madison, WI, USA, 2018; Volume 4, pp. 145–186. [Google Scholar]
- Dhaliwal, S.; Naresh, R.; Mandal, A.; Singh, R.; Dhaliwal, M. Dynamics and transformations of micronutrients in agricultural soils as influenced by organic matter build-up: A review. Environ. Sustain. Indic. 2019, 1–2, 100007. [Google Scholar] [CrossRef]
- Temminghoff, E.J.M.; Van Der Zee, S.E.A.T.M.; De Haan, F.A.M. Effects of dissolved organic matter on the mobility of copper in a contaminated sandy soil. Eur. J. Soil Sci. 1998, 49, 617–628. [Google Scholar] [CrossRef]
- Mesquita, M.E.; Carranca, C. Effect of dissolved organic matter on copper—zinc competitive adsorption by a sandy soil at different pH values. Environ. Technol. 2005, 26, 1065–1072. [Google Scholar] [CrossRef]
- Øgaard, A.F.; Sogn, T.A.; Eich-Greatorex, S. Effect of cattle manure on selenate and selenite retention in soil. Nutr. Cycl. Agroecosyst. 2006, 76, 39–48. [Google Scholar] [CrossRef]
- McGrath, S.; Micó, C.; Curdy, R.; Zhao, F.-J. Predicting molybdenum toxicity to higher plants: Influence of soil properties. Environ. Pollut. 2010, 158, 3095–3102. [Google Scholar] [CrossRef]
- Williams, R.J.B.; Stojkovska, A.; Cooke, G.W.; Widdowson, F.V. Effects of fertilisers and farmyard manure on the copper, manganese, molybdenum and zinc removed by arable crops at Rothamsted. J. Sci. Food Agric. 1960, 11, 570–575. [Google Scholar] [CrossRef]
- Thomas, C.L.; Acquah, G.E.; Whitmore, A.P.; McGrath, S.P.; Haefele, S.M. The effect of different organic fertilizers on yield and soil and crop nutrient concentrations. Agronomy 2019, 9, 776. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.H.; Haynes, R.J. Influence of improved pastures and grazing animals on nutrient cycling within New Zealand soils. N. Z. J. Ecol. 1990, 14, 49–57. [Google Scholar]
- Higgins, S.; Schellberg, J.; Bailey, J. Improving productivity and increasing the efficiency of soil nutrient management on grassland farms in the UK and Ireland using precision agriculture technology. Eur. J. Agron. 2019, 106, 67–74. [Google Scholar] [CrossRef]
- Pirhofer-Walzl, K.; Søegaard, K.; Jensen, H.H.; Eriksen, J.; Sanderson, M.A.; Rasmussen, J. Forage herbs improve mineral composition of grassland herbage. Grass Forage Sci. 2011, 66, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, P.W.; Humphreys, M.O. Progress in breeding perennial forage grasses for temperate agriculture. J. Agric. Sci. 2003, 140, 129–150. [Google Scholar] [CrossRef]
- McDonagh, J.; O’Donovan, M.; McEvoy, M.; Gilliland, T.J. Genetic gain in perennial ryegrass (Lolium perenne) varieties 1973 to 2013. Euphytica 2016, 212, 187–199. [Google Scholar] [CrossRef] [Green Version]
- DEFRA (Department for Environment, Food and Rural Affairs). Farming Statistics Final Crop Areas, Yields, Livestock Populations and Agricultural Workforce at June 2019—United Kingdom; DEFRA: London, UK, 2019. Available online: www.statistics.gov.uk (accessed on 2 March 2021).
- Shalloo, L.; Creighton, P.; O’Donovan, M. The economics of reseeding on a dairy farm. Ir. J. Agric. Food Res. 2011, 50, 113–122. [Google Scholar]
- Rivero, M.J.; Orr, R.J.; Coumbe, J.; Willson, B.; Evans, J.; Lee, M.R.F. Replacing a permanent pasture and its effect in the first year on a commercial dairy farm. Agro Sur. 2018, 46, 3–12. Available online: http://agrosur-journal.cl/agrosur/wp-content/uploads/2018/09/art02-Rivero-PUBLICAR.pdf (accessed on 2 March 2021).
- Blackwell, M.S.; Jarvis, S.C.; Wilkins, R.J.; Beaumont, D.A.; Cardenas, L.M.; Chadwick, D.R.; Collins, A.L.; Dungait, J.A.; Gibb, M.J.; Hopkins, A.; et al. The importance of sustained grassland and environmental research: A case study from North Wyke research station, UK, 1982–2017. Adv. Agron. 2018, 149, 161–235. [Google Scholar] [CrossRef] [Green Version]
- Agriculture and Horticulture Development Board (AHDB). Improving Pasture for Better Returns. 2018. Available online: https://ahdb.org.uk/knowledge-library/improving-pasture-for-better-returns (accessed on 5 January 2021).
- Kohmann, M.M.; Sollenberger, L.E.; Dubeux, J.C.B.; Silveira, M.L.; Moreno, L.S.B.; da Silva, L.S.; Aryal, P.; da Silva, L.S. Nitrogen fertilization and proportion of legume affect litter decomposition and nutrient return in grass pastures. Crop. Sci. 2018, 58, 2138–2148. [Google Scholar] [CrossRef]
- Høgh-Jensen, H.; Søegaard, K. Robustness in the mineral supply from temporary grasslands. Acta Agric. Scand. Sect. B Plant Soil Sci. 2012, 62, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Grigg, D.; Grigg, J.; Lucas, R. Maximising subterranean clover in Marlborough’s hill country is key to weaning 80% of sale lambs prime. Proc. N. Z. Grassl. Assoc. 2008, 70, 25–29. [Google Scholar] [CrossRef]
- Fraser, T.; Rowarth, J. Legumes, herbs or grass for lamb performance? Proc. N. Z. Grassl. Assoc. 1996, 58, 49–52. [Google Scholar] [CrossRef]
- Egan, M.; Galvin, N.; Hennessy, D. Incorporating white clover (Trifolium repens L.) into perennial ryegrass (Lolium perenne L.) swards receiving varying levels of nitrogen fertilizer: Effects on milk and herbage production. J. Dairy Sci. 2018, 101, 3412–3427. [Google Scholar] [CrossRef] [PubMed]
- Penrose, B.; Lovatt, J.A.; Palmer, S.; Thomson, R.; Broadley, M.R. Revisiting variation in leaf magnesium concentrations in forage grasses for improved animal health. Plant Soil 2020, 457, 43–55. [Google Scholar] [CrossRef]
- Orr, R.J.; Griffith, B.A.; Rose, S.; Hatch, D.J.; Hawkins, J.M.B.; Murray, P.J. Designing and creating the North Wyke Farm Platform. In Catchment Science; Haygarth, P., Jordan, P., Harris, R., Eds.; BBSRC: Dublin, Ireland, 2011; p. 35. [Google Scholar]
- Orr, R.J.; Murray, P.J.; Eyles, C.J.; Blackwell, M.; Cardenas, L.M.; Collins, A.; Dungait, J.; Goulding, K.; Griffith, B.A.; Gurr, S.J.; et al. The North Wyke farm platform: Effect of temperate grassland farming systems on soil moisture contents, runoff and associated water quality dynamics. Eur. J. Soil Sci. 2016, 67, 374–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orr, R.J.; Griffith, B.A.; Rivero, M.J.; Lee, M.R.F. Livestock performance for sheep and cattle grazing lowland permanent pasture: Benchmarking potential of forage-based systems. Agronomy 2019, 9, 101. [Google Scholar] [CrossRef] [Green Version]
- Harrod, T.R.; Hogan, D.V. The Soils of North Wyke and Rowden. 2008. Available online: http://www.rothamsted.ac.uk/sites/default/files/SoilsNWRowden.pdf (accessed on 29 March 2016).
- Mückenhausen, E.; Avery, B.W. Soil classification for England and Wales (higher categories). Soil survey, technical monograph No. 14, rothamsted experimental station, Harpenden/England 1980. J. Plant Nutr. Soil Sci. 1981, 144, 228. [Google Scholar] [CrossRef]
- DEFRA (Department for Environment, Food and Rural Affairs). The Fertilizer Manual (RB209), 8th ed.; The Stationary Office: Norwich, UK, 2010. [Google Scholar]
- Harris, P.; Darch, T.; Lu, B.; Comber, A.; Lee, M.R.F. Investigation of the Multivariate Structure of a Soils Micronutrients Data Set at the Farm-Scale; Spatial Accuracy: Beijing, China, 2018. [Google Scholar]
- McGrath, S.; Cunliffe, C.H. A simplified method for the extraction of the metals Fe, Zn, Cu, Ni, Cd, Pb, Cr, Co and Mn from soils and sewage sludges. J. Sci. Food Agric. 1985, 36, 794–798. [Google Scholar] [CrossRef]
- Hawkins, J.; Harris, P. User Guide to NWFP Field Survey Data. 2021. Available online: http://resources.rothamsted.ac.uk/sites/default/files/groups/North_Wyke_Farm_Platform/FP_UG.Doc_.003_FieldSurveyData_ver1.5.pdf (accessed on 22 December 2020).
- Zarcinas, B.A.; Cartwright, B.; Spouncer, L.R. Nitric acid digestion and multi-element analysis of plant material by inductively coupled plasma spectrometry. Commun. Soil Sci. Plant Anal. 1987, 18, 131–146. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Machine Learning; Springer: Berlin/Heidelberg, Germany, 2001; Volume 45, pp. 5–32. [Google Scholar]
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Olea, R.A. Declustering of clustered preferential sampling for histogram and semivariogram inference. Math. Geol. 2007, 39, 453–467. [Google Scholar] [CrossRef]
- Diggle, P.J.; Menezes, R.; Su, T.-L. Geostatistical inference under preferential sampling. J. R. Stat. Soc. Ser. C Appl. Stat. 2010, 59, 191–232. [Google Scholar] [CrossRef]
- Granger, S.J.; Harris, P.; Peukert, S.; Guo, R.; Tamburini, F.; Blackwell, M.; Howden, N.J.; McGrath, S. Phosphate stable oxygen isotope variability within a temperate agricultural soil. Geoderma 2017, 285, 64–75. [Google Scholar] [CrossRef] [Green Version]
- Wright, M.N.; Ziegler, A. Ranger: A fast implementation of random forests for high dimensional data in C++ and R. J. Stat. Softw. 2017, 77. [Google Scholar] [CrossRef] [Green Version]
- Deutsch, C.V.; Journel, A.G. GSLIB: Geostatistical Software Library and User’s Guide; Oxford University Press: New York, NY, USA, 1998. [Google Scholar]
- McAuliffe, G.; López-Aizpún, M.; Blackwell, M.; Castellano-Hinojosa, A.; Darch, T.; Evans, J.; Horrocks, C.; Le Cocq, K.; Takahashi, T.; Harris, P.; et al. Elucidating three-way interactions between soil, pasture and animals that regulate nitrous oxide emissions from temperate grazing systems. Agric. Ecosyst. Environ. 2020, 300, 106978. [Google Scholar] [CrossRef]
- Woledge, J.; Suarez, A.C. The growth and photosynthesis of seedling plants of white clover at low temperature. Ann. Bot. 1983, 52, 239–245. [Google Scholar] [CrossRef]
- Dennis, W.D.; Woledge, J. The effect of nitrogenous fertilizer on the photosynthesis and growth of white clover/perennial ryegrass swards. Ann. Bot. 1985, 55, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Reddy, K.J.; Munn, L.C.; Wang, L. Chemistry and Mineralogy of Molybdenum in Soils. In Molybdenum in Agriculture; Gupta, U.C., Ed.; Cambridge University Press: Cambridge, UK, 1997; pp. 4–22. [Google Scholar]
- Marks, J.A.; Perakis, S.S.; King, E.; Pett-Ridge, J. Soil organic matter regulates molybdenum storage and mobility in forests. Biogeochemistry 2015, 125, 167–183. [Google Scholar] [CrossRef]
- McGrath, S.; Micó, C.; Zhao, F.; Stroud, J.; Zhang, H.; Fozard, S. Predicting molybdenum toxicity to higher plants: Estimation of toxicity threshold values. Environ. Pollut. 2010, 158, 3085–3094. [Google Scholar] [CrossRef]
- Axelson, U.; Söderström, M.; Jonsson, A. Risk assessment of high concentrations of molybdenum in forage. Environ. Geochem. Health 2018, 40, 2685–2694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes, J.O.; Bendezú, S.G.; Joaquín, A.H. Molybdenum and cobalt application in bean (Phaseolus vulgaris L.) with two fertilization systems under no-tillage. J. Agric. Sci. Technol. B 2016, 6, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Abreu, I.; Mihelj, P.; Raimunda, D. Transition metal transporters in rhizobia: Tuning the inorganic micronutrient requirements to different living styles. Metallomics 2019, 11, 735–755. [Google Scholar] [CrossRef]
- Alam, F.; Kim, T.Y.; Kim, S.Y.; Alam, S.S.; Pramanik, P.; Kim, P.J.; Lee, Y.B. Effect of molybdenum on nodulation, plant yield and nitrogen uptake in hairy vetch (Vicia villosaRoth). Soil Sci. Plant Nutr. 2015, 61, 664–675. [Google Scholar] [CrossRef]
- Brennan, R.F.; Penrose, B.; Bell, R.W. Micronutrients limiting pasture production in Australia. Crop. Pasture Sci. 2019, 70, 1053–1064. [Google Scholar] [CrossRef]
- Wichard, T.; Mishra, B.; Myneni, S.C.B.; Bellenger, J.-P.; Kraepiel, A.M.L. Storage and bioavailability of molybdenum in soils increased by organic matter complexation. Nat. Geosci. 2009, 2, 625–629. [Google Scholar] [CrossRef]
- Neal, A.L.; Bacq-Labreuil, A.; Zhang, X.; Clark, I.; Coleman, K.; Mooney, S.J.; Ritz, K.; Crawford, J.W. Soil as an extended composite phenotype of the microbial metagenome. Sci. Rep. 2020, 10, 10649. [Google Scholar] [CrossRef]
- Fagnano, M.; Adamo, P.; Zampella, M.; Fiorentino, N. Environmental and agronomic impact of fertilization with composted organic fraction from municipal solid waste: A case study in the region of Naples, Italy. Agric. Ecosyst. Environ. 2011, 141, 100–107. [Google Scholar] [CrossRef]
- Johnston, A.E.; Poulton, P.R.; Coleman, K. Soil Organic Matter: Its Importance in Sustainable Agriculture and Carbon Dioxide Fluxes. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: London, UK, 2009; Volume 101. [Google Scholar] [CrossRef]
- Reeves, D. The role of soil organic matter in maintaining soil quality in continuous cropping systems. Soil Tillage Res. 1997, 43, 131–167. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Pasture System | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Blue | Green | Red | |||||||||
| High | Low | p | High | Low | p | High | Low | p | p Systems | ||
| Soil Cu (mg kg−1) | Mean | 36 | 14 | *** | 30 | 23 | *** | 27 | 13 | *** | ***, R < B and G |
| Min | 25 | 10 | 21 | 14 | 15 | 9 | |||||
| Max | 44 | 28 | 36 | 30 | 36 | 16 | |||||
| Soil Fe (mg kg−1) | Mean | 44,421 | 31,941 | *** | 40,555 | 26,331 | *** | 45,219 | 24,243 | *** | **, B > G and R |
| Min | 35,378 | 20,800 | 35,644 | 19,974 | 30,570 | 18,559 | |||||
| Max | 50,103 | 46,958 | 48,527 | 31,329 | 50,658 | 31,614 | |||||
| Soil Mg (mg kg−1) | Mean | 1440 | 573 | *** | 908 | 617 | *** | 752 | 809 | / | ***, B > G and R |
| Min | 739 | 518 | 502 | 518 | 561 | 549 | |||||
| Max | 2040 | 620 | 1914 | 828 | 1577 | 1090 | |||||
| Soil Mn (mg kg−1) | Mean | 852 | 313 | *** | 802 | 342 | *** | 983 | 235 | *** | / |
| Min | 506 | 138 | 406 | 128 | 399 | 93 | |||||
| Max | 1092 | 793 | 1480 | 496 | 1677 | 315 | |||||
| Soil Mo (mg kg−1) | Mean | 3.4 | 1.5 | *** | 3.0 | 1.5 | *** | 2.4 | 1.3 | *** | ***, R < B and G |
| Min | 2.0 | 1.2 | 2.4 | 1.2 | 1.4 | 0.9 | |||||
| Max | 4.5 | 1.9 | 4.1 | 1.7 | 3.1 | 1.7 | |||||
| Soil Se (mg kg−1) | Mean | 1.2 | 0.9 | *** | 1.2 | 0.9 | *** | 1.3 | 1.0 | *** | *, R > G |
| Min | 1.0 | 0.7 | 1.0 | 0.7 | 1.1 | 0.9 | |||||
| Max | 1.4 | 1.0 | 1.5 | 1.7 | 1.4 | 1.2 | |||||
| Soil Zn (mg kg−1) | Mean | 85 | 61 | *** | 92 | 73 | *** | 91 | 62 | *** | *, G > B and R |
| Min | 72 | 51 | 75 | 65 | 81 | 51 | |||||
| Max | 111 | 81 | 121 | 81 | 103 | 73 | |||||
| Pasture System | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Blue | Green | Red | |||||||||
| High | Low | p | High | Low | p | High | Low | p | p Systems | ||
| Elevation (m ODN) | Mean | 168 | 180 | ** | 157 | 150 | / | 152 | 140 | *** | ***, B > G and R |
| Min | 160 | 170 | 142 | 143 | 130 | 129 | |||||
| Max | 175 | 185 | 180 | 156 | 180 | 172 | |||||
| Slope (°) | Mean | 5.7 | 4.1 | / | 6.3 | 5.4 | / | 4.1 | 3.3 | / | **, R < B and G |
| Min | 5.0 | 1.5 | 2.0 | 3.2 | 0.4 | 0.6 | |||||
| Max | 6.9 | 6.0 | 11.2 | 8.5 | 6.1 | 8.7 | |||||
| Aspect (°) | Mean | 103 | 67 | / | 213 | 240 | / | 253 | 264 | / | ***, B < G and R |
| Min | 17 | 1 | 4 | 41 | 93 | 48 | |||||
| Max | 157 | 281 | 339 | 318 | 323 | 342 | |||||
| Total C (%) | Mean | 4.5 | 4.0 | / | 6.1 | 5.9 | / | 3.6 | 3.9 | / | ***, G > B and R |
| Min | 3.0 | 2.7 | 5.2 | 4.7 | 2.6 | 2.0 | |||||
| Max | 6.8 | 5.0 | 7.0 | 7.5 | 4.7 | 5.4 | |||||
| Total N (%) | Mean | 0.5 | 0.4 | *** | 0.6 | 0.6 | / | 0.4 | 0.4 | / | ***, G > B and R |
| Min | 0.4 | 0.3 | 0.5 | 0.5 | 0.3 | 0.2 | |||||
| Max | 0.7 | 0.5 | 0.7 | 0.7 | 0.5 | 0.5 | |||||
| SOM (%) | Mean | 10.3 | 8.4 | ** | 14.0 | 12.5 | * | 8.3 | 8.9 | / | ***, G > B and R |
| Min | 8.1 | 5.8 | 12.2 | 10.5 | 6.4 | 4.2 | |||||
| Max | 13.7 | 10.7 | 16.0 | 16.5 | 11.2 | 12.6 | |||||
| pH | Mean | 5.8 | 5.4 | *** | 5.6 | 5.6 | / | 5.7 | 5.6 | / | / |
| Min | 5.4 | 5.1 | 5.2 | 5.3 | 5.5 | 5.4 | |||||
| Max | 6.9 | 5.6 | 5.9 | 5.8 | 6.0 | 5.9 | |||||
| Olsen P (mg kg−1) | Mean | 32.2 | 23.6 | / | 36.2 | 24.3 | / | 15.8 | 16.0 | / | *, R < B and G |
| Min | 10.3 | 15.0 | 16.5 | 19.4 | 7.1 | 9.8 | |||||
| Max | 113 | 35.2 | 138 | 31.0 | 30.2 | 23.1 | |||||
| WE Total P (mg kg−1) | Mean | 5.0 | 5.6 | / | 9.9 | 9.5 | / | 6.0 | 4.8 | / | ***, G > B and R |
| Min | 3.6 | 4.7 | 3.8 | 6.6 | 4.2 | 3.6 | |||||
| Max | 8.0 | 7.3 | 28.3 | 12.5 | 8.0 | 7.1 | |||||
| Bulk density (g cm3) | Mean | 1.03 | 0.96 | / | 0.90 | 0.86 | / | 1.04 | 0.97 | / | ***, G < B and R |
| Min | 0.68 | 0.88 | 0.78 | 0.78 | 0.72 | 0.58 | |||||
| Max | 1.29 | 1.19 | 1.06 | 0.99 | 1.19 | 1.35 | |||||
| Importance Variables/% Increase MSE | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| System | Elevation | Aspect | Slope | Soil Series | Total N | Total C | SOM | BD | pH | Var. Expl/CVR2 | |
| Soil total Cu | Blue | 12 | 15 | 8 | 5 | 0 | 1 | 1 | 2 | 10 | 0.52 |
| Green | 12 | −0 | 5 | 0 | −2 | 2 | 3 | −1 | 2 | 0.16 | |
| Red | 14 | −2 | 1 | 14 | −1 | 2 | −4 | −2 | −2 | 0.30 | |
| Soil total Fe | Blue | 5 | 8 | 3 | 2 | −3 | 2 | −3 | 11 | 5 | 0.24 |
| Green | −5 | 1 | −3 | −2 | −1 | 0 | −1 | 0 | −2 | <0.1 | |
| Red | 10 | −3 | 8 | 9 | −3 | −3 | −3 | −3 | −7 | <0.1 | |
| Soil total Mg | Blue | 7 | 8 | 5 | 15 | 1 | −2 | 3 | 0 | 6 | 0.54 |
| Green | −2 | 7 | −1 | −1 | 1 | −0 | 5 | −3 | 3 | <0.1 | |
| Red | 17 | −1 | 16 | 8 | 4 | 6 | 8 | 1 | −1 | 0.59 | |
| Soil total Mn | Blue | 10 | −2 | 14 | 0 | −2 | −5 | −6 | 10 | 0 | 0.24 |
| Green | −2 | 6 | −1 | 2 | −3 | −2 | −1 | 6 | −0 | <0.1 | |
| Red | −1 | 4 | 12 | 1 | −3 | −3 | 1 | −1 | −1 | <0.1 | |
| Soil total Mo | Blue | 10 | 6 | 7 | 4 | 2 | −5 | −1 | 7 | −3 | <0.1 |
| Green | 1 | −2 | −6 | −1 | −2 | −3 | 1 | −2 | −1 | <0.1 | |
| Red | 15 | −1 | 6 | 12 | −1 | 1 | −3 | −4 | −5 | 0.22 | |
| Soil total Se | Blue | 10 | 11 | 11 | 3 | 5 | −5 | −1 | 4 | 7 | 0.38 |
| Green | 4 | 9 | −3 | 1 | −1 | −0 | 8 | 4 | 5 | 0.22 | |
| Red | 6 | 7 | 7 | −1 | −6 | −5 | −1 | −5 | −8 | <0.1 | |
| Soil total Zn | Blue | 5 | −3 | 0 | −1 | 7 | −4 | 3 | −1 | 1 | <0.1 |
| Green | −3 | 4 | 0 | −3 | −1 | −2 | 2 | 0 | −5 | <0.1 | |
| Red | 5 | 1 | 6 | 9 | −4 | −4 | −3 | −3 | −4 | <0.1 | |
| Pasture System | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Blue | Green | Red | |||||||||
| High | Low | p | High | Low | p | High | Low | p | p Systems | ||
| Herbage Cu (mg kg−1) | Mean | 9.9 | 5.8 | ** | 9.1 | 7.9 | / | 13.8 | 9.5 | *** | ***, R > B and G |
| Min | 5.7 | 3.3 | 5.6 | 3.7 | 5.4 | 5.1 | |||||
| Max | 16.1 | 8.0 | 14.3 | 15.0 | 32.9 | 20.7 | |||||
| Herbage Fe (mg kg−1) | Mean | 369 | 517 | / | 394 | 433 | / | 623 | 359 | / | / |
| Min | 122 | 230 | 87 | 178 | 115 | 147 | |||||
| Max | 1272 | 1162 | 1088 | 949 | 2281 | 1092 | |||||
| Herbage Mg (mg kg−1) | Mean | 1509 | 1323 | / | 1545 | 1537 | / | 1288 | 1336 | / | *, ***, G > B and R |
| Min | 1052 | 887 | 1177 | 1004 | 1064 | 857 | |||||
| Max | 1897 | 1744 | 1986 | 2186 | 1664 | 1832 | |||||
| Herbage Mn (mg kg−1) | Mean | 93 | 127 | / | 184 | 137 | * | 130 | 117 | / | ***, G > B and R |
| Min | 45 | 71 | 90 | 65 | 61 | 45 | |||||
| Max | 242 | 259 | 366 | 198 | 223 | 271 | |||||
| Herbage Mo (mg kg−1) | Mean | 2.6 | 3.9 | ** | 2.5 | 2.9 | / | 2.5 | 2.0 | / | ***, B > R |
| Min | 1.1 | 1.9 | 1.2 | 1.2 | 1.1 | 0.9 | |||||
| Max | 7.0 | 7.6 | 6.5 | 5.1 | 4.4 | 3.8 | |||||
| Herbage Se (mg kg−1) | Mean | 0.18 | 0.18 | / | 0.21 | 0.16 | ** | 0.18 | 0.12 | *** | *, **, R < B and G |
| Min | 0.08 | 0.12 | 0.10 | 0.10 | 0.11 | 0.05 | |||||
| Max | 0.39 | 0.24 | 0.34 | 0.24 | 0.29 | 0.22 | |||||
| Herbage Zn (mg kg−1) | Mean | 29 | 23 | / | 30 | 23 | * | 25 | 21 | / | / |
| Min | 20 | 13 | 16 | 18 | 16 | 15 | |||||
| Max | 49 | 78 | 66 | 28 | 43 | 47 | |||||
| System | Top 5 Importance Variables/% Increase MSE | Var. Expl/CVR2 | |||||
|---|---|---|---|---|---|---|---|
| Herbage Cu uptake | Blue | Soil Mo/11 | Aspect/8 | Soil Mg/8 | Elevation/7 | Soil K/7 | 0.57 |
| Green | SOM/11 | Soil Se/11 | Soil Mg/7 | Soil Mo/4 | Aspect/3 | 0.43 | |
| Red | Soil K/8 | WE TP/7 | Soil Cr/6 | Olsen P/5 | Soil Se/5 | 0.18 | |
| Herbage Fe uptake | Blue | Soil Mo/10 | Soil K/10 | Soil Mg/7 | Soil Se/7 | Elevation/6 | 0.43 |
| Green | SOM/8 | Soil Ni/7 | Soil Se/4 | Total C/3 | Soil Fe/3 | / | |
| Red | Aspect/7 | WE TP/7 | Soil Cu/7 | Elevation/6 | Soil Zn/6 | 0.32 | |
| Herbage Mg uptake | Blue | Soil Mo/11 | Aspect/9 | Elevation/7 | Soil K/7 | Soil Na/7 | 0.44 |
| Green | SOM/10 | Soil Ni/10 | Soil Se/9 | Soil Mg/4 | Aspect/2 | 0.26 | |
| Red | Soil K/13 | SOM/9 | Soil Na/9 | Soil Ti/7 | Soil Al/6 | 0.46 | |
| Herbage Mn uptake | Blue | Soil Mo/12 | Elevation/8 | Soil K/8 | Aspect/7 | Soil Na/7 | 0.44 |
| Green | SOM/10 | Soil Se/10 | Soil Ni/9 | Soil Cu/3 | Soil Fe/3 | 0.31 | |
| Red | Soil Se/8 | Soil Na/7 | Elevation/6 | Aspect/6 | Soil Al/6 | 0.28 | |
| Herbage Mo uptake | Blue | Soil Cr/4 | Aspect/3 | Soil K/3 | Soil Se/3 | Soil Cu/2 | / |
| Green | pH/8 | Elevation/7 | Olsen P/3 | Soil Ni/3 | Soil P/3 | 0.14 | |
| Red | Soil Mn/5 | WE TP/3 | Soil Cr/3 | Elevation/2 | Soil Mg/2 | / | |
| Herbage Se uptake | Blue | Soil Mo/11 | Aspect/8 | Soil Na/8 | Elevation/7 | Soil K/7 | 0.47 |
| Green | Soil Ni/10 | Soil Se/10 | SOM/8 | Soil Fe/5 | Soil Mo/5 | 0.23 | |
| Red | Soil K/8 | Soil Se/7 | Soil Al/6 | Soil Co/6 | Soil Mn/6 | 0.40 | |
| Herbage Zn uptake | Blue | Soil Mo/12 | Elevation/8 | Aspect/8 | Soil Se/8 | Soil Mg/7 | 0.59 |
| Green | Soil Se/10 | SOM/9 | Soil Ni/8 | pH/5 | Soil Fe/5 | 0.28 | |
| Red | Soil K/14 | Soil Se/8 | SOM/5 | Soil Al/5 | Soil Na/5 | 0.36 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, C.L.; Darch, T.; Harris, P.; Beaumont, D.A.; Haefele, S.M. The Distribution of Soil Micro-Nutrients and the Effects on Herbage Micro-Nutrient Uptake and Yield in Three Different Pasture Systems. Agronomy 2021, 11, 1731. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091731
Thomas CL, Darch T, Harris P, Beaumont DA, Haefele SM. The Distribution of Soil Micro-Nutrients and the Effects on Herbage Micro-Nutrient Uptake and Yield in Three Different Pasture Systems. Agronomy. 2021; 11(9):1731. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091731
Chicago/Turabian StyleThomas, Cathy L., Tegan Darch, Paul Harris, Deborah A. Beaumont, and Stephan M. Haefele. 2021. "The Distribution of Soil Micro-Nutrients and the Effects on Herbage Micro-Nutrient Uptake and Yield in Three Different Pasture Systems" Agronomy 11, no. 9: 1731. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091731