
New Pathotype Nomenclature for Better Characterisation the Virulence and Diversity of Blumeria graminis f.sp. avenae Populations


Abstract
:1. Introduction
2. Materials and Methods
2.1. Location of Pathogen Populations and Dates of Sampling
2.2. Multiplication of Inoculum
2.3. Differential Sets and Inoculation of the Leaf Segments
2.4. Virulence Determination, Pathotype Designation, and Distribution
2.5. Data Analysis
3. Results
3.1. Virulence Frequency
3.2. New Nomenclature for B. graminis f.sp. avenae Phenotypes
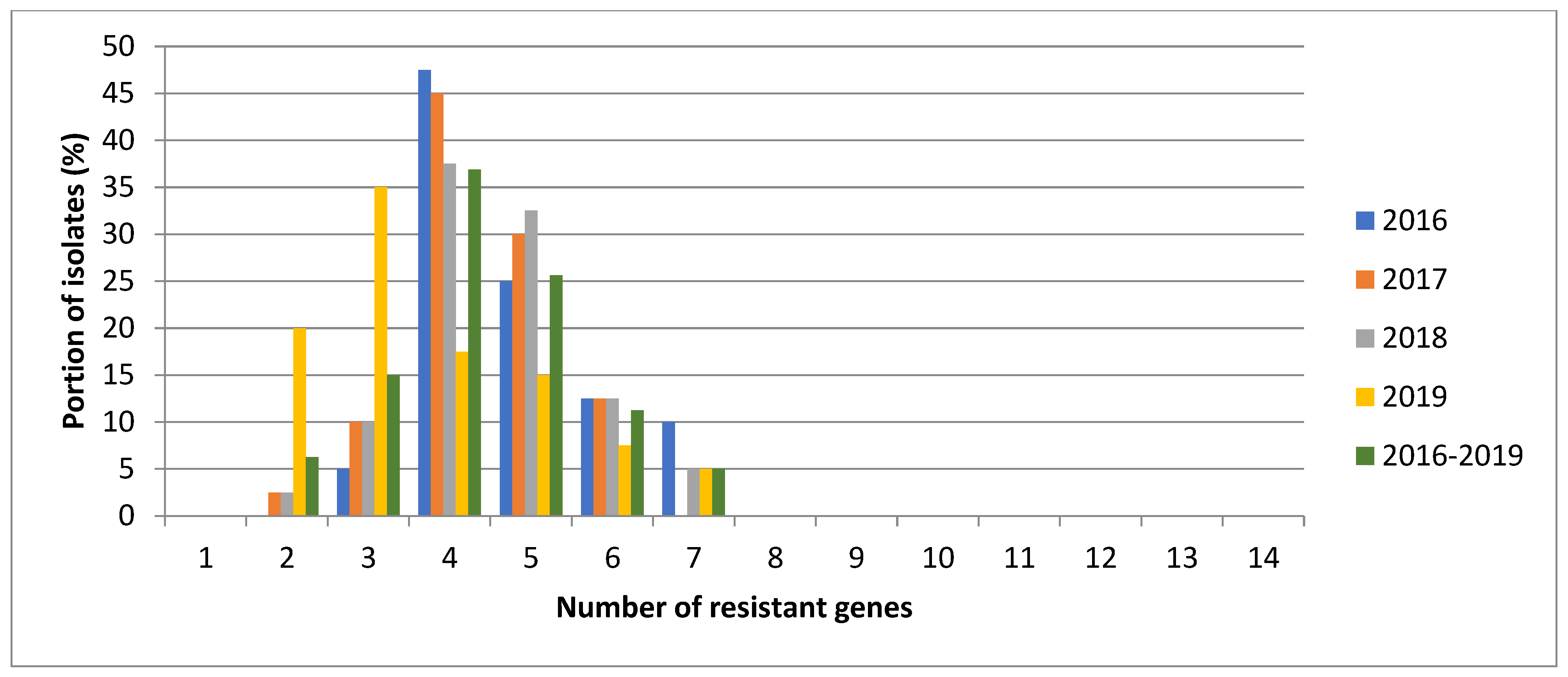
3.3. Pathotypes Structure
3.4. Diversity within and Distance between Populations
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nazir, N.; Bilal, S.; Bhat, K.; Shah, T.; Badri, Z.; Bhat, F.; Wani, T.; Mugal, M.; Parveen, S.; Dorjry, S. Effect of Climate Change on Plant Diseases. Artic. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 250–256. [Google Scholar] [CrossRef]
- Grulke, N.E. The nexus of host and pathogen phenology: Understanding the disease triangle with climate change. New Phytol. 2011, 189, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Braun, U.; Cook, R.T.A.; Inman, A.J.; Shin, H.D. The taxonomy of the powdery mildew fungi. In The Powdery Mildews: A Comprehensive Treatis; APS Press: New York, NY, USA, 2002; pp. 13–55. [Google Scholar]
- Baker, R.H.A.; Sansford, C.E.; Jarvis, C.H.; Cannon, R.J.C.; MacLeod, A.; Walters, K.F.A. The role of climatic mapping in predicting the potential geographical distribution of non-indigenous pests under current and future climates. Agric. Ecosyst. Environ. 2000, 82, 57–71. [Google Scholar] [CrossRef]
- Tang, X.; Luo, Y.; Ma, Z.; Fan, J.; Zhou, Y.; Cao, X.; Xu, X.; Jiang, Y.; Luo, Y.; Ma, Z.; et al. Effects of Climate Change on Epidemics of Powdery Mildew in Winter Wheat in China. Plant Dis. 2017, 101, 1753–1760. [Google Scholar] [CrossRef] [Green Version]
- Dean, R.; Van Kan, J.A.N.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.I.M.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Schwarzbach, E.; Smith, I.M. Erysiphe graminis DC. In European Handbook of Plant Diseases; Smith, I.M., Dunez, J., Lelliot, R.A., Philips, D.H., Archer, S.A., Eds.; Blackwell: Kay County, OK, USA, 1988; pp. 259–261. [Google Scholar]
- Roderick, H.W.; Jones, E.R.L.; Šebesta, J.; Sebesta, J. Resistance to oat powdery mildew in Britain and Europe: A review. Ann. Appl. Biol. 2000, 136, 85–91. [Google Scholar] [CrossRef]
- Sebesta, J.; Kummer, M.; Roderick, H.W.W.; Hoppe, H.D.D.; Swierczewski, A.; Mueller, K.; Cervenka, J.; Swierczewski, A.; Muller, K. Breeding oats for resistance to rusts and powdery mildew in central Europe. Ochr. Rostl. 1991, 27, 229–238. [Google Scholar]
- Xue, L.H.; Li, C.J.; Zhao, G.Q. First Report of Powdery Mildew Caused by Blumeria graminis on Avena sativa in China. Plant Dis. 2017, 101, 1954. [Google Scholar] [CrossRef]
- Banyal, D.K.; Sood, V.K.; Singh, A.; Mawar, R. Integrated management of oat diseases in north-western Himalaya. Range Manag. Agrofor. 2016, 37, 84–87. [Google Scholar]
- Traskovetskaya, V.; Gorash, A.; Liatukas, Ž.; Saulyak, N.; Ternovyi, K.; Babayants, O.; Ruzgas, V.; Leistrumaitė, A. Virulence and diversity of the blumeria graminis f. Sp. tritici populations in lithuania and Southern Ukraine. Zemdirbyste 2019, 106, 107–116. [Google Scholar] [CrossRef]
- Chong, J.; Leonard, K.J.; Salmeron, J.J. A North American System of Nomenclature for Puccinia coronata f. sp. avenae. Plant Dis. 2000, 84, 580–585. [Google Scholar] [CrossRef] [Green Version]
- Roelfs, A.P.; Martens, J.W. An International System of Nomenclature for Puccinia graminis f. sp. Tritici. Phytopathology 1988, 78, 526–533. [Google Scholar] [CrossRef] [Green Version]
- Long, D.L.; Kolmer, J.A. A north American system of nomenclature for Puccinia recindita f.sp. tritici. Phytopathology 1989, 79, 525–529. [Google Scholar] [CrossRef]
- Hsam, S.L.K.; Pederina, E.; Gorde, S.; Zeller, F.J. Genetic studies of powdery mildew resistance in common oat (Avena staiva L.). II. Cultivars and breeding lines grown in Northern and Eastern Europe. Hereditas 1998, 129, 227–230. [Google Scholar] [CrossRef]
- Okoń, S.; Paczos-Grzeda, E.; Ociepa, T.; Koroluk, A.; Sowa, S.; Kowalczyk, K.; Chrzą, M. Avena sterilis L. Genotypes as a Potential Source of Resistance to Oat Powdery Mildew. Plant Dis. 2016, 100, 2145–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okoń, S.; Kowalczyk, K. Screening oat landraces for resistance to Blumeria graminis f. sp. avenae. J. Plant Pathol. 2020, 102, 893–898. [Google Scholar] [CrossRef] [Green Version]
- Hsam, S.L.K.; Peters, N.; Paderina, E.V.; Felsenstein, F.; Oppitz, K.; Zeller, F.J. Genetic studies of powdery mildew resistance in common oat (Avena sativa L.) I. Cultivars and breeding lines grown in Western Europe and North America. Euphytica 1997, 96, 421–427. [Google Scholar] [CrossRef]
- Herrmann, M.H.; Mohler, V. Locating two novel genes for resistance to powdery mildew from Avena byzantine in the oat genome. Plant Breed. 2018, 137, 832–838. [Google Scholar] [CrossRef]
- Okoń, S.M.; Ociepa, T. Effectiveness of new sources of resistance against oat powdery mildew identified in A. sterilis. J. Plant Dis. Prot. 2018, 125, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mains, E.B. Inheritance of resistance to powdery mildew, Erysiphe graminis tritici, in wheat. Phytopathology 1934, 24, 1257–1261. [Google Scholar]
- Dreiseitl, A.; Kosman, E. Virulence phenotypes of Blumeria graminis f. sp. hordei in South Africa. Eur. J. Plant Pathol. 2015, 136, 113–121. [Google Scholar] [CrossRef]
- Kosman, E. Difference and diversity of plant pathogen populations: A new approach for measuring. Phytopathology 1996, 86, 1152–1155. [Google Scholar]
- Kosman, E.; Leonard, K.J. Conceptual analysis of methods applied to assessment of diversity within and distance between populations with asexual or mixed mode of reproduction. New Phytol. 2007, 174, 683–696. [Google Scholar] [CrossRef]
- Herrmann, A.; Löwer, C.; Schachtel, G. A new tool for entry and analysis of virulence data for plant pathogens. Plant Pathol. 1999, 48, 154–158. [Google Scholar] [CrossRef]
- Schachtel, G.A.; Dinoor, A.; Herrmann, A.; Kosman, E. Comprehensive Evaluation of Virulence and Resistance Data: A New Analysis Tool. Plant Dis. 2012, 96, 1060–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okoń, S.M. Identification of powdery mildew resistance genes in Polish common oat (Avena sativa L.) cultivars using host-pathogen tests. Acta Agrobot. 2012, 65, 63–68. [Google Scholar] [CrossRef]
- Okoń, S.; Ociepa, T.; Paczos-Grzęda, E.; Kowalczyk, K. Analysis of the level of resistance of Polish oat cultivars (Avena sativa L.) to powdery mildew (Blumeria graminis DC. f. sp. avenae Em. Marchal.). Agron. Sci. 2016, 61, 51–60. [Google Scholar]
- Kowalczyk, K.; Hsam, S.L.K.; Zeller, F.J. Identification of oat powdery mildew resistance 2 (OMR2) and Polish common oat (Avena sativa L.) cultivars. In Workshop “Resistance of Cereals to Biotic Stresses”; IHAR: Radzików, Poland, 2004; pp. 122–125. [Google Scholar]
- Okoń, S.M. Effectiveness of resistance genes to powdery mildew in oat. Crop Prot. 2015, 74, 48–50. [Google Scholar] [CrossRef]
- Hsam, S.L.K.; Mohler, V.; Zeller, F.J. The genetics of resistance to powdery mildew in cultivated oats (Avena sativa L.): Current status of major genes. J. Appl. Genet. 2014, 55, 155–162. [Google Scholar] [CrossRef]
- Cieplak, M.; Terlecka, K.; Ociepa, T.; Zimowska, B.; Okoń, S. Virulence Structure of Blumeria graminis f. sp. avenae Populations in Poland across 2014–2015. Plant Pathol. J. 2021, 37, 115–123. [Google Scholar] [CrossRef]
- Ociepa, T.; Okoń, S.; Nucia, A.; Leśniowska-Nowak, J.; Paczos-Grzęda, E.; Bisaga, M. Molecular identification and chromosomal localization of new powdery mildew resistance gene Pm11 in oat. Theor. Appl. Genet. 2020, 133, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Statistics Poland/Databases. Available online: https://stat.gov.pl/en/databases/ (accessed on 10 September 2021).
- Paczos-Grzęda, E.; Sowa, S. Virulence Structure and Diversity of Puccinia coronata f. sp. avenae P. Syd. & Syd. in Poland During 2013 to 2015. Plant Dis. 2019, 103, 1559–1564. [Google Scholar]
- Babayants, O.V.; Babayants, L.T.; Traskovetskaya, V.A.; Gorash, A.F.; Saulyak, N.I.; Galaev, A.V. Race Composition of Blumeria graminis (DC) Speer f. sp. Tritici in the South of Ukraine and Effectiveness of Pm-genes in 2004–2013. Cereal Res. Commun. 2015, 43, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Okoń, S.M.; Ociepa, T. Virulence structure of the Blumeria graminis DC. f. sp. avenae populations occurring in Poland across 2010–2013. Eur. J. Plant Pathol. 2017, 149, 711–718. [Google Scholar] [CrossRef]
- Etterson, J.R.; Shaw, R.G. Constraint to adaptive evolution in response to global warming. Science 2001, 294, 151–154. [Google Scholar] [CrossRef] [Green Version]
- Dumbadze, R.Z.; Sikharulidze, Z.V. Virulence Structure of the Wheat Stem Rust Population in Georgia. Int. J. Agric. Innov. Res. 2016, 4, 5. [Google Scholar]
- Fetch, T.; Zegeye, T.; Park, R.F.; Hodson, D.; Wanyera, R. Detection of wheat stem rust races TTHSK and PTKTK in the Ug99 race group in Kenya in 2014. Plant Dis. 2016, 100, 1495. [Google Scholar] [CrossRef]
- Skolotneva, E.S.; Kosman, E.; Patpour, M.; Kelbin, V.N.; Morgounov, A.I.; Shamanin, V.P.; Salina, E.A. Virulence Phenotypes of Siberian Wheat Stem Rust Population in 2017–2018. Front. Agron. 2020, 2, 6. [Google Scholar] [CrossRef]
- Rsaliyev, A.; Yskakova, G.; Maulenbay, A.; Zakarya, K.; Rsaliyev, S. Virulence and race structure of Puccinia graminis f. sp. tritici in Kazakhstan. Plant Prot. Sci. 2020, 56, 275–284. [Google Scholar] [CrossRef]
- Nemati, Z.; Mostowfizadeh-Ghalamfarsa, R.; Dadkhodaie, A.; Mehrabi, R.; Steffenson, B.J. Virulence of leaf rust physiological races in Iran from 2010 to 2017. Plant Dis. 2020, 104, 363–372. [Google Scholar] [CrossRef]
- Boshoff, W.H.P.; Labuschagne, R.; Terefe, T.; Pretorius, Z.A.; Visser, B. New Puccinia triticina races on wheat in South Africa. Australas. Plant Pathol. 2018, 47, 325–334. [Google Scholar] [CrossRef]
- Mantovani, P.; MacCaferri, M.; Tuberosa, R.; Kolmer, J. Virulence phenotypes and molecular genotypes in collections of puccinia triticina from Italy. Plant Dis. 2010, 94, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Leonard, K.J.; Paul, S.; Martinelli, J.A. Virulence of Oat Crown Rust in Brazil and Uruguay. Plant Dis. 2005, 89, 802–808. [Google Scholar] [CrossRef]
- Carson, M.L. Additional Sources of Broad-Spectrum Resistance to Puccinia coronata f. sp. avenae from Canadian Accessions of Avena barbata. Plant Dis. 2010, 94, 1405–1410. [Google Scholar] [CrossRef] [Green Version]
- Paczos-Grzęda, E.; Sowa, S.; Boczkowska, M.; Langdon, T. Detached Leaf Assays for Resistance to Crown Rust Reveal Diversity Within Populations of Avena sterilis. Plant Dis. 2019, 103, 832–840. [Google Scholar] [CrossRef]
- Paczos-Grzeda, E.; Sowa, S.; Koroluk, A.; Langdon, T. Characteristics of Resistance to Puccinia coronata f. sp. avenae in Avena fatua. Plant Dis. 2018, 102, 2616–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsam, S.L.K.; Zeller, F.J. Chromosomal location of genes for resistance to powdery mildew in cultivated oat (Avena sativa L.). 1. Gene Eg-3 in the cultivar ‘Mostyn’. Plant Breed. 1998, 178, 177–178. [Google Scholar] [CrossRef]
- Herrmann, M.; Roderick, H.W. Characterisation of new oat germplasm for resistance to powdery mildew. Euphytica 1996, 89, 405–410. [Google Scholar] [CrossRef]
- Mohler, V.; Zeller, F.J.; Hsam, S.L.K. Molecular mapping of powdery mildew resistance gene Eg-3 in cultivated oat (Avena sativa L. cv. Rollo). J. Appl. Genet. 2012, 53, 145–148. [Google Scholar] [CrossRef]
- Sánchez-Martín, J.; Rubiales, D.; Prats, E. Resistance to powdery mildew (Blumeria graminis f. sp. avenae) in oat seedlings and adult plants. Plant Pathol. 2011, 60, 846–856. [Google Scholar] [CrossRef] [Green Version]
- Okoń, S.; Ociepa, T.; Paczos-Grzęda, E.; Ladizinsky, G. Evaluation of resistance to Blumeria graminis DC f. sp. avenae, in Avena murphyi and A. magna genotypes. Crop Prot. 2018, 106, 177–181. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cultivar/Line | Gene Symbol | Host Set | Pedigree |
|---|---|---|---|
| Jumbo | Pm1 | 1 | Flämingsstern/AJ20–61//Faggot |
| CC3678 | Pm2 | 2 | A. hirtula |
| Mostyn | Pm3 | 1 | 05443/Condor |
| Av1860 | Pm4 | 2 | A. sativa/A. barbata |
| Am27 | Pm5 | 2 | A. sativa/A. macrostachya |
| Bruno | Pm6 | 1 | Halla/Gambo |
| APR122 | Pm7 | 2 | A. sativa/A. eriantha |
| Canyon | Pm7 | 3 | A. sativa/A. barbata |
| Rollo | Pm3 + Pm8 | 1 | LP75-512/W17286 |
| AVE2406 | Pm9 | 3 | A. byzantina |
| AVE2925 | Pm10 | 3 | A. byzantina |
| CN113536 | Pm11 | 3 | A. sativa/A. sterilis |
| CN67383 | UA.ster. | 4 | A. sterilis |
| Pl 51586 | UA.stri. | 4 | A. strigosa |
| Fuchs | - | - | - |
| Letter Code. | Host Set 1 | Pm1 | Pm3 | Pm6 | Pm8 |
|---|---|---|---|---|---|
| Host Set 2 | Pm2 | Pm4 | Pm5 | Pm7a | |
| Host Set 3 | Pm7b | Pm9 | Pm10 | Pm11 | |
| Host Set 4 | A.ster | A.st | - | - | |
| B | L | L | L | L | |
| C | L | L | L | H | |
| D | L | L | H | L | |
| F | L | L | H | H | |
| G | L | H | L | L | |
| H | L | H | L | H | |
| J | L | H | H | L | |
| K | L | H | H | H | |
| L | H | L | L | L | |
| M | H | L | L | H | |
| N | H | L | H | L | |
| P | H | L | H | H | |
| Q | H | H | L | L | |
| R | H | H | L | H | |
| S | H | H | H | L | |
| T | H | H | H | H |
| Cultivar | Gene | Frequency (%) | ||||
|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | 2019 | 2016–2019 | ||
| Jumbo | Pm1 | 100 | 77.5 | 97.5 | 95 | 92.5 |
| CC3678 | Pm2 | 0 | 0 | 0 | 20 | 5 |
| Mostyn | Pm3 | 90 | 97.5 | 92.5 | 62.5 | 85.6 |
| Av1860 | Pm4 | 0 | 0 | 0 | 0 | 0 |
| Am27 | Pm5 | 0 | 0 | 0 | 0 | 0 |
| Bruno | Pm6 | 100 | 100 | 87.5 | 62.5 | 87.5 |
| APR122 | Pm7 | 0 | 0 | 0 | 0 | 0 |
| Canyon | Pm7 | 12.5 | 0 | 0 | 0 | 3.1 |
| Rollo | Pm3 + Pm8 | 90 | 100 | 92.5 | 60 | 85.6 |
| AVE2406 | Pm9 | 7.5 | 10 | 30 | 7.5 | 13.8 |
| AVE2925 | Pm10 | 22.5 | 17.5 | 15 | 12.5 | 16.9 |
| CN113536 | Pm11 | 27.5 | 27.5 | 17.5 | 15 | 21.9 |
| CN67383 | UA.ster. | 20 | 10 | 22.5 | 17.5 | 17.5 |
| Pl 51586 | UA.stri. | 5 | 0 | 2.5 | 17.5 | 6.3 |
| Pm1 | Pm3 | Pm6 | Pm3 + 8 | Pm2 | Pm4 | Pm5 | Pm7 | Pm7 | Pm9 | Pm10 | Pm11 | U A. Sterilis | U A. Strigosa | Number of Isolates | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | 2019 | 2016–2019 | |||||||||||||||
| TBBB | + | + | + | + | 18 | 14 | 13 | 3 | 48 | ||||||||||
| TBCB | + | + | + | + | + | 5 | 5 | 3 | 13 | ||||||||||
| RBBB | + | + | + | 2 | 8 | 10 | |||||||||||||
| TBBL | + | + | + | + | + | 2 | 4 | 1 | 7 | ||||||||||
| TBGB | + | + | + | + | + | 1 | 4 | 1 | 6 | ||||||||||
| NBCB | + | + | 5 | 5 | |||||||||||||||
| TBDB | + | + | + | + | + | 4 | 1 | 5 | |||||||||||
| TBJB | + | + | + | + | + | + | 1 | 3 | 1 | 5 | |||||||||
| KBBB | + | + | + | 4 | 4 | ||||||||||||||
| TBCL | + | + | + | + | + | + | 3 | 1 | 4 | ||||||||||
| TBLB | + | + | + | + | + | 4 | 4 | ||||||||||||
| KBCB | + | + | + | + | 3 | 3 | |||||||||||||
| TBHB | + | + | + | + | + | + | 2 | 1 | 3 | ||||||||||
| LBDB | + | + | 2 | 2 | |||||||||||||||
| NBBB | + | + | 2 | 2 | |||||||||||||||
| NBBL | + | + | + | 2 | 2 | ||||||||||||||
| NBCG | + | + | + | + | 2 | 2 | |||||||||||||
| NBDB | + | + | + | 2 | 2 | ||||||||||||||
| QBBG | + | + | + | 2 | 2 | ||||||||||||||
| TBDL | + | + | + | + | + | + | 1 | 1 | 2 | ||||||||||
| TBKB | + | + | + | + | + | + | + | 2 | 2 | ||||||||||
| TLBB | + | + | + | + | + | 2 | 2 | ||||||||||||
| TLBL | + | + | + | + | + | + | 2 | 2 | |||||||||||
| DBBL | + | + | 1 | 1 | |||||||||||||||
| FBBB | + | + | 1 | 1 | |||||||||||||||
| FLBB | + | + | + | 1 | 1 | ||||||||||||||
| HBBB | + | + | 1 | 1 | |||||||||||||||
| KBBL | + | + | + | + | 1 | 1 | |||||||||||||
| LBCL | + | + | + | 1 | 1 | ||||||||||||||
| NBCL | + | + | + | + | 1 | 1 | |||||||||||||
| NBDL | + | + | + | + | 1 | 1 | |||||||||||||
| NBJL | + | + | + | + | + | 1 | 1 | ||||||||||||
| NBNL | + | + | + | + | + | 1 | 1 | ||||||||||||
| RBBG | + | + | + | + | 1 | 1 | |||||||||||||
| RBCB | + | + | + | + | 1 | 1 | |||||||||||||
| RBGB | + | + | + | + | 1 | 1 | |||||||||||||
| RLBG | + | + | + | + | + | 1 | 1 | ||||||||||||
| TBBG | + | + | + | + | + | 1 | 1 | ||||||||||||
| TBCG | + | + | + | + | + | + | 1 | 1 | |||||||||||
| TBDQ | + | + | + | + | + | + | + | 1 | 1 | ||||||||||
| TBFB | + | + | + | + | + | + | 1 | 1 | |||||||||||
| TBHL | + | + | + | + | + | + | + | 1 | 1 | ||||||||||
| TBJG | + | + | + | + | + | + | + | 1 | 1 | ||||||||||
| TBJL | + | + | + | + | + | + | + | 1 | 1 | ||||||||||
| TLDL | + | + | + | + | + | + | + | 1 | 1 | ||||||||||
| TLJB | + | + | + | + | + | + | + | 1 | 1 | ||||||||||
| Parameter | 2016 | 2017 | 2018 | 2019 |
|---|---|---|---|---|
| No. of isolates | 40 | 40 | 40 | 40 |
| No. of different pathotypes | 12 | 13 | 16 | 21 |
| Gene diversity (Nei index Hs) equivalent to ADWm diversity | 0.134 | 0.103 | 0.136 | 0.216 |
| Simpson index Si | 0.758 | 0.828 | 0.853 | 0.916 |
| Shannon normalized index Sh | 0.514 | 0.579 | 0.635 | 0.754 |
| Kosman index KWm | 0.164 | 0.129 | 0.168 | 0.300 |
| Location | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Choryń | Strzelce | Czesławice | Polanowice | Poland (Total) | ||||||||||||||||
| Year | 2016 | 2017 | 2018 | 2019 | 2016 | 2017 | 2018 | 2019 | 2016 | 2017 | 2018 | 2019 | 2016 | 2017 | 2018 | 2019 | 2016 | 2017 | 2018 | 2019 |
| Rainfall [mm] | 608 | 668 | 373 | 393 | 751 | 832 | 520 | 388 | 698 | 612 | 479 | 531 | 745 | 702 | 569 | 639 | 666 | 733 | 489 | 556 |
| Temperature [°C] | 9.8 | 9.7 | 10.7 | 11.1 | 9.3 | 8.8 | 9.8 | 10.3 | 8.7 | 8.4 | 9.3 | 9.8 | 9.4 | 9.1 | 10 | 10.4 | 9.2 | 9 | 9.8 | 10.2 |
| Wind velocity [m/s] | 3.6 | 4 | 3.9 | 3.9 | 3.3 | 3.4 | 3.1 | 3.3 | 2.9 | 3 | 2.7 | 3 | 3.1 | 2.6 | 3.1 | 3.1 | 3.23 | 3.25 | 3.2 | 3.33 |
| Insolation [h] | 1823 | 1739 | 2225 | 2040 | 1840 | 1684 | 2170 | 2065 | 1872 | 1783 | 2143 | 2210 | 1750 | 1710 | 1949 | 1977 | 1821 | 1729 | 2122 | 2073 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okoń, S.; Cieplak, M.; Kuzdraliński, A.; Ociepa, T. New Pathotype Nomenclature for Better Characterisation the Virulence and Diversity of Blumeria graminis f.sp. avenae Populations. Agronomy 2021, 11, 1852. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091852
Okoń S, Cieplak M, Kuzdraliński A, Ociepa T. New Pathotype Nomenclature for Better Characterisation the Virulence and Diversity of Blumeria graminis f.sp. avenae Populations. Agronomy. 2021; 11(9):1852. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091852
Chicago/Turabian StyleOkoń, Sylwia, Magdalena Cieplak, Adam Kuzdraliński, and Tomasz Ociepa. 2021. "New Pathotype Nomenclature for Better Characterisation the Virulence and Diversity of Blumeria graminis f.sp. avenae Populations" Agronomy 11, no. 9: 1852. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy11091852