
The Response of Vegetable Sweet Potato (Ipomoea batatas Lam) Nodes to Different Concentrations of Encapsulation Agent and MS Salts

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sterilization and Establishment of Axenic Plantlets
2.2. Sub-Culturing of Plantlets
2.3. Preparation of Sodium Alginate and CaCl2
2.4. Encapsulation of Sweet Potato Nodal Segments with Sodium Alginate and CaCl2
2.5. Growth Media and Plantlet Retrieval from the Beads
2.6. Evaluation of MS Salts Strength on the Conversion of Sweet Potato Plantlets
2.7. Plantlets Growth Evaluation
2.8. Acclimatization and Transplanting
2.9. Statistical Analysis
3. Results
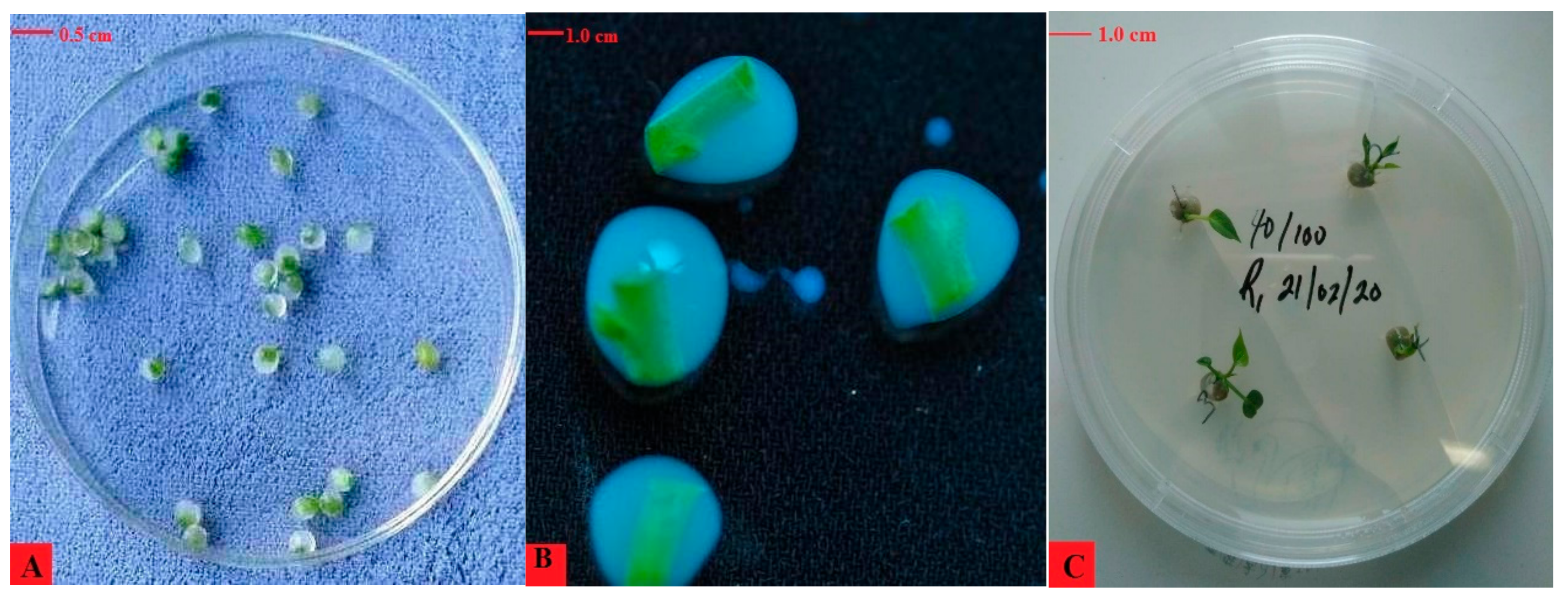
3.1. Effect of Sodium Alginate and CaCl2 Concentration on the Encapsulation of Vegetable Sweet Potato Nodes
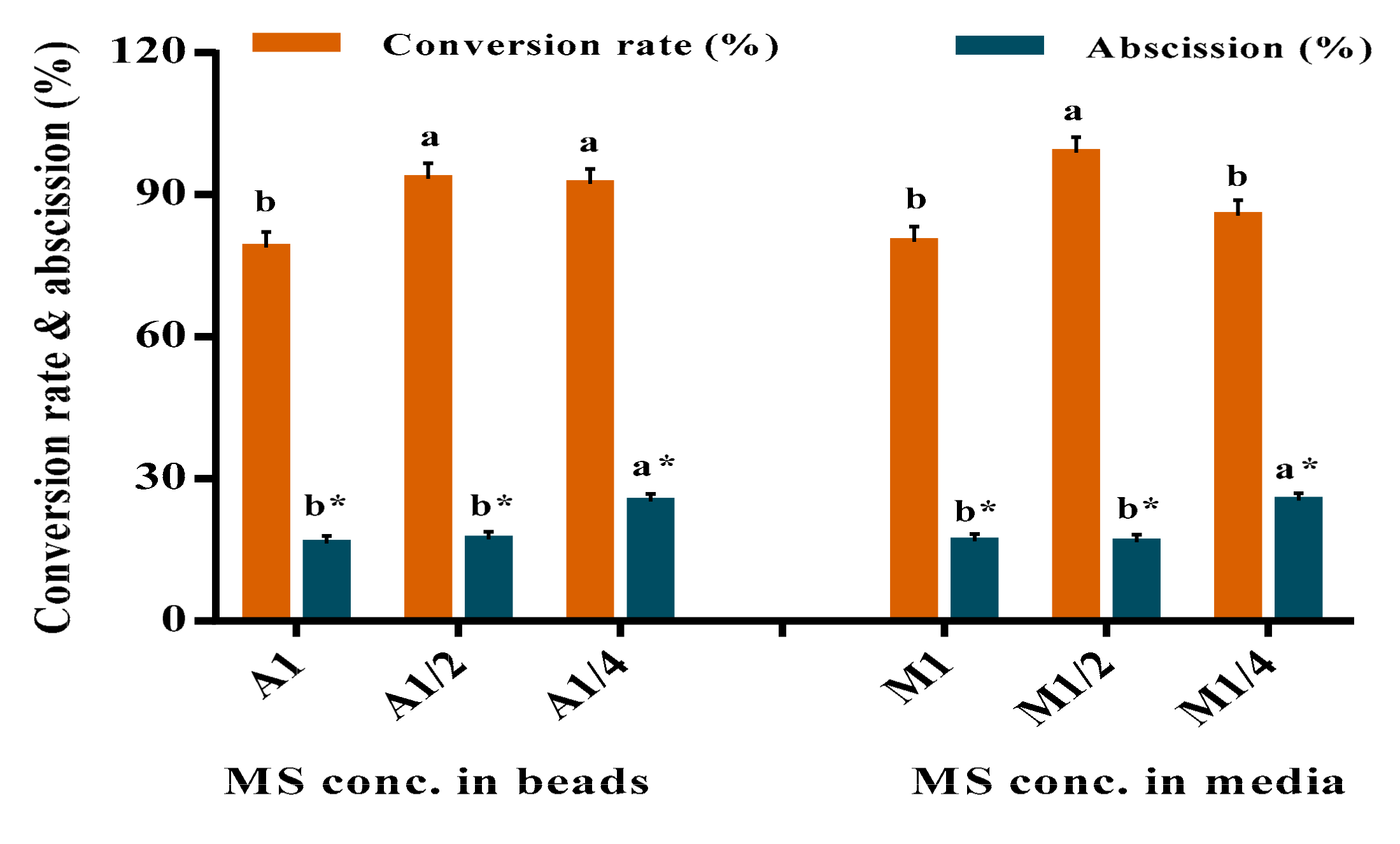
3.2. Effect of MS Salts Strength on the Conversion of Sweet Potato Plantlets
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murashige, T. The impact of plant tissue culture on agriculture. In Proceedings of the 4th International Congress of Plant Tissue and Cell Culture, University of Calgary, Calgary, AB, Canada, 20–25 August 1978; pp. 15–26. [Google Scholar]
- Rai, M.K.; Jaiswal, V.S.; Jaiswal, U. Encapsulation of shoot tips of guava (Psidium guajava L.) for short-term storage and germplasm exchange. Sci. Hortic. 2008, 118, 33–38. [Google Scholar] [CrossRef]
- Naik, S.K.; Chand, P.K. Nutrient-alginate encapsulation of in vitro nodal segments of pomegranate (Punica granatum L.) for germplasm distribution and exchange. Sci. Hortic. 2006, 108, 247–252. [Google Scholar] [CrossRef]
- Pattnaik, S.; Chand, P.K. Morphogenic response of the alginate-encapsulated axillary buds from in vitro shoot cultures of six mulberries. Plant Cell Tissue Organ Cult. 2000, 60, 177–185. [Google Scholar] [CrossRef]
- Piccioni, E.; Standardi, A. Encapsulation of micropropagated buds of six woody species. Plant Cell Tissue Organ Cult. 1995, 42, 221–226. [Google Scholar] [CrossRef]
- Brischia, R.; Piccioni, E.; Standardi, A. Micropropagation and synthetic seed in M.26 apple rootstock (II): A new protocol for production of encapsulated differentiating propagules. Plant Cell Tissue Organ Cult. 2002, 68, 137–141. [Google Scholar] [CrossRef]
- Attree, S.M.; Pomeroy, M.K.; Fowke, L.C. Production of vigorous, desiccation tolerant white spruce (Picea glauca [Moench.] Voss.) synthetic seeds in a bioreactor. Plant Cell Rep. 1994, 13, 601–606. [Google Scholar] [CrossRef]
- Ganapathi, T.R.; Srinivas, L.; Suprasanna, P.; Bapat, V.A. Regeneration of plants from alginate-encapsulated somatic embryos of banana cv. Rasthali (Musa SPP. AAB Group). In Vitro Cell. Dev. Biol. Plant 2001, 37, 178–181. [Google Scholar] [CrossRef]
- Gantait, S.; Kundu, S. Artificial seed technology for storage and exchange of plant genetic resources. In Advanced Technologies for Crop Improvement and Agricultural Productivity; Agrobios (International): Jodhpur, India, 2017; pp. 135–159. [Google Scholar]
- Bhattacharyya, P.; Kumar, V.; Van Staden, J. In vitro encapsulation based short term storage and assessment of genetic homogeneity in regenerated Ansellia africana (Leopard orchid) using gene targeted molecular markers. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 133, 299–310. [Google Scholar] [CrossRef]
- Zhang, P.; Chen, C.; Shen, Y.; Ding, T.; Ma, D.; Hua, Z.; Sun, D. Starch saccharification and fermentation of uncooked sweet potato roots for fuel ethanol production. Bioresour. Technol. 2013, 128, 835–838. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Shi, Y.; Xia, X.; Li, D.; Chen, Q. Life-cycle energy efficiency and environmental impacts of bioethanol production from sweet potato. Bioresour. Technol. 2013, 133, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Schafleitner, R.; Tincopa, L.R.; Palomino, O.; Rossel, G.; Robles, R.F.; Alagon, R.; Rivera, C.; Quispe, C.; Rojas, L.; Pacheco, J.A.; et al. A sweetpotato gene index established by de novo assembly of pyrosequencing and Sanger sequences and mining for gene-based microsatellite markers. BMC Genom. 2010, 11, 604. [Google Scholar] [CrossRef] [Green Version]
- Ara, H.; Jaiswal, U.; Jaiswal, V.S. Germination and plantlet regeneration from encapsulated somatic embryos of mango (Mangifera indica L.). Plant Cell Rep. 1999, 19, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Ara, H.; Jaiswal, U.; Jaiswal, V. Synthetic seed: Prospects and limitations. Curr. Sci. 2000, 78, 1438–1444. [Google Scholar]
- West, T.P.; Ravindra, M.B.; Preece, J.E. Encapsulation, cold storage, and growth of Hibiscus moscheutos nodal segments. Plant Cell Tissue Organ Cult. 2006, 87, 223–231. [Google Scholar] [CrossRef]
- Siddique, I.; Bukhari, N.A.W. Synthetic seed production by encapsulating nodal segment of Capparis decidua (Forsk.), in vitro regrowth of plantlets and their physio biochemical studies. Agrofor. Syst. 2018, 92, 1711–1719. [Google Scholar] [CrossRef]
- Patel, A.V.; Pusch, I.; Mix-Wagner, G.; Vorlop, K.D. A novel encapsulation technique for the production of artificial seeds. Plant Cell Rep. 2000, 19, 868–874. [Google Scholar] [CrossRef]
- Rihan, H.Z.; Kareem, F.; El-Mahrouk, M.E.; Fuller, M.P. Artificial seeds (principle, aspects and applications). Agronomy 2017, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Lata, H.; Chandra, S.; Khan, I.A.; Elsohly, M.A. Propagation through alginate encapsulation of axillary buds of Cannabis sativa L.—An important medicinal plant. Physiol. Mol. Biol. Plants 2009, 15, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.C.; Valkonen, J.P. Elimination of two viruses which interact synergistically from sweetpotato by shoot tip culture and cryotherapy. J. Virol. Methods 2008, 154, 135–145. [Google Scholar] [CrossRef]
- Gaba, V.; Singer, S. Propagation of sweetpotatoes, in situ germplasm conservation and conservation by tissue culture. In The Sweetpotato; Loebenstein, G., Thottappilly, G., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 65–80. [Google Scholar]
- Cantliffe, D.J.; Liu, J.R.; Schultheis, J.R. Development of artificial seeds of sweet potato for clonal propagation through somatic embryogenesis. In Methane from Bioass—A System Approach; Smith, W., Frank, J., Eds.; Elsevier: New York, NY, USA, 1987; pp. 183–195. [Google Scholar]
- Kikowska, M.; Thiem, B. Alginate-encapsulated shoot tips and nodal segments in mictopropagation of medicinal plands. A review. Herba Pol. 2011, 57, 45–57. [Google Scholar]
- Gray, D.J.; Purohit, A.; Triglano, R.N. Somatic embryogenesis and development of synthetic seed technology. Crit. Rev. Plant Sci. 1991, 10, 33–61. [Google Scholar] [CrossRef]
- Danso, K.E.; Ford-Lloyd, B.V. Encapsulation of nodal cuttings and shoot tips for storage and exchange of cassava germplasm. Plant Cell Rep. 2003, 21, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Dong, N.; Zhou, Y.; Ye, W.; Yu, S. In vitro regeneration protocol for synthetic seed production in upland cotton (Gossypium hirsutum L.). Plant Cell Tissue Organ Cult. (PCTOC) 2015, 123, 673–679. [Google Scholar] [CrossRef]
- Gantait, S.; Kundu, S.; Ali, N.; Sahu, N.C. Synthetic seed production of medicinal plants: A review on influence of explants, encapsulation agent and matrix. Acta Physiol. Plant. 2015, 37, 98. [Google Scholar] [CrossRef]
- Hegde, V.; Makeshkumar, T.; Sheela, M.; Visalakshi Chandra, C.; Koundinya, A.; Anil, S. Production of synthetic seed in cassava (Manihot esculenta Crantz). J. Root Crops 2016, 42, 5–9. [Google Scholar]
- Gray, D.J. Synthetic seed for clonal production of crop plants. In Recent Advances in the Development and Germination of Seeds; Taylorson, R.B., Ed.; NATO ASI Series (Series A: Life Sciences); Springer: Boston, MA, USA, 1989; Volume 187, pp. 29–45. [Google Scholar]
- Kamińska, M.; Gołębiewski, M.; Tretyn, A.; Trejgell, A. Efficient long-term conservation of Taraxacum pieninicum synthetic seeds in slow growth conditions. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 132, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Wilms, H.; Fanega Sleziak, N.; Van der Auweraer, M.; Brands, M.; Verleije, M.; Hardeman, D.; Andre, E.; Panis, B. Development of a fast and user-friendly cryopreservation protocol for sweet potato genetic resources. Sci. Rep. 2020, 10, 14674. [Google Scholar] [CrossRef]
- Parveen, S.; Shahzad, A. Encapsulation of nodal segments of Cassia angustifolia Vahl. for short-term storage and germplasm exchange. Acta Physiol. Plant. 2014, 36, 635–640. [Google Scholar] [CrossRef]
- Arumugam, G.; Sinniah, U.R.; Swamy, M.K.; Lynch, P.T. Encapsulation of in vitro Plectranthus amboinicus (Lour.) Spreng. shoot apices for propagation and conservation. 3 Biotech 2019, 9, 298. [Google Scholar] [CrossRef]
- Ubalua, A.O.; Okoroafor, U.E. Micropropagation and postflask management of sweet potato using locally available materials as substrates for hardening. Plant Knowl. J. 2013, 2, 56–61. [Google Scholar]
- Cantliffe, D.J. Advanced propagation systems for biomass species: A model system based on sweet potato. Biomass Bioenergy 1993, 5, 63–69. [Google Scholar] [CrossRef]
- Cheée, R.P.; Cantliffe, D.J. Improved Production Procedures for Somatic Embryos of Sweetpotato for a Synthetic Seed System. HortScience 1992, 27, 1314–1316. [Google Scholar] [CrossRef] [Green Version]
- Blakesley, D.; Al-Mazrooei, S.; Henshaw, G.G. Cryopreservation of embryogenic tissue of sweet potato (Ipomoea batatas): Use of sucrose and dehydration for cryoprotection. Plant Cell Rep. 1995, 15, 259–263. [Google Scholar] [CrossRef]
- Namanda, S.; Gibson, R.; Sindi, K. Sweetpotato Seed Systems in Uganda, Tanzania, and Rwanda. J. Sustain. Agric. 2011, 35, 870–884. [Google Scholar] [CrossRef]
- Vettorazzi, R.G.; Carvalho, V.S.; Sudré, C.P.; Rodrigues, R. Developing an in vitro optimized protocol to sweet potato landraces conservation. Acta Sci. Agron. 2017, 39, 359–367. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.; Drew, R. Current applications of tissue culture in plant propagation and improvement. Functional Plant Biol. 1990, 17, 267–289. [Google Scholar] [CrossRef]
- Torres, A.C.; Ze, N.M.E.; Cantliffe, D.J. Abscisic acid and osmotic induction of synchronous somatic embryo development of sweet potato. In Vitro Cell. Dev. Biol.-Plant 2001, 37, 262–267. [Google Scholar] [CrossRef]
- Chée, R.P.; Cantliffe, D.J. Selective enhancement of Ipomoea batatas Poir. embryogenic and non-embryogenic callus growth and production of embryos in liquid culture. Plant Cell Tissue Organ Cult. 1988, 15, 149–159. [Google Scholar] [CrossRef]
- DeWald, S.; Cantliffe, D. Histology of callus initiation and somatic embryogenesis from sweet potato (Ipomoea batatas Poir) shoot apicies. HortScience 1988, 23, 812. [Google Scholar]
- Luan, Y.-S.; Zhang, J.; Gao, X.-R.; An, L.-J. Mutation induced by ethylmethanesulphonate (EMS), in vitro screening for salt tolerance and plant regeneration of sweet potato (Ipomoea batatas L.). Plant Cell Tissue Organ Cult. 2007, 88, 77–81. [Google Scholar] [CrossRef]
- Manrique-Trujillo, S.; Díaz, D.; Reaño, R.; Ghislain, M.; Kreuze, J. Sweetpotato plant regeneration via an improved somatic embryogenesis protocol. Sci. Hortic. 2013, 161, 95–100. [Google Scholar] [CrossRef]
- Masekesa, T.R.; Gasura, E.; Ngadze, E.; Icishahayo, D.; Kujeke, G.T.; Chidzwondo, F.; Robertson, I. Efficacy of Zeatin, Kinetin and Thidiazuron in induction of adventitious root and shoot from petiole explants of sweetpotato cv. Brondal. S. Afr. J. Bot. 2016, 104, 1–5. [Google Scholar] [CrossRef]
- Villordon, A.Q.; LaBonte, D.R. Genetic Variation among Sweetpotatoes Propagated through Nodal and Adventitious Sprouts. J. Am. Soc. Hortic. Sci. Jashs 1996, 121, 170–174. [Google Scholar] [CrossRef] [Green Version]
- Rao, P.S.; Suprasanna, P.; Ganapathi, T.R.; Bapat, V.A. Synthetic seeds: Concepts, methods and application. In Plant Tissue Culture and Molecular Biology; Srivastava, P.V., Ed.; Narosa: New Delhi, India, 1998; pp. 607–619. [Google Scholar]
- Singh, A.K.; Varshney, R.; Sharma, M.; Agarwal, S.S.; Bansal, K.C. Regeneration of plants from alginate-encapsulated shoot tips of Withania somnifera (L.) Dunal, a medicinally important plant species. J. Plant Physiol. 2006, 163, 220–223. [Google Scholar] [CrossRef]
- Kumar, S.; Rai, M.K.; Singh, N.; Mangal, M. Alginate-encapsulation of shoot tips of jojoba [Simmondsia chinensis (Link) Schneider] for germplasm exchange and distribution. Physiol. Mol. Biol. Plants 2010, 16, 379–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micheli, M.; Hafiz, I.A.; Standardi, A. Encapsulation of in vitro-derived explants of olive (Olea europaea L. cv. Moraiolo): II. Effects of storage on capsule and derived shoots performance. Sci. Hortic. 2007, 113, 286–292. [Google Scholar] [CrossRef]
- Namanda, S.; Gatimu, R.; Agili, S.; Khisa66, S.; Ndyetabula, I.; Bagambisa, C. Micropropagation and Hardening Sweet potato Tissue Culture Plantlets. In A Manual Developed from the SASHA Project’s Experience in Tanzania; International Potato Center (CIP): Lima, Peru, 2015. [Google Scholar]
- Guri, A.Z.; Patel, K.N. Compositions and Methods to Prevent Microbial Contamination of Plant Tissue Culture Media. U.S. Patent No. 5,750,402, 12 May 1998. [Google Scholar]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Sakamoto, M.; Suzuki, T. Effect of Nutrient Solution Concentration on the Growth of Hydroponic Sweetpotato. Agronomy 2020, 10, 1708. [Google Scholar] [CrossRef]
- SAS. SAS Certification Prep Guide: Base Programming for SAS 9; SAS Institute: Cary, NC, USA, 2008. [Google Scholar]
- Motulsky, H. Prism 4 Statistics Guide—Statistical Analyses for Laboratory and Clinical Researchers.; GraphPad Software Inc.: San Diego, CA, USA, 2003; pp. 122–126. [Google Scholar]
- Loebenstein, G. Chapter Two-Control of Sweet Potato Virus Diseases. In Advances in Virus Research; Loebenstein, G., Katis, N.I., Eds.; Academic Press: Cambridge, MA, USA, 2015; Volume 91, pp. 33–45. [Google Scholar]
- Rihan, H.Z.; Al-Issawi, M.; Burchett, S.; Fuller, M.P. Artificial Seed Production from Encapsulated Microshoots of Cauliflower (Brassica oleraceae var. Botrytis). In VII International Symposium on In Vitro Culture and Horticultural Breeding; ISHS: Leuven, Belgium, 2012; pp. 419–425. [Google Scholar]
- Hatzilazarou, S.; Kostas, S.; Economou, A.S. Plant regeneration of Nerium oleander L. from alginate-encapsulated shoot explants after short-term cold storage. J. Hortic. Sci. Biotechnol. 2019, 94, 441–447. [Google Scholar] [CrossRef]
- Hatzilazarou, S.; Kostas, S.; Nendou, T.; Economou, A. Conservation, Regeneration and Genetic Stability of Regenerants from Alginate-Encapsulated Shoot Explants of Gardenia jasminoides Ellis. Polymers 2021, 13, 1666. [Google Scholar] [CrossRef]
- Ahmed, M.R.; Anis, M.; Al-Etta, H.A. Encapsulation technology for short-term storage and germplasm exchange of Vitex trifolia L. Rend. Lincei 2015, 26, 133–139. [Google Scholar] [CrossRef]
- Rathore, M.S.; Kheni, J. Alginate encapsulation and in vitro plantlet regeneration in critically endangered medicinal plant, Withania coagulans (Stocks) Dunal. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 129–134. [Google Scholar] [CrossRef]
- Maruyama, E.; Kinoshita, I.; Ishii, K.; Shigenaga, H.; Ohba, K.; Saito, A. Alginate-encapsulated technology for the propagation of the tropical forest trees: Cedrela odorata L., Guazuma crinita MART., and Jacaranda mimosaefolia D. DON. Silvae Genet. (Ger.) 1997, 46, 17–23. [Google Scholar]
- Adriani, M.; Piccioni, E.; Standardi, A. Effect of different treatments on the conversion of ‘Hayward’ kiwifruit synthetic seeds to whole plants following encapsulation of in vitro-derived buds. N. Z. J. Crop Hortic. Sci. 2000, 28, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Benelli, C. Encapsulation of Shoot Tips and Nodal Segments for in Vitro Storage of “Kober 5BB” Grapevine Rootstock. Horticulturae 2016, 2, 10. [Google Scholar] [CrossRef]
- Akhtar, R.; Shahzad, A. Alginate encapsulation in Glycyrrhiza glabra L. with phyto-chemical profiling of root extracts of in vitro converted plants using GC-MS analysis. Asian Pac. J. Trop. Biomed. 2017, 7, 855–861. [Google Scholar] [CrossRef]
- Shilpha, J.; Pandian, S.; Largia, M.J.V.; Sohn, S.I.; Ramesh, M. Short-term storage of Solanum trilobatum L. synthetic seeds and evaluation of genetic homogeneity using SCoT markers. Plant Biotechnol. Rep. 2021, 15, 651–661. [Google Scholar] [CrossRef]
- Saha, S.; Sengupta, C.; Ghosh, P. Encapsulation, short-term storage, conservation and molecular analysis to assess genetic stability in alginate-encapsulated microshoots of Ocimum kilimandscharicum Guerke. Plant Cell Tissue Organ Cult. (PCTOC) 2015, 120, 519–530. [Google Scholar] [CrossRef]
- Bapat, V.A.; Mhatre, M. Bioencapsulation of Somatic Embryos in Woody Plants. In Protocol for Somatic Embryogenesis in Woody Plants; Jain, S.M., Gupta, P.K., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 77, pp. 539–552. [Google Scholar]
- Rezali, N.I.; Jaafar Sidik, N.; Saleh, A.; Osman, N.I.; Mohd Adam, N.A. The effects of different strength of MS media in solid and liquid media on in vitro growth of Typhonium flagelliforme. Asian Pac. J. Trop. Biomed. 2017, 7, 151–156. [Google Scholar] [CrossRef]
- Wan Nurul Hidayah, W.; Norrizah, J.; Sharifah Aminah, S.; Sharipah Ruzaina, S.; Faezah, P. Effect of medium strength and hormones concentration on regeneration of Pogostemon cablin using nodes explant. Asian J. Biotechnol. 2012, 4, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Bukhari, N.A.W.; Siddique, I.; Perveen, K.; Siddiqui, I.; Alwahibi, M.S. Synthetic Seed Production and Physio-Biochemical Studies in Cassia Angustifolia Vahl.—A Medicinal Plant. Acta Biol. Hung. 2014, 65, 355–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadel, D.; Kintzios, S.; Economou, S.A.; Moschopoulou, G.; Constantinidou, A.H.-I. Effect of Different Strength of Medium on Organogenesis, Phenolic Accumulation and Antioxidant Activity of Spearmint (Mentha spicata L.). Open Hortic. J. 2010, 3, 31–35. [Google Scholar] [CrossRef]
- Bouwkamp, J.C. Sweet Potato Products: A Natural Resource for the Tropics, 1st ed.; CRC Press: Boca Raton, FL, USA, 1985. [Google Scholar]






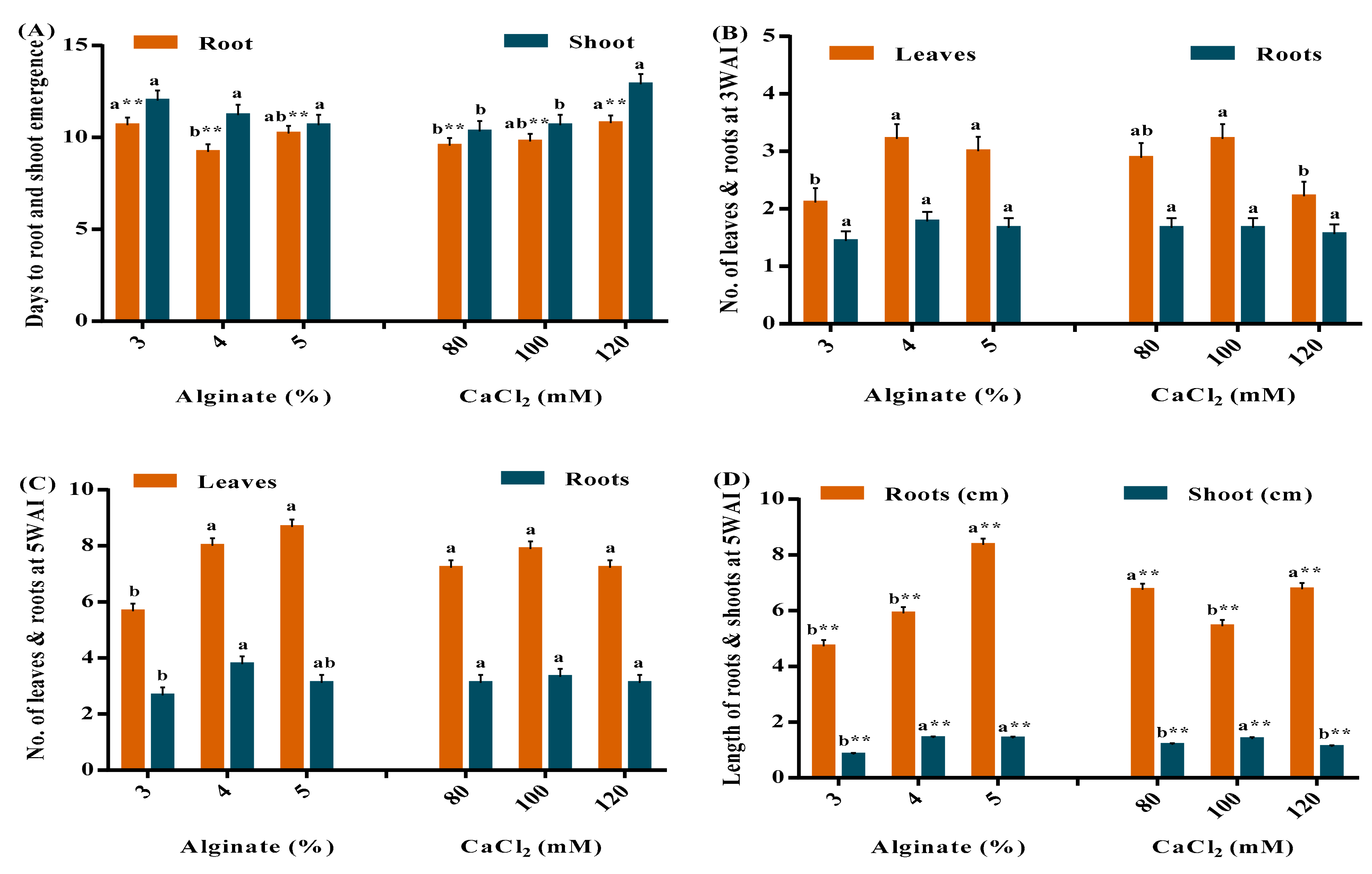{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days to Root Emergence | Root Length (cm) | Shoot Length (cm) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Treatments | 80 mM CaCl2 | 100 mM CaCl2 | 120 mM CaCl2 | 80 mM CaCl2 | 100 mM CaCl2 | 120 mM CaCl2 | 80 mM CaCl2 | 100 mM CaCl2 | 120 mM CaCl2 |
| 3% Alginate | 10.0 ± 0.6 bc | 8.6 ± 0.3 c | 13.3 ± 0.3 a | 3.3 ± 0.3 e | 5.8 ± 0.3 cd | 5.0 ± 0.4 cd | 0.7 ± 0.1 c | 1.2 ± 0.0 b | 0.5 ± 0.0 c |
| 4% Alginate | 9.0 ± 0.6 c | 8.6 ± 0.9 c | 10 ± 1.2 bc | 6.1 ± 0.4 c | 5.6 ± 0.1 cd | 5.9 ± 1.0 cd | 1.2 ± 0.2 b | 1.6 ± 0.0 a | 1.4 ± 0.1 ab |
| 5% Alginate | 9.6 ± 0.3 c | 12.0 ± 1.2 ab | 9.0 ± 0.6 c | 10.8 ± 0.1 a | 4.9 ± 0.3 d | 9.3 ± 0.2 b | 1.6 ± 0.1 a | 1.2 ± 0.2 bc | 1.4 ± 0.1 ab |
| Number of Days to Shoot Emergence | Number of Roots at Two Weeks after Inoculation | |||||
|---|---|---|---|---|---|---|
| Treatments | MS Concentration in Encapsulated Nodes (A) g/L | |||||
| MS Conc. in Media (M) g/L | Full MS (A1) | ½ MS (A1/2) | ¼ MS (A1/4) | Full MS (A1) | ½ MS (A1/2) | ¼ MS (A1/4) |
| Full MS in media (M1) | 8.6 ± 0.9 a | 5.6 ± 0.3 bc | 5.0 ± 0.6 cd | 1.3 ± 0.3 b | 2.6 ± 0.3 a | 2.6 ± 0.3 a |
| ½ MS in media (M1/2) | 7.0 ± 1.2 ab | 5.6 ± 0.3 bc | 7.6 ± 0.3 a | 2.6 ± 0.3 a | 2.6 ± 0.3 a | 2.0 ± 0.0 ab |
| ¼ MS in media (M1/4) | 8.0 ± 0.6 a | 3.6 ± 0.3 d | 5.6 ± 0.7 bc | 1.6 ± 0.3 b | 2.0 ± 0.6 ab | 2.0 ± 0.0 ab |
| Number of Leaves | Length of Shoots (cm) | |||||
|---|---|---|---|---|---|---|
| Treatments | MS Concentration in Encapsulated Nodes (A) g/L | |||||
| MS Conc. in Media (M) g/L | Full MS (A1) | ½ MS (A1/2) | ¼ MS (A1/4) | Full MS (A1) | ½ MS (A1/2) | ¼ MS (A1/4) |
| Full MS in media (M1) | 7.0 ± 0.6 d | 9.3 ± 0.3 ab | 10.3 ± 0.8 a | 0.9 ± 0.1 c | 1.3 ± 0.3 bc | 1.4 ± 0.1 bc |
| ½ MS in media (M1/2) | 8.6 ± 0.7 bc | 10.6 ± 0.3 a | 9.3 ± 0.3 ab | 1.3 ± 0.1 bc | 2.6 ± 0.1 a | 2.4 ± 0.4 a |
| ¼ MS in media (M1/4) | 8.0 ± 0.6 bcd | 8.6 ± 0.3 bc | 7.3 ± 0.3 cd | 1.7 ± 0.2 b | 1.7 ± 0.3 b | 1.8 ± 0.1 b |
| Leaf Abscission (%) | |||
|---|---|---|---|
| Treatments | MS Concentration in encapsulated nodes(A) g/L | ||
| MS Conc. in Media (M) g/L | Full MS (A1) | ½ MS (A1/) | ¼ MS (A1/4) |
| Full MS in media (M1) | 17 ± 1.6 bc | 17 ± 3.2 bc | 17 ± 0.0 bc |
| ½ MS in media (M1/2) | 15 ± 1.2 c | 12 ± 1.1 c | 23 ± 1.9 b |
| ¼ MS in media (M1/4) | 17 ± 2.8 bc | 23 ± 5.3 b | 36 ± 4.1 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tadda, S.A.; Kui, X.; Yang, H.; Li, M.; Huang, Z.; Chen, X.; Qiu, D. The Response of Vegetable Sweet Potato (Ipomoea batatas Lam) Nodes to Different Concentrations of Encapsulation Agent and MS Salts. Agronomy 2022, 12, 19. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010019
Tadda SA, Kui X, Yang H, Li M, Huang Z, Chen X, Qiu D. The Response of Vegetable Sweet Potato (Ipomoea batatas Lam) Nodes to Different Concentrations of Encapsulation Agent and MS Salts. Agronomy. 2022; 12(1):19. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010019
Chicago/Turabian StyleTadda, Shehu A., Xiaohua Kui, Hongjuan Yang, Min Li, Zhehong Huang, Xuanyang Chen, and Dongliang Qiu. 2022. "The Response of Vegetable Sweet Potato (Ipomoea batatas Lam) Nodes to Different Concentrations of Encapsulation Agent and MS Salts" Agronomy 12, no. 1: 19. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010019