
Grass–Legume Mixtures Show Potential to Increase Above- and Belowground Biomass Production for Andean Forage-Based Fallows


Abstract
:1. Introduction
- 1.
- To what extent do species mixtures (or bicultures) promote above- and belowground biomass production relative to monocultures across differing soil conditions?
- 2.
- Are there specific bicultures (or component species) that are especially promising for supporting aboveground vs. belowground biomass production (and associated implications for forage vs. soil improvement)?
2. Materials and Methods
2.1. Study Site and Soil Collection
2.2. Experimental Design and Establishment
2.3. Plant Harvest and Data Collection
2.4. Calculating the Relative Yield Total
2.5. Statistical Approach
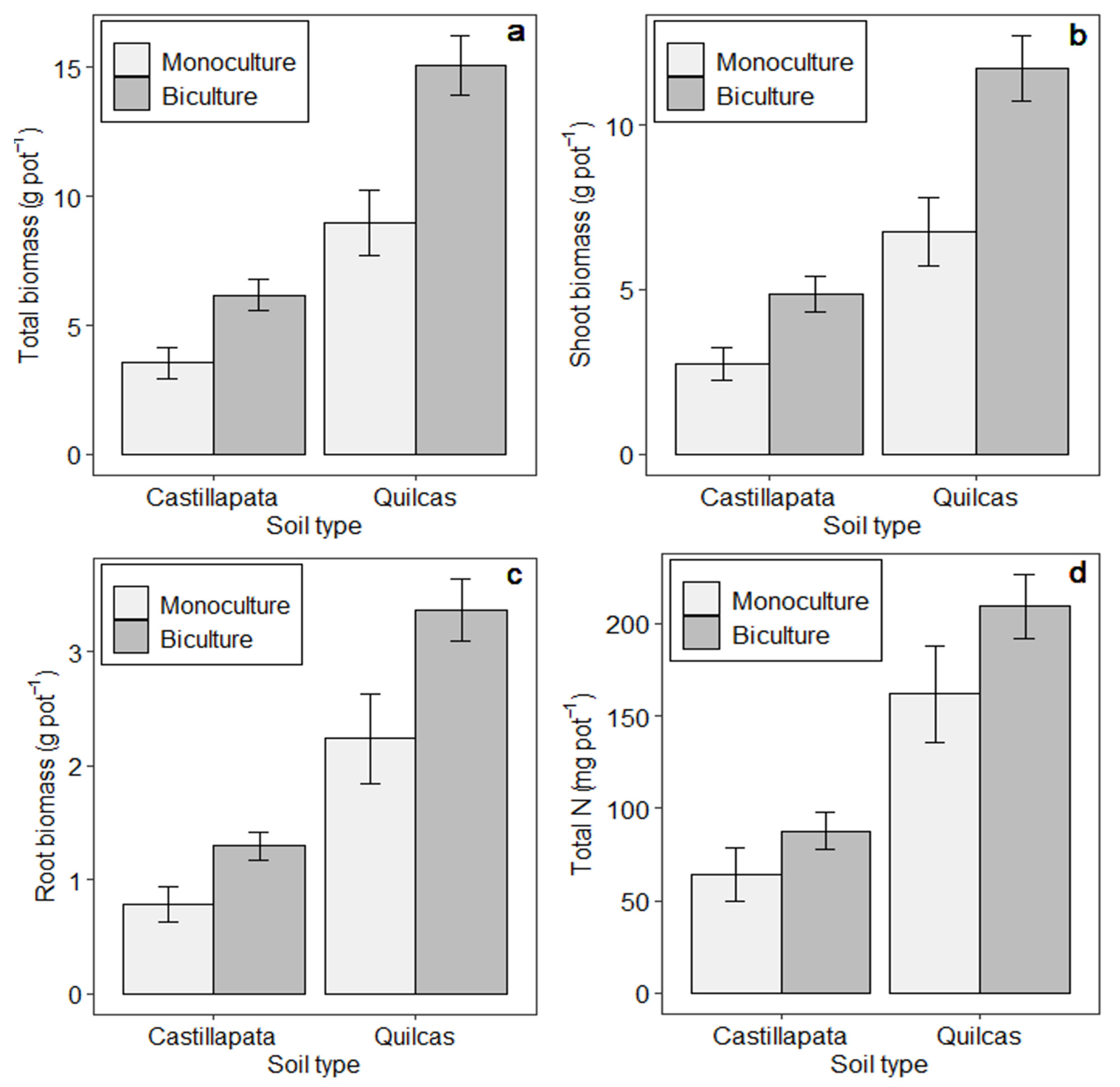
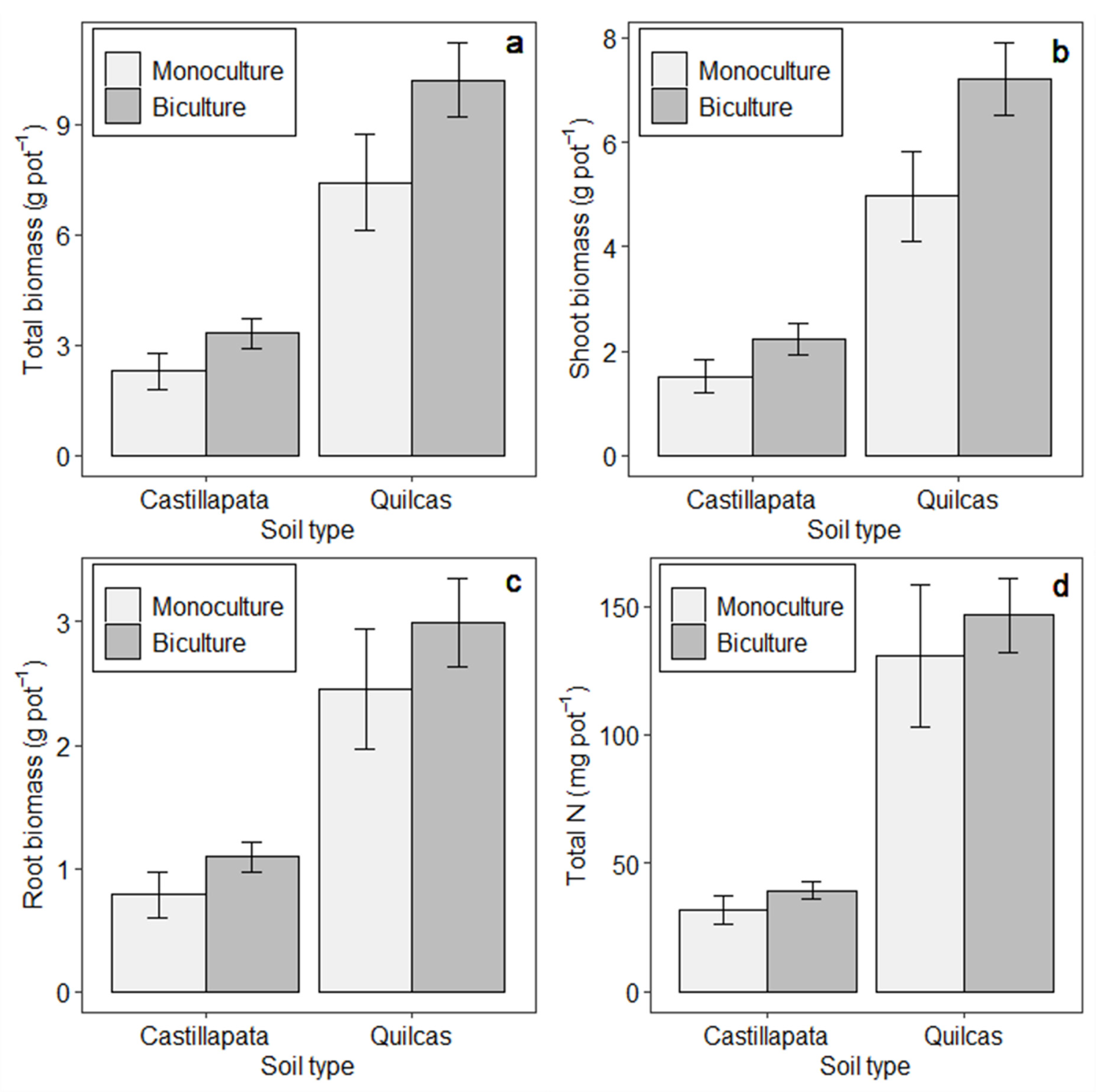
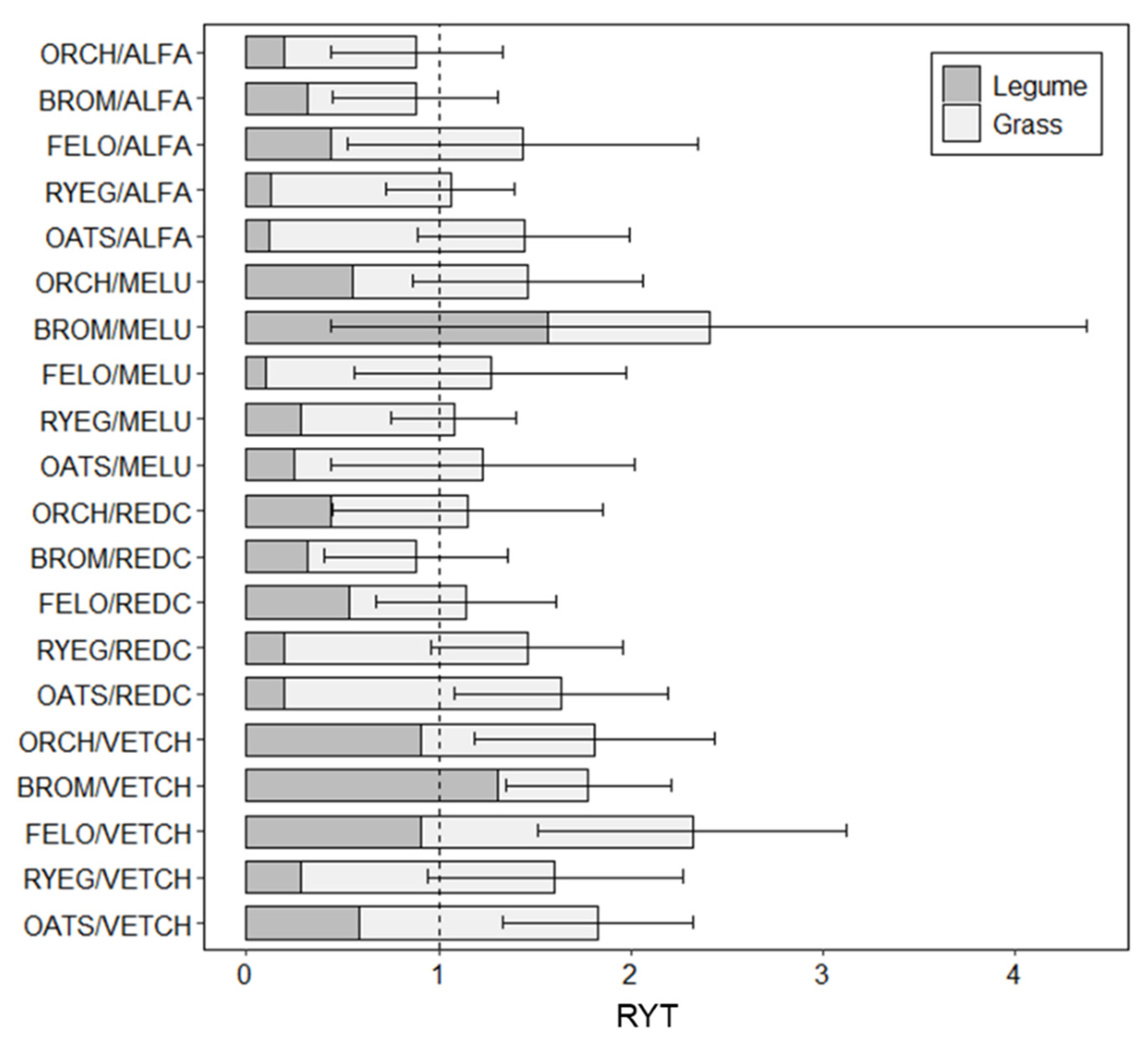
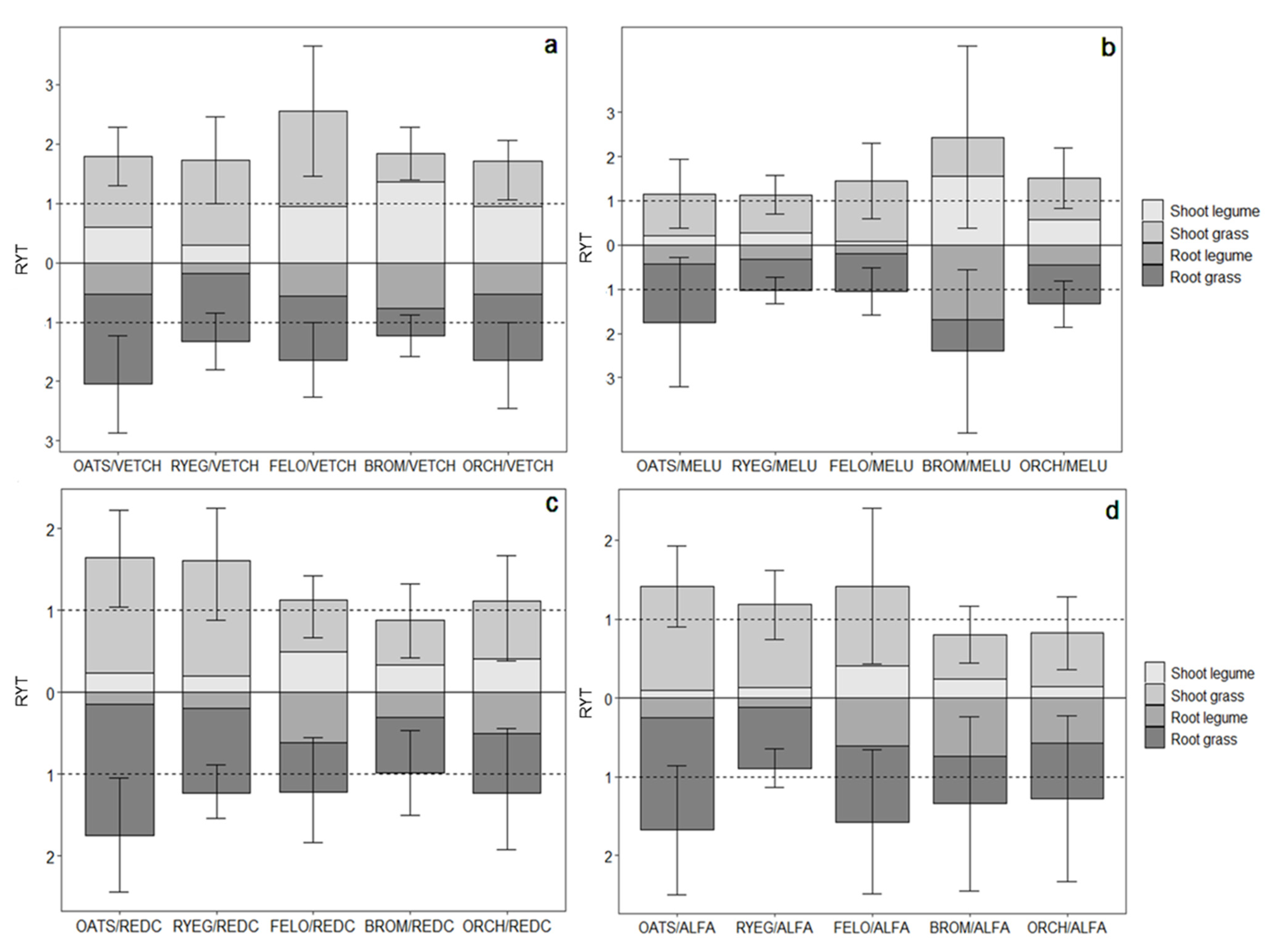
3. Results
3.1. Treatment and Soil Effects on Biomass Production and N Uptake
3.2. Comparing RYT across Mixtures
4. Discussion
4.1. Overall Performance of Bicultures vs. Monocultures
4.2. Comparing the Performance of Different Bicultures
4.3. Implications for Smallholder Agriculture Systems
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vanek, S.J.; Meza, K.; Ccanto, R.; Olivera, E.; Scurrah, M.; Fonte, S.J. Participatory design of improved forage/fallow options across soil gradients with farmers of the Central Peruvian Andes. Agric. Ecosyst. Environ. 2020, 300, 106933. [Google Scholar] [CrossRef]
- Rolando, J.L.; Dubeux, J.C.B.; Ramirez, D.A.; Ruiz-Moreno, M.; Turin, C.; Mares, V.; Lynn, E.; Sollenberger; Quiroz, R. Land Use effects on soil fertility and nutrient cycling in the Peruvian High-Andean puna grasslands. Soil Sci. Soc. Am. J. 2018, 82, 463–474. [Google Scholar] [CrossRef]
- Pestalozzi, H. Sectoral fallow systems and the management of soil fertility: The rationality of indigenous knowledge in the High Andes of Bolivia. Mt. Res. Dev. 2000, 20, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Abadín, J.; González-Prieto, S.J.; Sarmiento, L.; Villar, M.C.; Carballas, T. Successional dynamics of soil characteristics in a long fallow agricultural system of the high tropical Andes. Soil Biol. Biochem. 2002, 34, 1739–1748. [Google Scholar] [CrossRef] [Green Version]
- Sarmiento, L. Water balance and soil loss under long fallow agriculture in the Venezuelan Andes. Mt. Res. Dev. 2000, 20, 246–253. [Google Scholar] [CrossRef] [Green Version]
- Fonte, S.J.; Vanek, S.J.; Oyarzun, P.; Parsa, S.; Quintero, D.C.; Rao, I.M.; Lavelle, P. Pathways to Agroecological Intensification of Soil Fertility Management by Smallholder Farmers in the Andean Highlands; Elsevier Inc.: Amsterdam, The Netherlands, 2012; p. 116. [Google Scholar] [CrossRef]
- Sanchez, P.A. Improved fallows come of age in the tropics. Agrofor. Syst. 1999, 47, 3–12. [Google Scholar] [CrossRef]
- Nyamadzawo, G.; Chikowo, R.; Nyamugafata, P.; Nyamangara, J.; Giller, K.E. Soil organic carbon dynamics of improved fallow-maize rotation systems under conventional and no-tillage in Central Zimbabwe. Nutr. Cycl. Agroecosyst. 2008, 81, 85–93. [Google Scholar] [CrossRef]
- Qingjie, W.; Hao, C.; Hongwen, L.; Wenying, L.; Xiaoyan, W.; McHugh, A.D.; He, J.; Huanwen, G. Controlled traffic farming with no tillage for improved fallow water storage and crop yield on the Chinese Loess Plateau. Soil Tillage Res. 2009, 104, 192–197. [Google Scholar] [CrossRef]
- Wezel, A.; Soboksa, G.; McClelland, S.; Delespesse, F.; Boissau, A. The blurred boundaries of ecological, sustainable, and agroecological intensification: A review. Agron. Sustain. Dev. 2015, 35, 1283–1295. [Google Scholar] [CrossRef]
- Sarmiento, L.; Abadín, J.; González-Prieto, S.; Carballas, T. Assessing and modeling the role of the native legume Lupinus meridanus in fertility restoration in a heterogeneous mountain environment of the tropical Andes. Agric. Ecosyst. Environ. 2012, 159, 29–39. [Google Scholar] [CrossRef]
- Sarmiento, L.; Llambí, L.D.; Escalona, A.; Marquez, N. Vegetation patterns, regeneration rates and divergence in an old-field succession of the high tropical Andes. Plant Ecol. 2003, 166, 145–156. [Google Scholar] [CrossRef]
- de Valença, A.W.; Vanek, S.J.; Meza, K.; Ccanto, R.; Olivera, E.; Scurrah, M.; Lantinga, E.A.; Fonte, S.J. Land use as a driver of soil fertility and biodiversity across an agricultural landscape in the Central Peruvian Andes. Ecol. Appl. 2017, 27, 1138–1154. [Google Scholar] [CrossRef] [PubMed]
- Manning, P.; Van Der Plas, F.; Soliveres, S.; Allan, E.; Maestre, F.T.; Mace, G.; Whittingham, M.J.; Fischer, M. Redefining ecosystem multifunctionality. Nat. Ecol. Evol. 2018, 2, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Cabel, J.F.; Oelofse, M. An indicator framework for assessing agroecosystem resilience. Ecol. Soc. 2012, 17. [Google Scholar] [CrossRef]
- Vandermeer, J.H. The Ecology of Intercropping; Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Kahmen, A.; Renker, C.; Unsicker, S.B.; Buchmann, N. Niche complementarity for nitrogen: An explanation for the biodiversity and ecosystem functioning relationship? Ecology 2006, 87, 1244–1255. [Google Scholar] [CrossRef]
- Brooker, R.W.; Bennett, A.E.; Cong, W.F.; Daniell, T.J.; George, T.S.; Hallett, P.D.; White, W.J.; Hawes, C. Improving intercropping: A synthesis of research in agronomy, plant physiology and ecology. New Phytologist. 2015, 206, 107–117. [Google Scholar] [CrossRef]
- Blesh, J. Functional traits in cover crop mixtures: Biological nitrogen fixation and multifunctionality. J. Appl. Ecol. 2017, 55, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Isbell, F.; Adler, P.R.; Eisenhauer, N.; Fornara, D.; Kimmel, K.; Kremen, C.; Letourneau, D.K.; Liebman, M.; Polley, H.W.; Quijas, S.; et al. Benefits of increasing plant diversity in sustainable agroecosystems. J. Ecol. 2017, 105, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Schipanski, M.E.; Drinkwater, L.E. Nitrogen fixation in annual and perennial legume-grass mixtures across a fertility gradient. Plant Soil. 2012, 357, 147–159. [Google Scholar] [CrossRef]
- Wick, A.; Berti, M.; Lawley, Y.; Liebig, M. Integration of Annual and Perennial Cover Crops for Improving Soil Health; Elsevier Inc.: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Li, L.; Zhang, L.; Zhang, F. Crop Mixtures and the Mechanisms of Overyielding. Encycl. Biodivers. 2013, 2, 382–395. [Google Scholar] [CrossRef]
- Fornara, D.A.; Tilman, D. Plant functional composition influences rates of soil carbon and nitrogen accumulation. J. Ecol. 2008, 96, 314–322. [Google Scholar] [CrossRef]
- Marshall, A.H.; Collins, R.P.; Humphreys, M.W.; Scullion, J. A new emphasis on root traits for perennial grass and legume varieties with environmental and ecological benefits. Food Energy Secur. 2016, 5, 26–39. [Google Scholar] [CrossRef] [Green Version]
- Rasse, D.P.; Rumpel, C.; Dignac, M.F. Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant Soil 2005, 269, 341–356. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Trumbore, S.E. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeHaan, L.R.; Weisberg, S.; Tilman, D.; Fornara, D. Agricultural and biofuel implications of a species diversity experiment with native perennial grassland plants. Agric. Ecosyst. Environ. 2010, 137, 33–38. [Google Scholar] [CrossRef]
- García, S. Evaluating the Resource Management Strategies of the Agro-Ecosystems in Farm Communities of the Mantaro Valley, Central Andes of Peru. Ph.D. Dissertation, Katholieke Universiteit Lueven, Lueven, Belgium, 2011. [Google Scholar]
- Acker, J.; Leptoukh, G. Online Analysis Enhances Use of NASA Earth Science Data. Eos Trans. Am. Geophys. Union 2007, 88, 14–17. Available online: https://giovanni.gsfc.nasa.gov/giovanni/ (accessed on 21 November 2021). [CrossRef]
- Bouyoucos, G.J. Hydrometer Method Improved for Making Particle Size Analyses of Soils 1. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Weil, R.R.; Islam, K.R.; Stine, M.A.; Gruver, J.B.; Samson-Liebig, S.E. Estimating active carbon for soil quality assessment: A simplified method for laboratory and field use. Am. J. Altern. Agric. 2003, 18, 3–17. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Australia, 2021. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Estimated Marginal Means, Aka Least-Squares Means. R Package Version 2019, 1.3. 2. Available online: https://cran.r-project.org/web/packages/emmeans/index.html (accessed on 6 December 2021).
- Yu, Y.; Stomph, T.J.; Makowski, D.; Van der Werf, W. Temporal niche differentiation increases the land equivalent ratio of annual intercrops: A meta-analysis. Field Crop. Res. 2015, 184, 133–144. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Wright, J.P.; Cadotte, M.W.; Carroll, I.T.; Hector, A.; Srivastava, D.S.; Loreau, M.; Weis, J.J. Impacts of plant diversity on biomass production increase through time because of species complementarity. Proc. Natl. Acad. Sci. USA 2007, 104, 18123–18128. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Isbell, F.; Cowles, J.M. Biodiversity and ecosystem functioning. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 471–493. [Google Scholar] [CrossRef]
- Ren, W.; Hu, L.; Zhang, J.; Sun, C.; Tang, J.; Yuan, Y.; Chen, X. Can positive interactions between cultivated species help to sustain modern agriculture? Front. Ecol. Environ. 2014, 12, 507–514. [Google Scholar] [CrossRef]
- Hooper, D.U.; Dukes, J.S. Overyielding among plant functional groups in a long-term experiment. Ecol. Lett. 2004, 7, 95–105. [Google Scholar] [CrossRef]
- Fornara, D.A.; Tilman, D. Ecological mechanisms associated with the positive diversity-productivity relationship in an N-limited grassland. Ecology 2009, 90, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Faucon, M.P.; Houben, D.; Lambers, H. Plant Functional Traits: Soil and Ecosystem Services. Trends Plant Sci. 2017, 22, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Sainju, U.M.; Singh, B.P.; Whitehead, W.F. Tillage, cover crops, and nitrogen fertilization effects on cotton and sorghum root biomass, carbon, and nitrogen. Agron. J. 2005, 97, 1279–1290. [Google Scholar] [CrossRef] [Green Version]
- Nyfeler, D.; Huguenin-Elie, O.; Suter, M.; Frossard, E.; Lüscher, A. Grass-legume mixtures can yield more nitrogen than legume pure stands due to mutual stimulation of nitrogen uptake from symbiotic and non-symbiotic sources. Agric. Ecosyst. Environ. 2011, 140, 155–163. [Google Scholar] [CrossRef]
- Eaglesham, A.R.J.; Ayanaba, A.; Rao, V.R.; Eskew, D.L. Improving the nitrogen nutrition of maize by intercropping with cowpea. Soil Biol. Biochem. 1981, 13, 169–171. [Google Scholar] [CrossRef]
- Ma, Z.; Chen, H.Y.H. Effects of species diversity on fine root productivity in diverse ecosystems: A global meta-analysis. Glob. Ecol. Biogeogr. 2016, 25, 1387–1396. [Google Scholar] [CrossRef]
- Mueller, K.E.; Tilman, D.; Fornara, D.A.; Hobbie, S.E. Root depth distribution and the diversity-productivity relationship in a long-term grassland experiment. Ecology 2013, 94, 787–793. [Google Scholar] [CrossRef] [Green Version]
- Dhima, K.V.; Lithourgidis, A.S.; Vasilakoglou, I.B.; Dordas, C.A. Competition indices of common vetch and cereal intercrops in two seeding ratio. Field Crop. Res. 2007, 100, 249–256. [Google Scholar] [CrossRef]
- Finney, D.M.; Kaye, J.P. Functional diversity in cover crop polycultures increases multifunctionality of an agricultural system. J. Appl. Ecol. 2017, 54, 509–517. [Google Scholar] [CrossRef]
- Garnier, E. Growth Analysis of Congeneric Annual and Perennial Grass Species. J. Ecol. 1992, 80, 665–675. [Google Scholar] [CrossRef]
- Franzluebbers, A.J. Farming strategies to fuel bioenergy demands and facilitate essential soil services. Geoderma 2015, 259, 251–258. [Google Scholar] [CrossRef]
- King, A.J.; Blesh, J. Crop rotations for increased soil carbon: Perenniality as a guiding principle. Ecol. Appl. 2018, 28, 249–261. [Google Scholar] [CrossRef]
- Dahlin, A.S.; Stenberg, M. Transfer of N from red clover to perennial ryegrass in mixed stands under different cutting strategies. Eur. J. Agron. 2010, 33, 149–156. [Google Scholar] [CrossRef]
- Entz, M.H.; Baron, V.S.; Carr, P.M.; Meyer, D.W.; Smith, S.R.; McCaughey, W.P. Potential of forages to diversify cropping systems in the northern Great Plains. Agron. J. 2002, 94, 240–250. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Papadopoulos, Y.A.; Rodd, A.V.; Grimmett, M.; Fillmor, S.A.E.; Crouse, M.; Prithiviraj, B. Nitrogen fixation and transfer of red clover genotypes under legume–grass forage-based production systems. Nutr. Cycl. Agroecosyst. 2016, 106, 233–247. [Google Scholar] [CrossRef]
- Yuan, Z.Q.; Yu, K.L.; Guan, X.; Fang, C.; Li, M.; Sh, X.Y.; Li, F. Medicago sativa improves soil carbon sequestration following revegetation of degraded arable land in a semi-arid environment on the Loess Plateau, China. Agric. Ecosyst. Environ. 2016, 232, 93–100. [Google Scholar] [CrossRef]
- Hauggaard-Nielsen, H.; Jensen, E.S. Evaluating pea and barley cultivars for complementarity in intercropping at different levels of soil N availability. Field Crop. Res. 2001, 72, 185–196. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Barbercheck, M.; Douglas, M.R.; Finney, D.M.; Haider, K.; Kaye, J.P.; White, C. A framework for evaluating ecosystem services provided by cover crops in agroecosystems. Agric. Syst. 2014, 125, 12–22. [Google Scholar] [CrossRef]
- Sanderson, M.A.; Skinner, R.H.; Barker, D.J.; Edwards, G.R.; Tracy, B.F.; Wedin, D.A. Plant species diversity and management of temperate forage and grazing land ecosystems. Crop Sci. 2004, 44, 1132–1144. [Google Scholar] [CrossRef]
- Christopher, S.F.; Lal, R. Nitrogen management affects carbon sequestration in North American cropland soils. Crit. Rev. Plant Sci. 2007, 26, 45–64. [Google Scholar] [CrossRef]
- Gould, I.J.; Quinton, J.N.; Weigelt, A.; De Deyn, G.B.; Bardgett, R.D. Plant diversity and root traits benefit physical properties key to soil function in grasslands. Ecol. Lett. 2016, 19, 1140–1149. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Zadworny, M. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytologist. 2015, 207, 505–518. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Quilcas | Castillapata |
|---|---|---|
| Sand (%) | 41 | 63 |
| Silt (%) | 30 | 28 |
| Clay (%) | 29 | 9 |
| Texture class | Clay Loam | Sandy Loam |
| N (g kg−1) | 1.3 | 5.1 |
| Avail. P (mg kg−1) | 14 | 31 |
| Active C (mg kg−1) | 683.6 | 1229.0 |
| OM (g kg−1) | 20.6 | 80.6 |
| K+ (meq 100 g−1) | 0.22 | 0.146 |
| Ca+2 (meq 100 g−1) | 2.31 | 6.29 |
| Mg+2 (meq 100 g−1) | 0.552 | 0.199 |
| pH | 5.64 | 5.89 |
| Grasses | Legumes a | Grasses (No. Seeds Pot−1) | Legumes (No. Seeds Pot−1) | Quilcas (N) | Castillapata (N) |
|---|---|---|---|---|---|
| OATS | VETCH | 1 | 2 | 4 | 4 |
| RYEG | VETCH | 2 | 2 | 4 | 4 |
| FELO | VETCH | 3 | 2 | 4 | 4 |
| BROM | VETCH | 3 | 2 | 4 | 3 |
| ORCH | VETCH | 3 | 2 | 3 | 4 |
| OATS | REDC | 1 | 2 | 4 | 3 |
| RYEG | REDC | 2 | 2 | 3 | 3 |
| FELO | REDC | 3 | 2 | 3 | 3 |
| BROM | REDC | 3 | 2 | 4 | 3 |
| ORCH | REDC | 3 | 2 | 3 | 4 |
| OATS | MELU | 1 | 2 | 1 | 4 |
| RYEG | MELU | 2 | 2 | 2 | 3 |
| FELO | MELU | 3 | 2 | 3 | 4 |
| BROM | MELU | 3 | 2 | 3 | 3 |
| ORCH | MELU | 3 | 2 | 3 | 4 |
| OATS | ALFA | 1 | 3 | 4 | 4 |
| RYEG | ALFA | 2 | 3 | 4 | 3 |
| FELO | ALFA | 3 | 3 | 3 | 3 |
| BROM | ALFA | 3 | 3 | 4 | 4 |
| ORCH | ALFA | 3 | 3 | 2 | 4 |
| OATS | 2 | 4 | 4 | ||
| RYEG | 4 | 4 | 4 | ||
| FELO | 6 | 4 | 4 | ||
| BROM | 6 | 3 | 4 | ||
| ORCH | 6 | 4 | 4 | ||
| VETCH | 4 | 4 | 4 | ||
| REDC | 4 | 4 | 4 | ||
| MELU | 4 | 4 | 4 | ||
| ALFA | 6 | 4 | 4 |
| Treatments a | Total Biomass (g Pot−1) | Total N (g Pot−1) | ||||||
|---|---|---|---|---|---|---|---|---|
| Quilcas | Castillapata | Quilcas | Castillapata | |||||
| Mean | SE | Mean | SE | Mean | SE | Mean | SE | |
| OATS/VETCH | 28.33 | 3.80 | 13.01 | 1.79 | 0.40 | 0.12 | 0.22 | 0.01 |
| RYEG/VETCH | 15.58 | 1.41 | 13.45 | 3.22 | 0.16 | 0.01 | 0.20 | 0.04 |
| FELO/VETCH | 19.30 | 3.11 | 14.18 | 2.78 | 0.40 | 0.08 | 0.22 | 0.05 |
| BROM/VETCH | 14.42 | 1.33 | 7.36 | 3.88 | 0.38 | 0.02 | 0.17 | 0.08 |
| ORCH/VETCH | 17.15 | 1.22 | 8.81 | 1.14 | 0.32 | 0.07 | 0.23 | 0.03 |
| OATS/REDC | 29.76 | 2.47 | 8.02 | 3.45 | 0.25 | 0.02 | 0.09 | 0.03 |
| RYEG/REDC | 19.79 | 5.13 | 7.98 | 1.31 | 0.26 | 0.11 | 0.08 | 0.00 |
| FELO/REDC | 9.70 | 1.82 | 3.15 | 0.56 | 0.17 | 0.03 | 0.04 | 0.01 |
| BROM/REDC | 7.14 | 1.65 | 0.84 | 0.14 | 0.11 | 0.03 | 0.02 | 0.00 |
| ORCH/REDC | 7.06 | 2.90 | 1.96 | 0.79 | 0.11 | 0.03 | 0.03 | 0.01 |
| OATS/MELU | 21.14 | N/A | 6.14 | 2.05 | 0.15 | N/A | 0.05 | 0.01 |
| RYEG/MELU | 15.17 | 3.38 | 4.26 | 0.75 | 0.16 | 0.05 | 0.05 | 0.01 |
| FELO/MELU | 10.74 | 2.08 | 5.79 | 1.66 | 0.13 | 0.02 | 0.06 | 0.01 |
| BROM/MELU | 7.61 | 2.03 | 1.87 | 0.23 | 0.11 | 0.02 | 0.03 | 0.01 |
| ORCH/MELU | 8.94 | 1.49 | 1.91 | 0.19 | 0.13 | 0.01 | 0.03 | 0.01 |
| OATS/ALFA | 25.09 | 7.90 | 8.25 | 1.15 | 0.24 | 0.06 | 0.06 | 0.01 |
| RYEG/ALFA | 15.98 | 3.90 | 4.94 | 1.01 | 0.17 | 0.03 | 0.05 | 0.01 |
| FELO/ALFA | 9.01 | 4.33 | 5.80 | 1.14 | 0.16 | 0.08 | 0.05 | 0.01 |
| BROM/ALFA | 6.19 | 0.83 | 1.11 | 0.39 | 0.15 | 0.06 | 0.02 | 0.01 |
| ORCH/ALFA | 5.61 | 4.25 | 1.70 | 0.25 | 0.07 | 0.05 | 0.02 | 0.00 |
| OATS | 18.07 | 4.29 | 6.39 | 1.64 | 0.21 | 0.04 | 0.06 | 0.01 |
| RYEG | 15.09 | 2.93 | 6.13 | 2.18 | 0.15 | 0.03 | 0.06 | 0.02 |
| FELO | 14.90 | 3.91 | 3.81 | 0.98 | 0.35 | 0.14 | 0.03 | 0.01 |
| BROM | 9.41 | 1.09 | 1.14 | 0.41 | 0.13 | 0.02 | 0.02 | 0.01 |
| ORCH | 7.32 | 1.33 | 2.18 | 0.66 | 0.13 | 0.03 | 0.03 | 0.01 |
| VETCH | 10.54 | 3.22 | 9.47 | 0.52 | 0.32 | 0.10 | 0.29 | 0.02 |
| REDC | 2.97 | 0.89 | 1.22 | 0.50 | 0.10 | 0.03 | 0.04 | 0.02 |
| MELU | 0.92 | 0.53 | 0.56 | 0.33 | 0.03 | 0.01 | 0.02 | 0.01 |
| ALFA | 1.80 | 0.98 | 1.03 | 0.92 | 0.04 | 0.02 | 0.03 | 0.03 |
| All treatments b | ||||||||
| Treatments | <0.001 | <0.001 | ||||||
| Soil | <0.001 | <0.001 | ||||||
| Soil × Treatment | 0.103 | 0.191 | ||||||
| Contrast: | ||||||||
| Biculture vs. Monoculture | <0.001 | <0.001 | ||||||
| Perennialsc | ||||||||
| Treatments | <0.001 | <0.001 | ||||||
| Soil | <0.001 | <0.001 | ||||||
| Soil × Treatment | 0.264 | 0.356 | ||||||
| Contrast: | ||||||||
| Biculture vs. Monoculture | <0.001 | <0.001 | ||||||
| Treatments a | Root Dry Biomass (g Pot−1) | Shoot Dry Biomass (g Pot−1) | ||||||
|---|---|---|---|---|---|---|---|---|
| Quilcas | Castillapata | Quilcas | Castillapata | |||||
| Mean | SE | Mean | SE | Mean | SE | Mean | SE | |
| OATS/VETCH | 4.67 | 1.36 | 1.40 | 0.21 | 23.66 | 3.18 | 11.61 | 1.59 |
| RYEG/VETCH | 4.88 | 0.78 | 3.52 | 0.76 | 10.70 | 0.71 | 9.93 | 2.51 |
| FELO/VETCH | 3.05 | 0.84 | 2.79 | 0.58 | 16.24 | 3.54 | 11.39 | 2.89 |
| BROM/VETCH | 1.00 | 0.10 | 0.67 | 0.28 | 13.42 | 1.26 | 6.69 | 3.60 |
| ORCH/VETCH | 4.85 | 0.72 | 1.15 | 0.30 | 12.30 | 0.53 | 7.66 | 1.09 |
| OATS/REDC | 4.78 | 0.63 | 0.86 | 0.28 | 24.98 | 2.12 | 7.16 | 3.17 |
| RYEG/REDC | 7.07 | 1.34 | 2.25 | 0.28 | 12.72 | 3.80 | 5.73 | 1.36 |
| FELO/REDC | 3.53 | 0.55 | 1.10 | 0.20 | 6.17 | 1.27 | 2.05 | 0.42 |
| BROM/REDC | 1.40 | 0.37 | 0.22 | 0.03 | 5.74 | 1.33 | 0.62 | 0.12 |
| ORCH/REDC | 2.69 | 1.15 | 0.82 | 0.39 | 4.36 | 1.77 | 1.14 | 0.43 |
| OATS/MELU | 5.83 | N/A | 0.78 | 0.25 | 15.31 | N/A | 5.36 | 1.91 |
| RYEG/MELU | 4.62 | 0.23 | 1.52 | 0.32 | 10.55 | 3.15 | 2.74 | 0.48 |
| FELO/MELU | 2.38 | 0.58 | 1.64 | 0.46 | 8.36 | 1.98 | 4.14 | 1.21 |
| BROM/MELU | 1.18 | 0.33 | 0.30 | 0.09 | 6.43 | 1.71 | 1.57 | 0.15 |
| ORCH/MELU | 3.18 | 1.06 | 0.66 | 0.05 | 5.76 | 0.44 | 1.24 | 0.17 |
| OATS/ALFA | 3.55 | 1.20 | 1.04 | 0.25 | 21.54 | 6.72 | 7.21 | 0.92 |
| RYEG/ALFA | 4.61 | 1.17 | 1.86 | 0.44 | 11.37 | 2.76 | 3.08 | 0.58 |
| FELO/ALFA | 3.08 | 1.53 | 2.12 | 0.10 | 5.93 | 2.82 | 3.68 | 1.06 |
| BROM/ALFA | 1.07 | 0.23 | 0.29 | 0.12 | 5.11 | 0.64 | 0.82 | 0.28 |
| ORCH/ALFA | 1.78 | 1.33 | 0.76 | 0.12 | 3.83 | 2.92 | 0.95 | 0.14 |
| OATS | 2.29 | 0.68 | 0.74 | 0.28 | 15.79 | 3.63 | 5.65 | 1.36 |
| RYEG | 6.08 | 1.36 | 2.24 | 0.82 | 9.01 | 1.58 | 3.89 | 1.36 |
| FELO | 4.80 | 1.13 | 1.48 | 0.39 | 10.1 | 2.87 | 2.33 | 0.68 |
| BROM | 1.35 | 0.18 | 0.22 | 0.07 | 8.06 | 1.27 | 0.92 | 0.35 |
| ORCH | 2.88 | 0.64 | 0.85 | 0.33 | 4.44 | 0.83 | 1.33 | 0.35 |
| VETCH | 0.74 | 0.11 | 0.83 | 0.20 | 9.80 | 3.13 | 8.63 | 0.44 |
| REDC | 0.95 | 0.30 | 0.50 | 0.22 | 2.02 | 0.64 | 0.72 | 0.28 |
| MELU | 0.20 | 0.08 | 0.08 | 0.03 | 0.73 | 0.46 | 0.49 | 0.3 |
| ALFA | 0.64 | 0.39 | 0.14 | 0.11 | 1.17 | 0.59 | 0.89 | 0.81 |
| All treatments b | ||||||||
| Treatments | <0.001 | <0.001 | ||||||
| Soil | <0.001 | <0.001 | ||||||
| Soil × Treatment | 0.018 | 0.120 | ||||||
| Contrast: | ||||||||
| Biculture vs. monoculture | <0.001 | <0.001 | ||||||
| Perennials c | ||||||||
| Treatments | <0.001 | <0.0001 | ||||||
| Soil | <0.001 | <0.0001 | ||||||
| Soil × Treatment | 0.353 | 0.162 | ||||||
| Contrast: | ||||||||
| Biculture vs. monoculture | <0.001 | <0001 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meza, K.; Vanek, S.J.; Sueldo, Y.; Olivera, E.; Ccanto, R.; Scurrah, M.; Fonte, S.J. Grass–Legume Mixtures Show Potential to Increase Above- and Belowground Biomass Production for Andean Forage-Based Fallows. Agronomy 2022, 12, 142. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010142
Meza K, Vanek SJ, Sueldo Y, Olivera E, Ccanto R, Scurrah M, Fonte SJ. Grass–Legume Mixtures Show Potential to Increase Above- and Belowground Biomass Production for Andean Forage-Based Fallows. Agronomy. 2022; 12(1):142. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010142
Chicago/Turabian StyleMeza, Katherin, Steven J. Vanek, Yulissa Sueldo, Edgar Olivera, Raúl Ccanto, Maria Scurrah, and Steven J. Fonte. 2022. "Grass–Legume Mixtures Show Potential to Increase Above- and Belowground Biomass Production for Andean Forage-Based Fallows" Agronomy 12, no. 1: 142. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010142