
Flavonoids and Limonoids Profiles Variation in Leaves from Mandarin Cultivars and Its Relationship with Alternate Bearing


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.1.1. Reagents and Equipment
2.1.2. Standard Solutions Preparation
2.1.3. Sample Preparation
2.1.4. Chromatographic and MS/MS Conditions
2.1.5. Data Treatment
3. Results and Discussion
3.1. Optimization of MS/MS Parameters of the Target Compounds
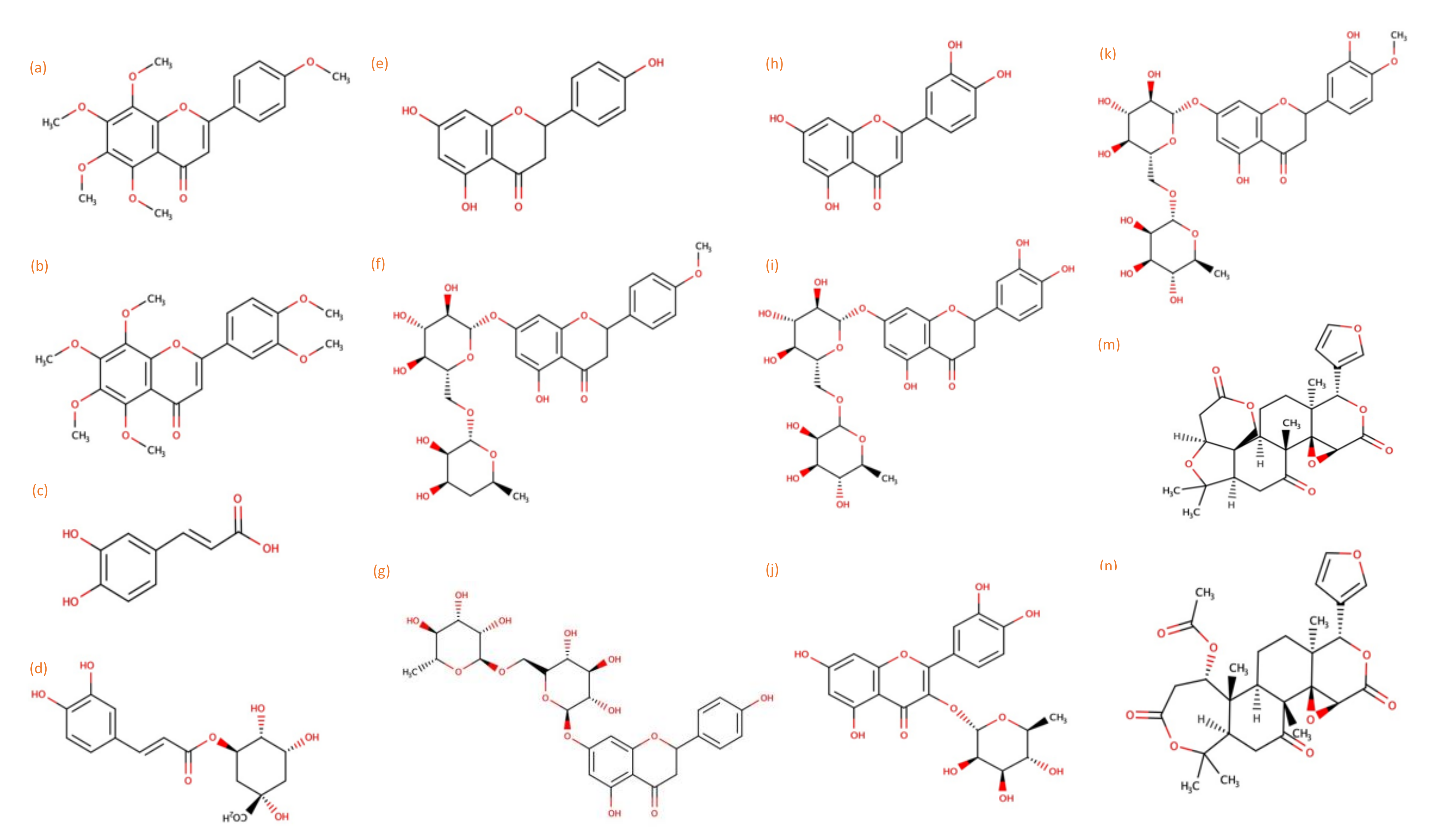
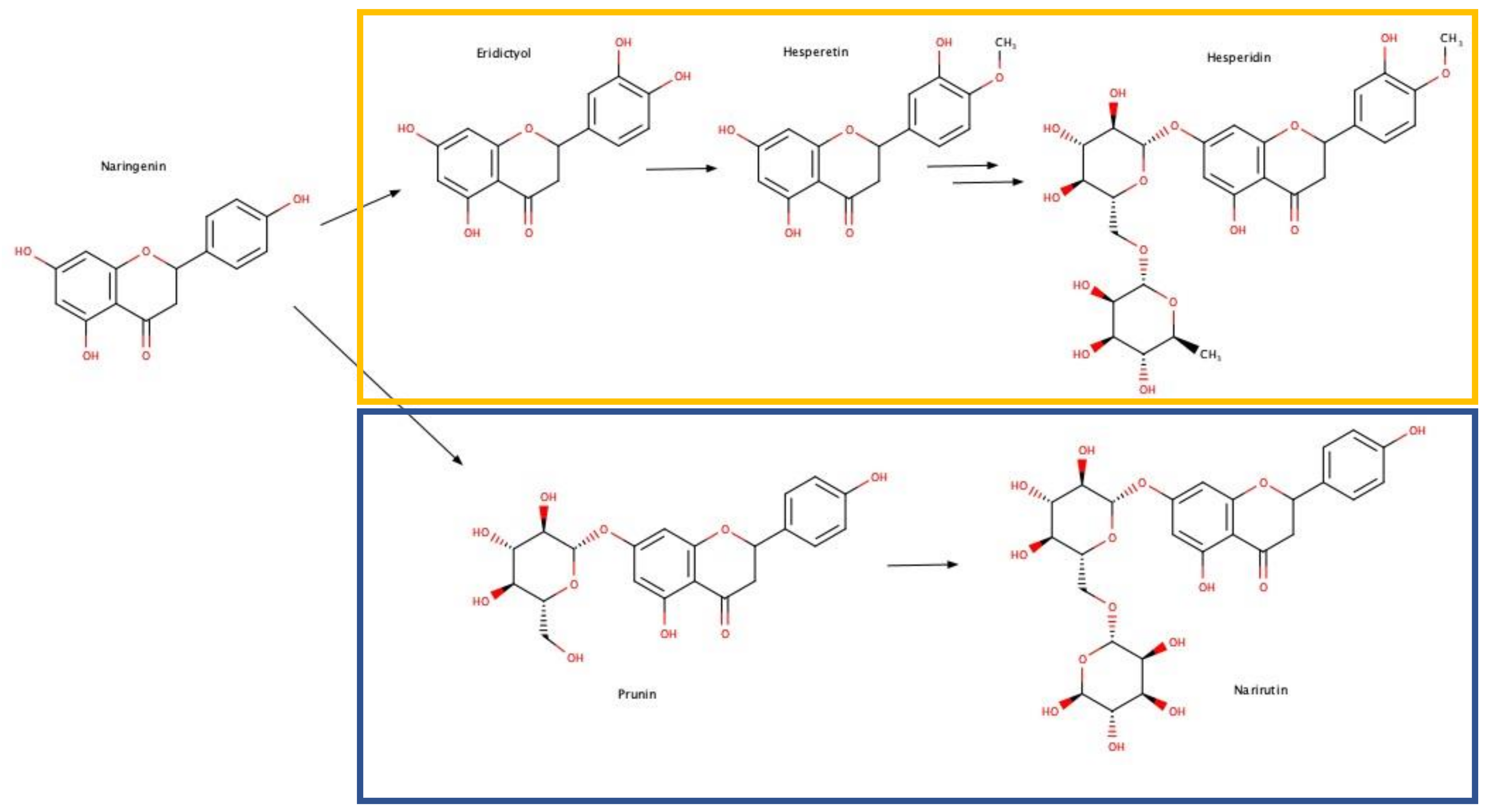
3.2. Identification and Quantification of Phytochemicals in Leaves
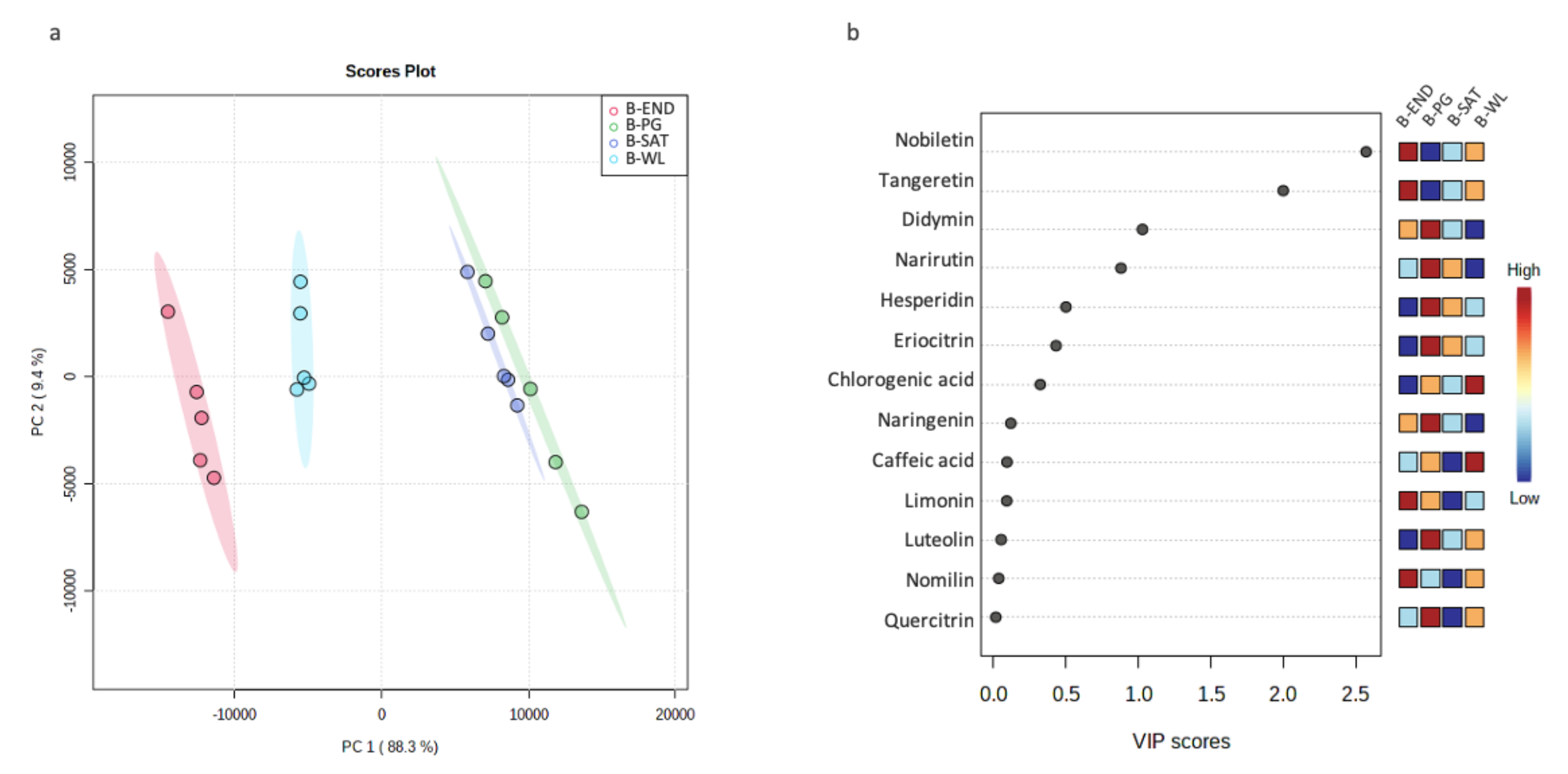
3.3. Chemotaxonomic Characterization of Bearing Branches
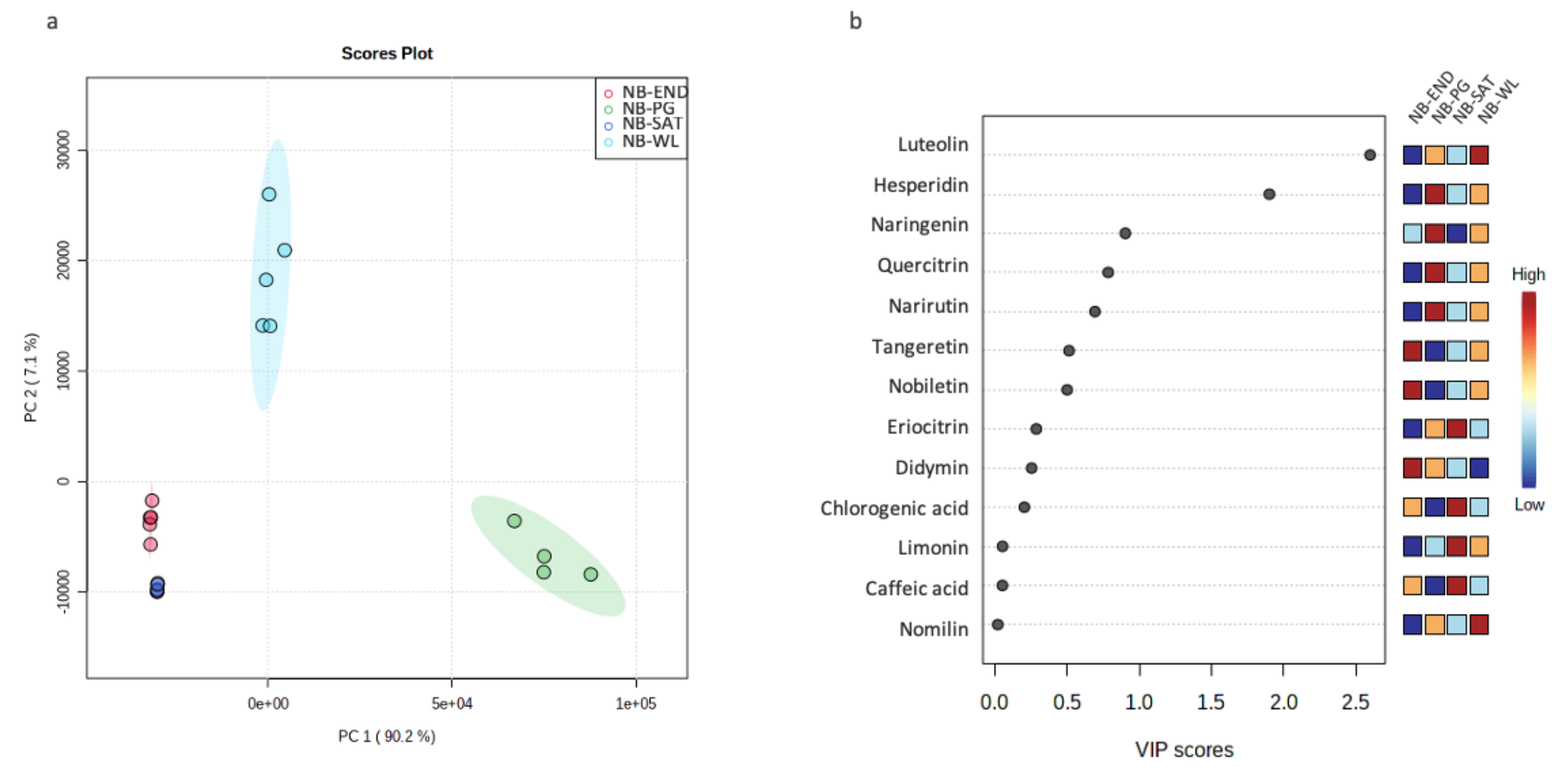
3.4. Chemotaxonomic Characterization of Non-Bearing Branches
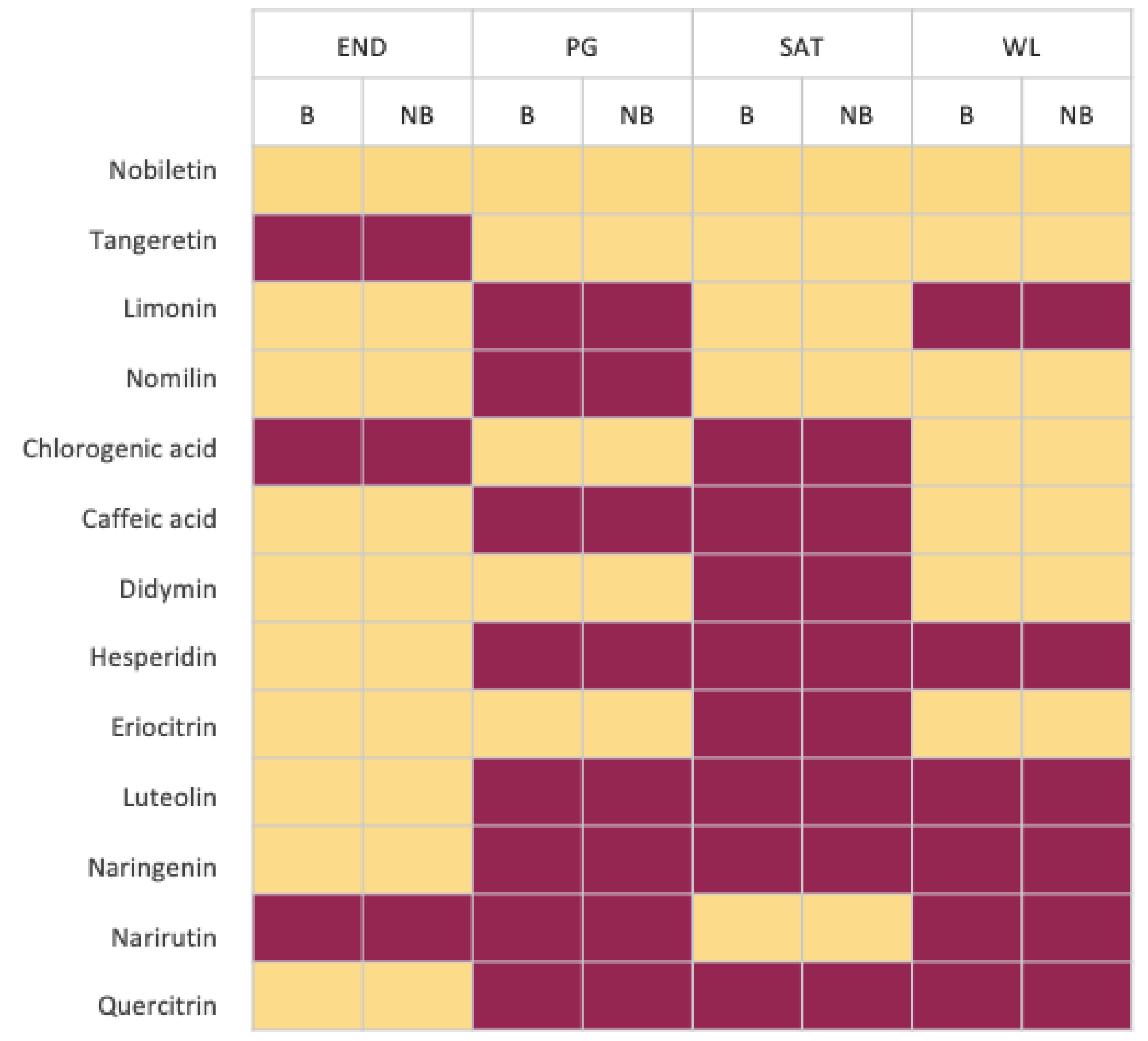
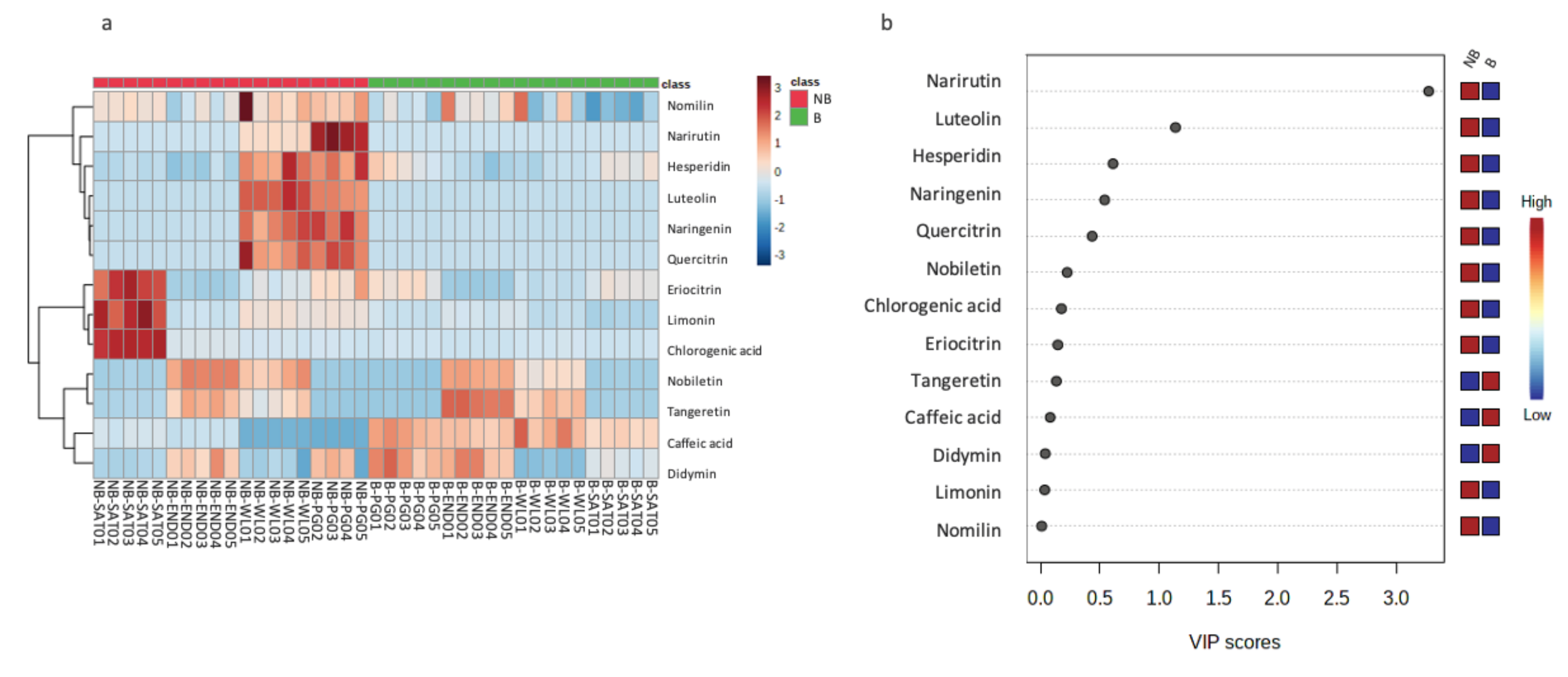
3.5. Metobolites’ Profile Comparison between Leaves from Bearing and Non-Bearing Branches
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Available online: http://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 5 October 2021).
- Shalom, L.; Samuels, S.; Zur, N.; Shlizerman, L.; Zemach, H.; Weissberg, M.; Ophir, R.; Blumwald, E.; Sadka, A. Alternate Bearing in Citrus: Changes in the Expression of Flowering Control Genes and in Global Gene Expression in ON- versus OFF-Crop Trees. PLoS ONE 2012, 7, e46930. [Google Scholar] [CrossRef] [Green Version]
- Agust, M.; Almela, V.; Pons, J. Effects of girdling on alternate bearing in citrus. J. Hortic. Sci. 1992, 67, 203–210. [Google Scholar] [CrossRef]
- Talon, M.; Tadeo, F.R.; Ben-Cheikh, W.; Gomez-Cadenas, A.; Mehouachi, J.; Pérez-Botella, J.; Primo-Millo, E. Hormonal regulation of fruit set and abscission in citrus: Classical concepts and new Evidence. Acta Hortic. 1998, 463, 209–217. [Google Scholar] [CrossRef]
- Bower, J.P.; Cutting, J.G.M.; Lovatt, C.J.; Blanke, M.M. Interaction of plant growth regulator and carbohydrate in flowering and fruit set. Acta Hortic. 1990, 425–434. [Google Scholar] [CrossRef]
- Davenport, T.L. Citrus Flowering. In Horticultural Reviews; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 349–408. [Google Scholar]
- Peña, L.; Martín-Trillo, M.; Juárez, J.; Pina, J.A.; Navarro, L.; Martínez-Zapater, J.M. Constitutive expression of Arabidopsis LEAFY or APETALA1 genes in citrus reduces their generation time. Nat. Biotechnol. 2001, 19, 263–267. [Google Scholar] [CrossRef]
- Pillitteri, L.J.; Lovatt, C.J.; Walling, L.L. Isolation and characterization of LEAFY and APETALA1 homologues from Citrus sinensis L. Osbeck “Washington”. J. Am. Soc. Hortic. Sci. 2004, 129, 846–856. [Google Scholar] [CrossRef] [Green Version]
- Araki, T. Transition from vegetative to reproductive phase. Curr. Opin. Plant Biol. 2001, 4, 63–68. [Google Scholar] [CrossRef]
- Moss, G.I. Effect of fruit on flowering in relation to biennial bearing in sweet orange (Citrus sinensis). J. Hortic. Sci. 1971, 46, 177–184. [Google Scholar] [CrossRef]
- Wada, K.C.; Takeno, K. Stress-induced flowering. Plant Signal. Behav. 2010, 5, 944–947. [Google Scholar] [CrossRef] [PubMed]
- Takeno, K. Stress-Induced Flowering. Abiotic Stress Responses Plants 2012, 331–345. [Google Scholar] [CrossRef]
- Kazan, K.; Lyons, R. The link between flowering time and stress tolerance. J. Exp. Bot. 2016, 67, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Zagoskina, N.V.; Kazantseva, V.V.; Fesenko, A.N.; Shirokova, A.V. Accumulation of Phenolic Compounds at the Initial Steps of Ontogenesis of Fagopyrum esculentum Plants That Differ in Their Ploidy Levels. Biol. Bull. 2018, 45, 171–178. [Google Scholar] [CrossRef]
- Coley, P.D.; Bryant, J.P.; Chapin, F.S. Resource Availability and Plant Antiherbivore Defense. Science 1985, 230, 895–899. [Google Scholar] [CrossRef] [Green Version]
- Waterman, P.; Mole, S. Analysis of Phenolic Plant Metabolites; Blackwell Scientific Publications: Oxford, UK, 1994. [Google Scholar]
- Grace, S.G.; Logan, B.A. Energy Dissipation and Radical Scavenging by the Plant Phenylpropanoid Pathway. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 1499–1510. [Google Scholar] [CrossRef] [Green Version]
- Rivas, F.; Fornes, F.; Agustí, M. Girdling induces oxidative damage and triggers enzymatic and non-enzymatic antioxidative defences in Citrus leaves. Environ. Exp. Bot. 2008, 64, 256–263. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Cheynier, V. Polyphenols in foods are more complex than often thought. Am. J. Clin. Nutr. 2005, 81, 223S–229S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClure, J.W. Physiology and Functions of Flavonoids. In The Flavonoids; Harbone, J.B., Mabry, T.J., Mabry, H., Eds.; Springer: Boston, MA, USA, 1975; pp. 970–1055. [Google Scholar]
- Gualdani, R.; Cavalluzzi, M.; Lentini, G.; Habtemariam, S. The Chemistry and Pharmacology of Citrus Limonoids. Molecules 2016, 21, 1530. [Google Scholar] [CrossRef]
- Shi, Y.S.; Zhang, Y.; Li, H.T.; Wu, C.H.; El-Seedi, H.R.; Ye, W.K.; Wang, Z.W.; Li, C.B.; Zhang, X.F.; Kai, G.Y. Limonoids from Citrus: Chemistry, anti-tumor potential, and other bioactivities. J. Funct. Foods 2020, 75, 104213. [Google Scholar] [CrossRef]
- Roy, A.; Saraf, S. Limonoids: Overview of Significant Bioactive Triterpenes Distributed in Plants Kingdom. Biol. Pharm. Bull. 2006, 29, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Hafeez, F.; Akram, W.; Shaalan, E.A.-S. Mosquito larvicidal activity of citrus limonoids against Aedes albopictus. Parasitol. Res. 2011, 109, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Warikoo, R.; Ray, A.; Sandhu, J.K.; Samal, R.; Wahab, N.; Kumar, S. Larvicidal and irritant activities of hexane leaf extracts of Citrus sinensis against dengue vector Aedes aegypti L. Asian Pac. J. Trop. Biomed. 2012, 2, 152–155. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.K.; Dangles, O. A comprehensive review on flavanones, the major citrus polyphenols. J. Food Compos. Anal. 2014, 33, 85–104. [Google Scholar] [CrossRef]
- Yu, X.; Shi, D.; Zhi, X.; Li, Q.; Yao, X.; Xu, H. Synthesis and quantitative structure-activity relationship (QSAR) study of C7-oxime ester derivatives of obacunone as insecticidal agents. RSC Adv. 2015, 5, 31700–31707. [Google Scholar] [CrossRef]
- Hasegawa, S.; Berhow, M.A.; Manners, G.D. Citrus limonoid research: An overview. ACS Symp. Ser. 2000, 758, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Celano, R.; Campone, L.; Pagano, I.; Carabetta, S.; Di Sanzo, R.; Rastrelli, L.; Piccinelli, A.L.; Russo, M. Characterisation of nutraceutical compounds from different parts of particular species of Citrus sinensis ‘Ovale Calabrese’ by UHPLC-UV-ESI-HRMS. Nat. Prod. Res. 2019, 33, 244–251. [Google Scholar] [CrossRef]
- Russo, M.; Arigò, A.; Calabrò, M.L.; Farnetti, S.; Mondello, L.; Dugo, P. Bergamot (Citrus bergamia Risso) as a source of nutraceuticals: Limonoids and flavonoids. J. Funct. Foods 2016, 20, 10–19. [Google Scholar] [CrossRef]
- Wang, F.; Yu, X.; Liu, X.; Shen, W.; Zhu, S.; Zhao, X. Temporal and spatial variations on accumulation of nomilin and limonin in the pummelos. Plant Physiol. Biochem. 2016, 106, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.D.; Souza, A.L.; Gerszten, R.E.; Clish, C.B. Targeted metabolomics. Curr. Protoc. Mol. Biol. 2012, 1, 30.2.1–30.2.24. [Google Scholar] [CrossRef]
- Xi, W.; Zhang, Y.; Sun, Y.; Shen, Y.; Ye, X.; Zhou, Z. Phenolic composition of Chinese wild mandarin (Citrus reticulata Balnco.) pulps and their antioxidant properties. Ind. Crops Prod. 2014, 52, 466–474. [Google Scholar] [CrossRef]
- Safdar, M.N.; Kausar, T.; Jabbar, S.; Mumtaz, A.; Ahad, K.; Saddozai, A.A. Extraction and quantification of polyphenols from kinnow (Citrus reticulate L.) peel using ultrasound and maceration techniques. J. Food Drug Anal. 2017, 25, 488–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Ou, P.; Hasegawa, S.; Herman, Z.; Fong, C.H. Limonoid biosynthesis in the stem of Citrus limon. Phytochemistry 1988, 27, 115–118. [Google Scholar] [CrossRef]
- Gerolino, E.; Chierrito, T.; Santana-Filho, A.; Rorigues, S.; Gonçalves, R.; Oliveira, A. Evaluation of limonoid production in suspension cell culture of Citrus sinensis. Rev. Bras. Farmacogn. 2015, 56. [Google Scholar] [CrossRef] [Green Version]
- Berim, A.; Gang, D.R. Methoxylated flavones: Occurrence, importance, biosynthesis. Phytochem. Rev. 2016, 15, 363–390. [Google Scholar] [CrossRef]
- Abad-García, B.; Garmón-Lobato, S.; Berrueta, L.A.; Gallo, B.; Vicente, F. On line characterization of 58 phenolic compounds in Citrus fruit juices from Spanish cultivars by high-performance liquid chromatography with photodiode-array detection coupled to electrospray ionization triple quadrupole mass spectrometry. Talanta 2012, 99, 213–224. [Google Scholar] [CrossRef]
- Mou, J.; Zhang, Z.; Qiu, H.; Lu, Y.; Zhu, X.; Fan, Z.; Zhang, Q.; Ye, J.; Fernie, A.R.; Cheng, Y.; et al. Multiomics-based dissection of citrus flavonoid metabolism using a Citrus reticulata × Poncirus trifoliata population. Hortic. Res. 2021, 8, 56. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, F.; Lian, Y.; Xiao, H.; Zheng, J. Biosynthesis of citrus flavonoids and their health effects. Crit. Rev. Food Sci. Nutr. 2020, 60, 566–583. [Google Scholar] [CrossRef]
- Frydman, A.; Weisshaus, O.; Bar-Peled, M.; Huhman, D.V.; Sumner, L.W.; Marin, F.R.; Lewinsohn, E.; Fluhr, R.; Gressel, J.; Eyal, Y. Citrus fruit bitter flavors: Isolation and functional characterization of the gene Cm1,2RhaT encoding a 1,2 rhamnosyltransferase, a key enzyme in the biosynthesis of the bitter flavonoids of citrus. Plant J. 2004, 40, 88–100. [Google Scholar] [CrossRef]
- Chung, S.W.; Yu, D.J.; Oh, H.D.; Ahn, J.H.; Huh, J.H.; Lee, H.J. Transcriptional regulation of abscisic acid biosynthesis and signal transduction, and anthocyanin biosynthesis in ‘Bluecrop’ highbush blueberry fruit during ripening. PLoS ONE 2019, 14, e0220015. [Google Scholar] [CrossRef] [Green Version]
- Monselise, S.P.; Goldschmidt, E.E. Alternate bearing in citrus and ways of control. In Proceedings of the International Society of Citriculture, Tokyo, Japan, 9–12 November 1981; pp. 239–242. [Google Scholar]



 means does not differ significantly using Wilcoxon test, p ≤ 0.05, and
means does not differ significantly using Wilcoxon test, p ≤ 0.05, and  means differs significantly using Wilcoxon test, p ≤ 0.05.
means does not differ significantly using Wilcoxon test, p ≤ 0.05, and means differs significantly using Wilcoxon test, p ≤ 0.05.
means differs significantly using Wilcoxon test, p ≤ 0.05.
means does not differ significantly using Wilcoxon test, p ≤ 0.05, and means differs significantly using Wilcoxon test, p ≤ 0.05.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | [M-H]− | [M-H]− ⟶ m/z | DW | DP | EP | CE | CXP | Rt (min) |
|---|---|---|---|---|---|---|---|---|
| Protocatechuic acid | 153 | 108.8 | 150 | −40 | −10 | −22 | −15 | 9.4 |
| 90.7 | 150 | −40 | −10 | −42 | −1 | |||
| Chlorogenic acid | 353.7 | 190.8 | 150 | −50 | −10 | −24 | −31 | 11.1 |
| 84.7 | 150 | −50 | −10 | −68 | −9 | |||
| Caffeic acid | 179 | 133.3 | 150 | −45 | −10 | −40.7 | −21.1 | 12.7 |
| 88.7 | 150 | −45 | −10 | −48 | −14.1 | |||
| Eriocitrin | 595.2 | 286.6 | 150 | −80 | −10 | −38 | −9 | 14.5 |
| 134.6 | 150 | −80 | −10 | −90 | −19 | |||
| Narirutin | 579.2 | 150.5 | 150 | −95 | −10 | −75 | −20.3 | 15.9 |
| 270.8 | 150 | −95 | −10 | −39.9 | −8.4 | |||
| Naringin | 579.2 | 270.8 | 150 | −105 | −10 | −54 | −7 | 16.3 |
| 150.7 | 150 | −122.03 | −10 | −75.03 | −21 | |||
| Quercitrin | 447.1 | 300.5 | 150 | −90 | −10 | −36 | −11 | 16.3 |
| 270.8 | 150 | −90 | −10 | −66 | −41 | |||
| Hesperidin | 609.2 | 300.8 | 150 | −85 | −10 | −46 | −49 | 16.5 |
| 285.8 | 150 | −85 | −10 | −70 | −41 | |||
| 163.5 | 150 | −85 | −10 | −88 | −25 | |||
| Neohesperidin | 609.2 | 300.7 | 150 | −115 | −10 | −56 | −9.32 | 16.9 |
| 285.7 | 150 | −115 | −10 | −72.89 | −42.3 | |||
| 150.5 | 150 | −115 | −10 | −100 | −21 | |||
| Didymin | 593.2 | 284.8 | 150 | −100 | −10 | −48 | −45 | 19.8 |
| 164 | 150 | −100 | −10 | −92 | −27 | |||
| Poncirin | 593.3 | 284.8 | 150 | −120 | −10 | −54 | −9 | 20.0 |
| 163.4 | 150 | −120 | −10 | −92 | −23 | |||
| Luteolin | 285 | 150.6 | 150 | −85 | −10 | −41.6 | −20.9 | 20.5 |
| 132.9 | 150 | −85 | −10 | −54 | −17 | |||
| Quercetin | 301 | 150.8 | 150 | −75 | −10 | −36 | −21 | 20.7 |
| 178.7 | 150 | −75 | −10 | −28 | −27 | |||
| 120.8 | 150 | −75 | −10 | −44 | −16.3 | |||
| Kaempferol | 284.9 | 92.9 | 150 | −80 | −10 | −51.4 | −14.1 | 23.1 |
| 116.88 | 150 | −80 | −10 | −67 | −18.8 | |||
| Naringenin | 271 | 150.7 | 150 | −60 | −10 | −28.87 | −23.14 | 23.2 |
| 118.7 | 150 | −60 | −10 | −40.9 | −16.7 | |||
| 106.6 | 150 | −60 | −10 | −37.8 | −16.9 | |||
| Analyte | [M-H]+ | [M-H]+⟶ m/z | DW | DP | EP | CE | CXP | Rt (min) |
| Bergapten | 217.2 | 202 | 150 | 64.1 | 10 | 30.9 | 10.9 | 25.8 |
| 174.3 | 150 | 64.1 | 10 | 41.4 | 6.1 | |||
| 90.1 | 150 | 64.1 | 10 | 57.9 | 3.1 | |||
| Limonin | 471.2 | 425.3 | 150 | 71 | 10 | 28.6 | 22 | 26.7 |
| 161.1 | 150 | 71 | 10 | 37.7 | 12.7 | |||
| Nobiletin | 403.1 | 373.1 | 150 | 61 | 10 | 41 | 14 | 26.7 |
| 388.1 | 150 | 61 | 10 | 31 | 16 | |||
| Nomilin | 515.2 | 469.3 | 150 | 44.9 | 10 | 20.18 | 11.4 | 27.9 |
| 161.1 | 150 | 44.9 | 10 | 35.1 | 11.2 | |||
| Tangeretin | 372.97 | 343 | 150 | 71 | 10 | 39 | 26 | 28.4 |
| 297.1 | 150 | 71 | 10 | 47 | 10 | |||
| 211.1 | 150 | 71 | 10 | 60.87 | 7.3 |
| Compound | B-SAT | B-END | B-WL | B-PG |
|---|---|---|---|---|
| (μg/g) * | ||||
| Chlorogenic Acid | 3.88 b ± 0.40 | 3.53 b ± 0.37 | 6.70 a ± 0.78 | 5.04 b ± 1.39 |
| Caffeic Acid | 3.87 b ± 0.21 | 4.34 ab ± 0.23 | 5.38 a ± 1.05 | 5.18 a ± 0.51 |
| Didymin | 10.57 b ± 1.11 | 20.19 a ± 3.27 | 3.30 c ± 0.43 | 21.17 a ± 3.76 |
| Hesperidin | 3008.58 a ± 151.89 | 2512.39 b ± 182.61 | 2624.31 b ± 114.09 | 3152.43 a ± 298.13 |
| Eriocitrin | 47.93 a ± 6.42 | 13.90 c ± 1.19 | 29.85 b ± 2.44 | 57.24 a ± 8.02 |
| Luteolin | 1.45 a ± 0.05 | 1.21 a ± 0.08 | 1.81 a ± 0.28 | 2.31 a ± 1.26 |
| Naringenin | 0.38 b ± 0.02 | 0.57 a ± 0.01 | 0.34 b ± 0.01 | 0.63 a ± 0.15 |
| Narirutin | 87.03 a ± 12.68 | 50.08 b ± 7.58 | 17.58 c ± 7.58 | 98.20 a ± 14.35 |
| Quercitrin | 2.29 b ± 0.07 | 2.43 b ± 0.13 | 2.98 ab ± 0.55 | 3.69 a ± 0.61 |
| Nobiletin | 258.17 c ± 16.48 | 1121.36 a ± 34.94 | 747.15 b ± 55.16 | 208.17 c± 13.63 |
| Tangeretin | 64.96 c ± 6.15 | 731.28 a ± 30.15 | 475.84 b ± 56.24 | 34.34 c ± 5.01 |
| Limonin | 0.45 c ± 0.04 | 0.88 a ± 0.08 | 0.72 b ± 0.12 | 0.76 ab ± 0.08 |
| Nomilin | 0.42 b ± 0.03 | 0.62 a ± 0.08 | 0.57 ab ± 0.13 | 0.52 ab ± 0.04 |
| Compound | NB-SAT | NB-END | NB-WL | NB-PG |
|---|---|---|---|---|
| (μg/g) * | ||||
| Chlorogenic Acid | 126.02 a ± 6.40 | 11.31 b ± 0.62 | 6.75 b ± 1.41 | 6.35 b ± 1.50 |
| Caffeic Acid | 2.32 a ± 0.12 | 2.00 b ± 0.09 | 0.12 c ± 0.04 | 0.04 c ± 0.02 |
| Didymin | 6.44 bc ± 0.47 | 17.40 a ± 2.97 | 5.03 c ± 2.56 | 14.07 ab ± 8.77 |
| Hesperidin | 2591.54 b ± 55.63 | 2433.21 b ± 179.79 | 4224.72 a ± 450.98 | 4270.21 a ± 400.09 |
| Eriocitrin | 132.39 a ± 15.01 | 17.09 d ± 1.82 | 39.62 c ± 5.34 | 69.98 b ± 17.91 |
| Luteolin | 3.16 c ± 0.60 | 2.13 c ± 0.15 | 2659.07 a ± 267.40 | 2023.38 b ± 121.10 |
| Naringenin | 0.32 b ± 0.02 | 0.68 b ± 0.12 | 509.61 a ± 100.14 | 574.03 a ± 101.15 |
| Narirutin | 97.45 c ± 8.08 | 62.85 c ± 5.04 | 10049.66 b ± 1005.64 | 42198.36 a ± 3700.22 |
| Quercitrin | 2.80 b ± 0.21 | 2.44 b ± 0.16 | 350.39 a ± 122.31 | 362.45 a ± 50.83 |
| Nobiletin | 290.48 e ± 24.07 | 1257.23 a ± 87.18 | 1028.88 b ± 151.80 | 228.71 c ± 12.86 |
| Tangeretin | 99.89 c ± 10.05 | 517.97 a ± 78.41 | 347.23 b ± 82.59 | 33.27 c ± 5.98 |
| Limonin | 3.40 a ± 0.44 | 0.77 c ± 0.05 | 1.30 b ± 0.09 | 1.28 b ± 0.03 |
| Nomilin | 0.6 ab ± 0.01 | 0.54 b ± 0.05 | 0.19 a ± 0.14 | 0.66 ab ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez Ceraolo, C.; Vázquez, V.; Migues, I.; Cesio, M.V.; Rivas, F.; Heinzen, H. Flavonoids and Limonoids Profiles Variation in Leaves from Mandarin Cultivars and Its Relationship with Alternate Bearing. Agronomy 2022, 12, 121. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010121
Rodríguez Ceraolo C, Vázquez V, Migues I, Cesio MV, Rivas F, Heinzen H. Flavonoids and Limonoids Profiles Variation in Leaves from Mandarin Cultivars and Its Relationship with Alternate Bearing. Agronomy. 2022; 12(1):121. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010121
Chicago/Turabian StyleRodríguez Ceraolo, Cecilia, Valeria Vázquez, Ignacio Migues, María Verónica Cesio, Fernando Rivas, and Horacio Heinzen. 2022. "Flavonoids and Limonoids Profiles Variation in Leaves from Mandarin Cultivars and Its Relationship with Alternate Bearing" Agronomy 12, no. 1: 121. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010121