
Cause of Death: Phytophthora or Flood? Effects of Waterlogging on Phytophthora medicaginis and Resistance of Chickpea (Cicer arietinum)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant and Oomycete Material
2.2. Experiment 1: Effect of Early and Late Waterlogging on Phytophthora Root Rot in Chickpea
2.2.1. Design and Conditions
2.2.2. In Planta Waterlogging and Inoculation
2.3. Experiment 2: Detection of P. medicaginis DNA in Chickpea Root and Soil Media under Transient and Long-Term Waterlogging Using qPCR
2.3.1. Design and Conditions
2.3.2. In Planta Waterlogging and Inoculation
2.4. Experiment 3: Detection of P. medicaginis Zoospores and PRR in Chickpea under Hypoxic Hydroponic Conditions
2.4.1. Design and Conditions
2.4.2. In Planta Flooding & Infection
2.5. Statistical Analysis
3. Results
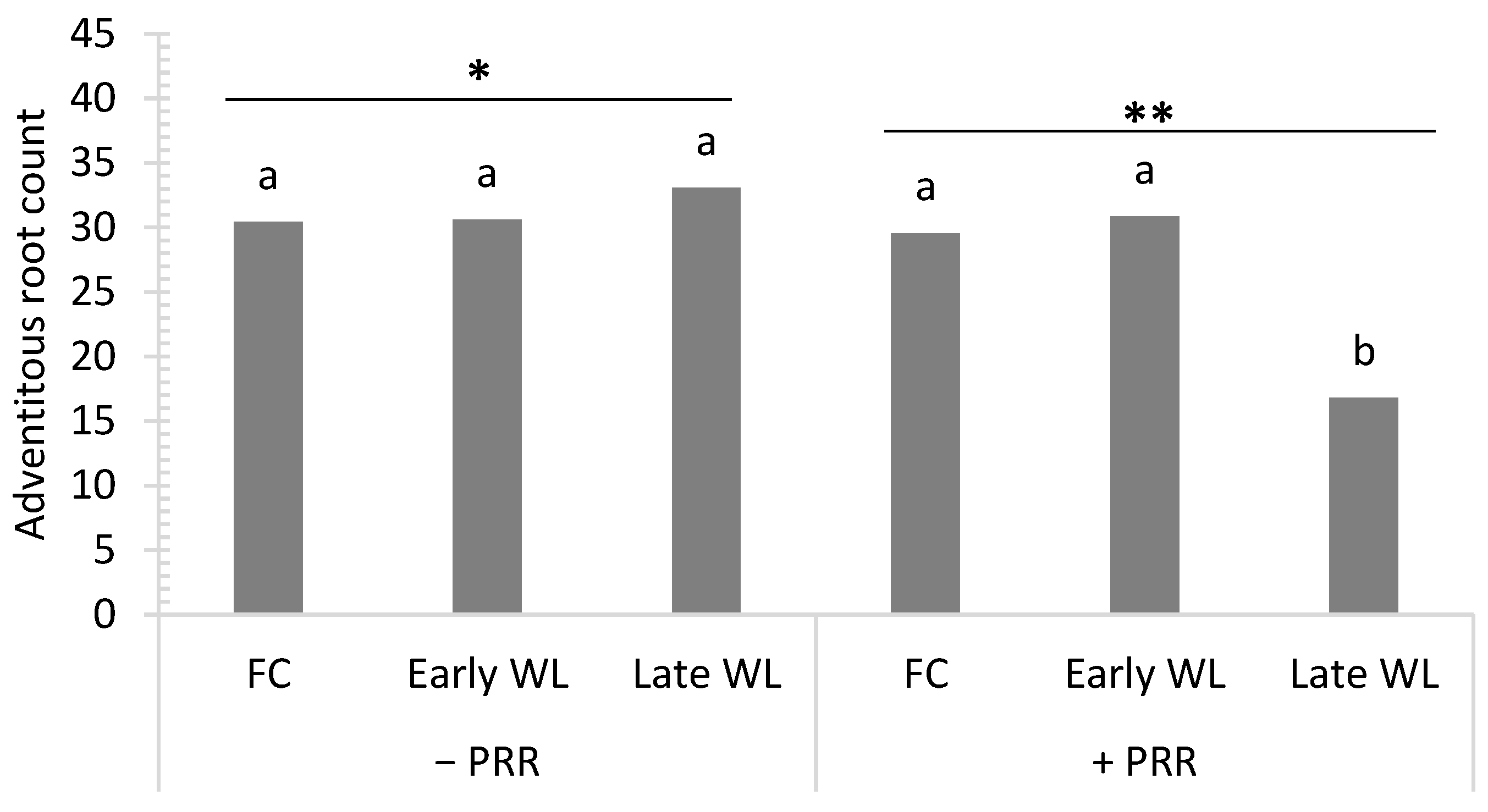
3.1. Experiment 1: Effect of Early and Late Growth Stage Waterlogging and Phytophthora Root Rot in Chickpea
3.2. Experiment 2: Detection of P. medicaginis DNA in Chickpea Root and Soil Media under Transient and Long-Term Waterlogging Using qPCR
3.3. Experiment 3: Detection of P. medicaginis Zoospores and PRR in Chickpea under Hypoxic Hydroponic Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Redden, R.; Berger, J. History and origin of chickpea. In Chickpea Breeding and Management; Cromwell Press: Wiltshire, UK, 2007; pp. 1–13. [Google Scholar]
- Rea, G.; Laurenzi, M.; Tranquilli, E.; D’Ovidio, R.; Federico, R.; Angelini, R. Developmentally and wound-regulated expression of the gene encoding a cell wall copper amine oxidase in chickpea seedlings. FEBS Lett. 1998, 437, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Rawal, V.; Navarro, D. The Global Economy of Pulses; FAO: Rome, Italy, 2019; p. 190. [Google Scholar]
- Rees, R.O.; Brouwer, J.B.; Mahoney, J.E.; Walton, G.H.; Brinsmead, R.B.; Knights, E.J.; Beech, D.F. Pea and chickpea production in Australia. In Expanding the Production and Use of Cool Season Food Legumes; Muehlbauer, F.J., Kaiser, W.J., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 412–425. [Google Scholar]
- Merga, B.; Haji, J. Economic importance of chickpea: Production, value, and world trade. Cogent Food Agric. 2019, 5, 1615718. [Google Scholar] [CrossRef]
- Hirabayashi, Y.; Mahendran, R.; Koirala, S.; Konoshima, L.; Yamazaki, D.; Watanabe, S.; Kim, H.; Kanae, S. Global flood risk under climate change. Nat. Clim. Change 2013, 3, 816–821. [Google Scholar] [CrossRef]
- Food and Agriculture Organization. The Impact of Natural Hazards and Disasters on Agriculture; UN: Rome, Italy, 2015. [Google Scholar]
- Bithell, S.; Moore, K.; McKay, A.; Harden, S.; Simpfendorfer, S. Phytophthora root rot of chickpea: Inoculum concentration and seasonally dependent success for qPCR based predictions of disease and yield loss. Australas. Plant Pathol. 2021, 50, 91–103. [Google Scholar] [CrossRef]
- Schwinghamer, M.; Southwell, R.; Moore, K.; Knights, E. Phytophthora root rot of chickpea. In Compendium of Chickpea and Lentil Diseases and Pests; The American Phytopathological Society: St Paul, MN, USA, 2011; pp. 22–25. [Google Scholar]
- Erwin, D.C.; Bartnicki-Garcia, S.; Tsao, P.H. (Eds.) Phytophthora: Its Biology, Taxonomy, Ecology, and Pathology; American Phytopathological Society: St. Paul, MN, USA, 1983; p. 392. [Google Scholar]
- Davison, E.; Tay, F. The effect of aeration on colony diameter, sporangium production and zoospore germination of Phytophthora cinnamomi. New Phytol. 1986, 103, 735–744. [Google Scholar] [CrossRef]
- Palta, J.A.; Ganjeali, A.; Turner, N.C.; Siddique, K.H.M. Effects of transient subsurface waterlogging on root growth, plant biomass and yield of chickpea. Agric. Water Manag. 2010, 97, 1469–1476. [Google Scholar] [CrossRef]
- Colmer, T.; Voesenek, L. Flooding tolerance: Suites of plant traits in variable environments. Funct. Plant Biol. 2009, 36, 665–681. [Google Scholar] [CrossRef]
- Dron, N.M.; Sutton, T.; Simpfendorfer, S.; Harden, S.; Hobson, K. Phenotyping for waterlogging tolerance as a proxy for Phytophthora medicaginis resistance in chickpea. Plant Health Prog. 2021, 22, 287–293. [Google Scholar] [CrossRef]
- Nguyen, V.; Vuong, T.; VanToai, T.; Lee, J.; Wu, X.; Mian, M.R.; Dorrance, A.; Shannon, J.; Nguyen, H. Mapping of quantitative trait loci associated with resistance to Phytophthora sojae and flooding tolerance in soybean. Crop Sci. 2012, 52, 2481–2493. [Google Scholar] [CrossRef]
- Knights, E.J.; Southwell, R.J.; Schwinghamer, M.W.; Harden, S. Resistance to Phytophthora medicaginis Hansen and Maxwell in wild Cicer species and its use in breeding root rot resistant chickpea (Cicer arietinum L.). Aust. J. Agric. Res. 2008, 59, 383–387. [Google Scholar] [CrossRef]
- Bakker, D.; Hamilton, G.; Houlbrooke, D.; Spann, C. The effect of raised beds on soil structure, waterlogging, and productivity on duplex soils in Western Australia. Soil Res. 2005, 43, 575–585. [Google Scholar] [CrossRef]
- Pagliai, M.; Vignozzi, N.; Pellegrini, S. Soil structure and the effect of management practices. Soil Tillage Res. 2004, 79, 131–143. [Google Scholar] [CrossRef]
- Croser, J.; Ahmad, F.; Clarke, H.; Siddique, K. Utilisation of wild Cicer in chickpea improvement—progress, constraints, and prospects. Aust. J. Agric. Res. 2003, 54, 429–444. [Google Scholar] [CrossRef]
- Chauhan, Y. Screening for tolerance to salinity and waterlogging: Case studies with pigeonpea and chickpea. In Proceedings of the Consultants’ Workshop: Adaptation of Chickpea and Pigeonpea to Abiotic Stresses, International Crops Research Institute for the Semi-Arid Tropics, Patancheru, India, 19–21 December 1984. [Google Scholar]
- Cowie, A.L.; Jessop, R.S.; MacLeod, D.A. A Study of Waterlogging Damage in Australian Chickpea Cultivars; International Chickpea and Pigeonpea Newsletter; International Crops Research Institute for the Semi-Arid Tropics: Patancheruvu, India, 1995. [Google Scholar]
- Brinsmead, R.B.; Rettke, M.L.; Irwin, J.A.G.; Ryley, M.J.; Langdon, P.W. Resistance in chickpea to Phytophthora-megasperma f sp. medicaginis. Plant Dis. 1985, 69, 504–506. [Google Scholar] [CrossRef]
- Manik, S.M.N.; Pengilley, G.; Dean, G.; Field, B.; Shabala, S.; Zhou, M. Soil and Crop Management Practices to Minimize the Impact of Waterlogging on Crop Productivity. Front. Plant Sci. 2019, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Amalraj, A.; Taylor, J.; Bithell, S.; Li, Y.; Moore, K.; Hobson, K.; Sutton, T. Mapping resistance to Phytophthora root rot identifies independent loci from cultivated (Cicer arietinum L.) and wild (Cicer echinospermum PH Davis) chickpea. Theor. Appl. Genet. 2019, 132, 1017–1033. [Google Scholar] [CrossRef]
- Schwenke, G.; Nachimuthu, G.; Mercer, C.; McPherson, A. Tail-drain sediments are a potential hotspot for nitrous oxide emissions in furrow-irrigated Vertisols used to grow cotton: A laboratory incubation study. J. Environ. Qual. 2020, 49, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Corbin, E.; Brockwell, J.; Gault, R. Nodulation studies on chickpea (Cicer arietinum). Aust. J. Exp. Agric. Anim. Husb. 1977, 17, 126–134. [Google Scholar] [CrossRef]
- Samineni, S.; Siddique, K.H.; Gaur, P.M.; Colmer, T.D. Salt sensitivity of the vegetative and reproductive stages in chickpea (Cicer arietinum L.): Podding is a particularly sensitive stage. Environ. Exp. Bot. 2011, 71, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Amalraj, A.; Taylor, J.; Sutton, T. A hydroponics based high throughput screening system for Phytophthora root rot resistance in chickpea (Cicer arietinum L.). Plant Methods 2019, 15, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, D.; Cullis, B.; Gilmour, A.; Thompson, R. Asreml: An R Package to Fit the Linear Mixed Model; VSNi: Hemel Hempstead, UK, 2018; Available online: https://www.vsni.co.uk/software/asreml (accessed on 16 February 2021).
- Nielsen, S.; Rogers, S.; Conway, A. Biometry Training: Functions to Assist Design and Analysis of Agronomic Experiments, R Package version 0.8.0; University of Adelaide Biometry Hub: Adelaide, Australia, 2019; Available online: https://biometryhub.github.io/BiometryTraining/ (accessed on 6 April 2021).
- Team, R.C. R: A Language and Environment for Statistical Computing; R Version 4.0. 3; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; American Phytopathological Society: St. Paul, MN, USA, 1996. [Google Scholar]
- Moore, K.J. Oxygen and Carbon Dioxide in the Ecology Phytophthora Species; University of Sydney: Sydney, Australia, 1975. [Google Scholar]
- Kuan, T.L.; Erwin, D.C. Predisposition effect of water saturation of soil on Phytophthora root rot of alfalfa. Phytopathology 1980, 70, 981–986. [Google Scholar] [CrossRef]
- Jacobs, K.A.; MacDonald, J.D.; Berry, A.M.; Costello, L.R. The effect of low oxygen stress on Phytophthora cinnamomi infection and disease of cork oak roots. In Pillsbury, Norman H.; Verner, Jared; Tietje, William D., Technical Coordinators. 1997. Proceedings of a Symposium on Oak Woodlands: Ecology, Management, and Urban Interface Issues; 19–22 March 1996; San Luis Obispo, CA. Gen. Tech. Rep. PSW-GTR-160; Pacific Southwest Research Station, Forest Service, U.S. Department of Agriculture: Albany, CA, USA, 1997; pp. 553–558. [Google Scholar]
- Curtis, D.S.; Zentmyer, G.A. Effect of oxygen supply on Phytophthora root rot of avocado in nutrient solution. Am. J. Bot. 1949, 36, 471–474. [Google Scholar] [CrossRef]
- Ponnamperuma, F.N. Effects of flooding on soils. In Flooding and Plant Growth; Academic Press: San Diego, CA, USA, 1984; pp. 10–42. [Google Scholar]
- Xu, L.; Pan, R.; Shabala, L.; Shabala, S.; Zhang, W.-Y. Temperature influences waterlogging stress-induced damage in Arabidopsis through the regulation of photosynthesis and hypoxia-related genes. Plant Growth Regul. 2019, 89, 143–152. [Google Scholar] [CrossRef]
- Cowie, A.L.; Jessop, R.S.; MacLeod, D.A. Effects of waterlogging on chickpeas I. Influence of timing of waterlogging. Plant Soil 1996, 183, 97–103. [Google Scholar] [CrossRef]
- Van West, P.; Appiah, A.A.; Gow, N.A. Advances in research on oomycete root pathogens. Physiol. Mol. Plant Pathol. 2003, 62, 99–113. [Google Scholar] [CrossRef]
- Stolzy, L.; Sojka, R. Effects of flooding on plant disease. In Flooding and Plant Growth; Academic Press: New York, NY, USA, 1984; pp. 221–264. [Google Scholar]
- Colmer, T.D. Long-distance transport of gases in plants: A perspective on internal aeration and radial oxygen loss from roots. Plant Cell Environ. 2003, 26, 17–36. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.; Armstrong, W. Formation of aerenchyma and the processes of plant ventilation in relation to soil flooding and submergence. Plant Biol. 1999, 1, 274–287. [Google Scholar] [CrossRef]
- Kotula, L.; Ranathunge, K.; Schreiber, L.; Steudle, E. Functional and chemical comparison of apoplastic barriers to radial oxygen loss in roots of rice (Oryza sativa L.) grown in aerated or deoxygenated solution. J. Exp. Bot. 2009, 60, 2155–2167. [Google Scholar] [CrossRef]
- Nishiuchi, S.; Yamauchi, T.; Takahashi, H.; Kotula, L.; Nakazono, M. Mechanisms for coping with submergence and waterlogging in rice. Rice 2012, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Casto, A.L.; McKinley, B.A.; Yu, K.M.J.; Rooney, W.L.; Mullet, J.E. Sorghum stem aerenchyma formation is regulated by SbNAC_D during internode development. Plant Direct 2018, 2, 85. [Google Scholar] [CrossRef] [Green Version]
- Ejiri, M.; Fukao, T.; Miyashita, T.; Shiono, K. A barrier to radial oxygen loss helps the root system cope with waterlogging-induced hypoxia. Breed. Sci. 2021, 71, 40–50. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Df | Adventitious Root Count | Root Disease | Nodulation | Reproductive Node * | Biomass * | Seed Weight | |
|---|---|---|---|---|---|---|---|
| Water (W) | 2 | † NS | 33.2 *** | 34.7 *** | 145.4 *** | 235.7 *** | 304.5 *** |
| PRR (P) | 1 | 5.9 * | 10.3 * | 6.3 * | 15.1 *** | 9.0 *** | 10.2 * |
| Genotype (G) | 7 | 5.5 *** | 3.0 * | 2.9 * | 13.5 *** | 18.4 *** | 32.7 *** |
| P × G | 7 | † NS | † NS | † NS | † NS | † NS | 2.5 * |
| W × P | 2 | 5.6 * | † NS | † NS | 6.3 * | † NS | † NS |
| W × G | 14 | † NS | † NS | 3.2 *** | 2.9 *** | † NS | 10.0 *** |
| W × P × G | 14 | 2.6 * | 2.8 *** | † NS | † NS | 2.1 * | 2.1 * |
| Water | PRR | Geno | Root Disease | Adventitious Root Count | Biomass (g) * | Seed Weight (g) |
|---|---|---|---|---|---|---|
| Field Capacity | − | 04067 | 1 (−0.2) | 24.5 (17.1–31.9) | 3.7 (3.2–4.2) | 0.26 (0.14–0.38) |
| Yorker | 1 (−0.2) | 34.0 (26.6–41.4) | 5.4 (5.0–5.9) | 1.20 (1.08–1.33) | ||
| Rupali | 1 (−0.2) | 31.0 (23.6–38.4) | 6.4 (5.9–6.8) | 1.27 (1.15–1.39) | ||
| + | 04067 | 1 (−0.2) | 19.5 (12.1–26.9) | 3.8 (3.3–4.2) | 0.34 (0.34–0.21) | |
| Yorker | 1.5 (0.3–2.7) | 29.3 (21.8–36.7) | 4.6 (4.1–5.1) | 1.08 0.96–1.21) | ||
| Rupali | 2.8 (1.5–4.0) | 27.5 (20.1–34.9) | 6.7 (6.2–7.2) | 1.31 (1.19–1.43) | ||
| Early WL | − | 04067 | 2.8 (1.5–4.0) | 25.8 (18.3–33.2) | 2.0 (1.5–2.4) | 0.06 (−0.06–0.19) |
| Yorker | 3.0 (1.8–4.2) | 28.5 (21.1–35.9) | 3.1 (2.6–3.5) | 0.65 (0.53–0.77) | ||
| Rupali | 3.5 (2.3–4.7) | 25.0 (17.59–2.41) | 3.0 (2.5–3.5) | 0.49 (0.37–0.61) | ||
| + | 04067 | 3.3 (2.0–4.5) | 34.3 (26.8–41.7) | 2.3 (1.8–2.7) | 0.04 (−0.08–0.16) | |
| Yorker | 3.5 (2.3–4.7) | 42.0 (34.6–49.4) | 3.1 (2.6–3.6) | 0.43 (0.31–0.55) | ||
| Rupali | 4.3 (3.0–5.5) | 36.5 (29.1–43.9) | 3.1 (2.6–3.6) | 0.57 (0.45–0.69) | ||
| Late WL | − | 04067 | 4.0 (2.8–5.2) | 28.3 (20.8–35.7) | 1.7 (1.2–2.2) | 0.00 (−0.12–0.12) |
| Yorker | 4.0 (2.8–5.2) | 49.3 (41.8–56.7) | 2.2 (1.7–2.7) | 0.25 (0.25–0.12) | ||
| Rupali | 5.3 (4.0–6.5) | 24.5 (17.1–31.9) | 1.6 (1.1–2.0) | 0.06 (−0.07–0.18) | ||
| + | 04067 | 6.8 (5.5–8.0) | 10.8 (3.3–18.2) | 0.9 (0.5–1.4) | 0.00 (−0.12–0.12) | |
| Yorker | 7.0 (5.8–8.2) | 18.8 (11.3–26.2) | 1.4 (1.0–1.9) | 0.01 (−0.11–0.13) | ||
| Rupali | 7.3 (6.0–8.5) | 13.3 (5.8–20.7) | 1.4(1.0–1.9) | 0.00 (−0.12–0.12) |
| Treatment | Genotype | Shoot Weight (g) | Root Weight (g) | Chlorosis Score (1–9) | Root Disease Score (1–9) | P. med kDNA Copies/g Soil (Log10) |
|---|---|---|---|---|---|---|
| Control (FC) | 04067 | 2.5 (2.1–2.9) | 0.7 (0.6–0.8) | 1.0 | 1 (0.5–1.5) | 0 |
| Yorker | 4.0 (3.6–4.4) | 0.9 (0.8–1.0) | 1.0 | 1 (0.5–1.5) | 0 | |
| Rupali | 4.6 (4.2–5.0) | 0.9 (0.8–1.0) | 1.0 | 1 (0.5–1.5) | 0 | |
| PRR (FC) | 04067 | 2.9 (2.5–3.3) | 0.8 (0.7–0.9) | 1.0 (0.2–1.8) | 1.0 (0.5–1.5) | 1.5 (1.4–1.6) |
| Yorker | 3.8 (3.4–4.3) | 1.1 (1.0–1.2) | 1.0 (0.2–1.8) | 1.0 (0.5–1.5) | 1.5 (1.4–1.6) | |
| Rupali | 4.7 (4.3–5.1) | 1.0 (0.9–1.1) | 1.3 (0.6–2.1) | 2.5 (2.0–3.0) | 1.5 (1.4–1.6) | |
| PRR + transient WL (48 h) | 04067 | 2.4 (1.9–2.8) | 0.7 (0.6–0.8) | 1.0 (0.2–1.8) | 1.2 (0.7–1.6) | 1.6 (1.5–1.7) |
| Yorker | 3.3 (2.9–3.8) | 0.9 (0.8–1.0) | 1.3 (0.6–2.1) | 1.5 (1.0–2.0) | 1.6 (1.5–1.7) | |
| Rupali | 3.8 (3.4–4.3) | 0.9 (0.8–1.0) | 2.2 (1.4–3.0) | 2.2 (1.7–1.6) | 1.6 (1.5–1.7) | |
| Inoculated + repeated transient WL (48 h + 48 h) | 04067 | 2.2 (1.8–2.6) | 0.7 (0.5–0.7) | 1.0 (0.2–1.8) | 2.5 (2.0–3.0) | 1.6 (1.5–1.7) |
| Yorker | 3.0 (2.6–3.5) | 0.9 (0.8–1.0) | 1.8 (1.1–2.6) | 2.8 (2.4–3.3) | 1.7 (1.6–1.8) | |
| Rupali | 3.1 (2.7–3.5) | 0.9 (0.8–1.0) | 2.3 (1.6–3.1) | 3.8 (3.4–4.3) | 1.7 (1.6–1.8) | |
| PRR + WL (240 h) | 04067 | 1.6 (1.2–2.0) | 0.5 (0.4–0.7) | 2.7 (1.9–3.4) | 3.8 (3.4–4.3) | 1.6 (1.5–1.7) |
| Yorker | 1.9 (1.5–2.3) | 0.6 (0.5–0.7) | 5.0 (4.2–5.8) | 5.2 (4.7–5.6) | 1.5 (1.4–1.6) | |
| Rupali | 1.6 (1.2–2.0) | 0.5 (0.4–0.6) | 6.8 (6.1–7.6) | 6.8 (6.4–7.3) | 1.8 (1.7–1.9) | |
| df | F-value | |||||
| Treatment (T) | 4 | 36.3 *** | 10.8 *** | 42.1 *** | 112.1 *** | † NS |
| Genotype (G) | 2 | 36.9 *** | 19.0 *** | 14.6 *** | 30.6 *** | † NS |
| T × G | 14 | 2.9 * | † NS | 3.4 * | 4.0 * | † NS |
| Treatment | Genotype | Shoot Weight (g) | Root Weight (g) | Chlorosis Score (1–9) | Root Disease Score (1–9) |
|---|---|---|---|---|---|
| Aerated control | 04067 | 1.8 (1.5–2.0) | 1.8 (1.5–2.3) | 1 (0.5–1.5) | 1 (0.5–1.5) |
| Yorker | 2.2 (1.9–2.4) | 2.8 (2.5–3.2) | 1 (0.5–1.5) | 1 (0.5–1.5) | |
| Rupali | 2.6 (2.4–2.6) | 3.6 (3.2–4.0) | 1 (0.5–1.5) | 1 (0.5–1.5) | |
| Aerated PRR | 04067 | 1.1 (0.9–1.4) | 1.9 (1.5–2.3) | 1 (0.5–1.5) | 3.2 (2.7–3.7) |
| Yorker | 1.2 (1.0–1.5) | 2.4 (2.0–2.7) | 1 (0.5–1.5) | 4.2 (3.7–4.7) | |
| Rupali | 1.3 (1.1–1.52) | 2.8 (2.4–3.2) | 1 (0.5–1.5) | 4.3 (3.9–4.8) | |
| Hypoxic PRR | 04067 | 0.5 (0.2–0.7) | 0.8 (0.4–1.2) | 2.3 (1.9–2.8) | 3.7 (3.2–4.2) |
| Yorker | 0.4 (0.2–0.7) | 1.1 (0.7–1.4) | 2.3 (1.9–2.8) | 4.2 (3.7–4.7) | |
| Rupali | 0.6 (0.4–0.8) | 1.2 (0.8–1.6) | 3.3 (2.9–3.8) | 6.8 (6.4–7.3) | |
| Df | F-value | ||||
| Treatment (T) | 2 | 109.0 *** | 53.3 *** | 34.9 *** | 267.3 *** |
| Genotype (G) | 2 | 5.5 * | 16.5 *** | † NS | 36.5 *** |
| T × G | 8 | † NS | † NS | † NS | 23.7 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dron, N.; Simpfendorfer, S.; Sutton, T.; Pengilley, G.; Hobson, K. Cause of Death: Phytophthora or Flood? Effects of Waterlogging on Phytophthora medicaginis and Resistance of Chickpea (Cicer arietinum). Agronomy 2022, 12, 89. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010089
Dron N, Simpfendorfer S, Sutton T, Pengilley G, Hobson K. Cause of Death: Phytophthora or Flood? Effects of Waterlogging on Phytophthora medicaginis and Resistance of Chickpea (Cicer arietinum). Agronomy. 2022; 12(1):89. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010089
Chicago/Turabian StyleDron, Nicole, Steven Simpfendorfer, Tim Sutton, Georgina Pengilley, and Kristy Hobson. 2022. "Cause of Death: Phytophthora or Flood? Effects of Waterlogging on Phytophthora medicaginis and Resistance of Chickpea (Cicer arietinum)" Agronomy 12, no. 1: 89. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12010089