
Alleviation of Drought Stress in Soybean by Applying a Biostimulant Based on Amino Acids and Macro- and Micronutrients

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Layout
2.2. Determination of Physiological Variables
2.3. Determination of Antioxidant Compound and Enzymes
2.4. Determination of Biometric Parameters of Plants
2.5. Determination of Production Components
2.6. Statistical Analysis
3. Results
3.1. Physiological Variables
3.2. Antioxidant Compound and Enzymes
3.3. Biometric Components
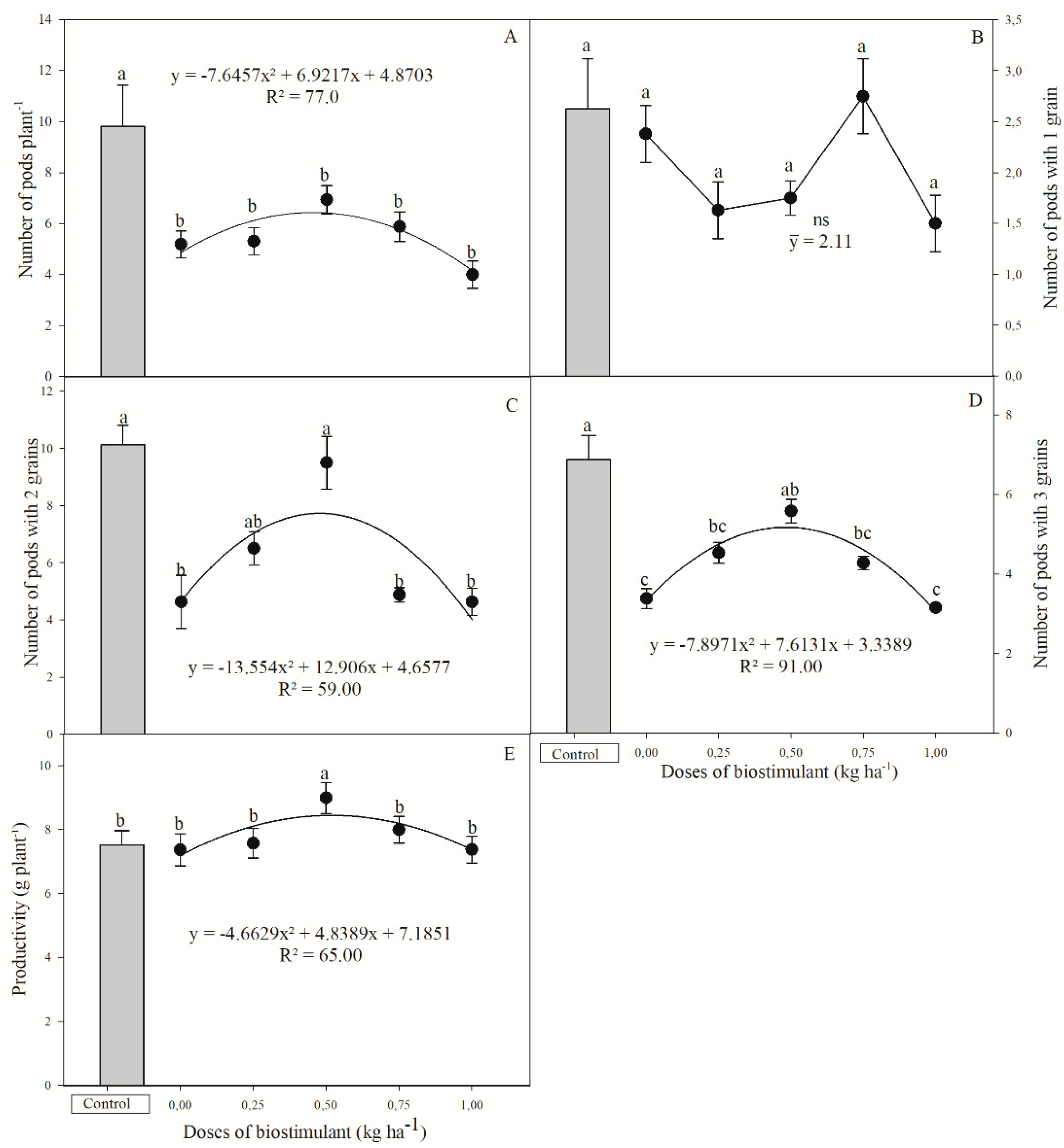
3.4. Production Components
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farouk, S.; Al-Huqail, A.A. Sodium nitroprusside application regulates antioxidant capacity, improves phytopharmaceutical production and essential oil yield of marjoram herb under drought. Ind. Crops Prod. 2020, 158, 113034. [Google Scholar] [CrossRef]
- Farouk, S.; Al-Ghamdi, A.A.M. Sodium nitroprusside application enhances drought tolerance in marjoram herb by promoting chlorophyll biosynthesis and enhancing osmotic adjustment capacity. Arab. J. Geosci. 2021, 14, 430. [Google Scholar] [CrossRef]
- El-Gamal, S.M.A.; El-Din, W.M.S.; Farouk, S.; Moktar, N.A.Y.O. Integrated effects of biochar and potassium silicate on borage plant under different irrigation regimes in sandy soil. J. Hortic. Sci. Ornam. Plants 2021, 13, 60–76. [Google Scholar] [CrossRef]
- Dong, S.; Wang, W.; Jiang, Y.; Ma, Z.; Yan, C.; Liu, L.; Cui, G. Antioxidant and proteomic analysis of soybean response to drought during soybean flowering. Ekoloji 2019, 28, 2041–2052. [Google Scholar]
- Anda, A.; Soós, G.; Menyhárt, L.; Kucserka, T.; Simon, B. Yield features of two soybean varieties under different water supplies and field conditions. Field Crops Res. 2020, 245, 107673. [Google Scholar] [CrossRef]
- Wijewardana, C.; Alsajri, F.A.; Irby, J.T.; Golden, B.R.; Henry, W.B.; Reddy, K.R. Water deficit effects on Soybean root morphology and early-season vigor. Agronomy 2019, 9, 826. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef]
- Bellaloui, N.; Mengistu, A.; Fisher, D.K.; Abel, C.A. Soybean seed composition constituents as affected by drought and phomopsisin phomopsis susceptible and resistant genotypes. J. Crop Improv. 2012, 26, 428–453. [Google Scholar] [CrossRef]
- Wijewardana, C.; Reddy, K.R.; Krutz, L.J.; Gao, W.; Bellaloui, N. Drought stress has transgenerational effects on soybean seed germination and seedling vigor. PLoS ONE 2019, 14, e0214977. [Google Scholar] [CrossRef]
- Wijewardana, C.; Reddy, K.R.; Alsajri, F.A.; Irby, J.T.; Krutz, J.; Golden, B.R. Quantifying soil moisture deficit effects on soybean yield and yield component distribution patterns. Irrig. Sci. 2018, 36, 241–255. [Google Scholar] [CrossRef]
- Wijewardana, C.; Reddy, K.R.; Bellaloui, N. Soybean seed physiology, quality, and chemical composition under soil moisture stress. Food Chem. 2019, 278, 92–100. [Google Scholar] [CrossRef]
- Rosa, V.R.; Santos, A.L.F.; Silva, A.A.; Sab, M.P.V.; Germino, G.H.; Cardoso, F.B.; Silva, M.A. Increased soybean tolerance to water deficiency through biostimulant based on fulvic acids and Ascophyllum nodosum (L.) seaweed extract. Plant Physiol. Biochem. 2021, 158, 228–243. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E. Fisiologia e Desenvolvimento Vegetal, 6th ed.; Artmed: Porto Alegre, Brazil, 2017; 888p. [Google Scholar]
- Shukla, P.S.; Shotton, K.; Norman, E.; Neily, W.; Critchley, A.T.; Prithiviraj, B. Seaweed extract improve drought tolerance of soybean by regulating stress-response genes. AoB PLANTS 2018, 10, 051. [Google Scholar] [CrossRef]
- Schmidt, R.R.; Weits, D.A.; Feulner, C.F.J.; Dongen, J.T.V. Oxygen sensing and integrative stress signaling in plants. Plant Physiol. 2018, 176, 1131–1142. [Google Scholar] [CrossRef]
- Farouk, S.; Al-Huqail, A.A. Sustainable biochar and/or melatonin improve salinity tolerance in borage plants by modulating osmotic adjustment, antioxidants, and ion homeostasis. Plants 2022, 11, 765. [Google Scholar] [CrossRef]
- Podgórska, A.; Burian, M.; Szal, B. Extra-cellular but extra-ordinarily important for cells: Apoplastic reactive oxygen species metabolism. Front. Plant Sci. 2017, 8, 1353. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, K.D. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Repke, R.A.; Silva, D.M.R.; Santos, J.C.C.; Silva, M.A. Increased soybean tolerance to high-temperature through biostimulant based on Ascophyllum nodosum (L.) seaweed extract. J. Appl. Phycol. 2022, 11, 1–14. [Google Scholar] [CrossRef]
- Mącik, M.; Gryta, A.; Frąc, M. Biostimulants in agriculture: An overview on concepts, strategies and effects on soil microorganisms. Adv. Agron. 2020, 162, 31–87. [Google Scholar] [CrossRef]
- Kocira, S.; Szparaga, A.; Kocira, A.; Czerwińska, E.; Wójtowicz, A.; Bronowicka-Mielniczuk, U.; Koszel, M.; Findura, P. Modeling biometric traits, yield and nutritional and antioxidant properties of seeds of three soybean cultivars through the application of biostimulant containing seaweed and amino acids. Front. Plant Sci. 2018, 9, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariani, L.; Ferrante, A. Agronomic management for enhancing plant tolerance to abiotic stresses-drought, salinity, hypoxia, and lodging. Horticulturae 2017, 4, 52. [Google Scholar] [CrossRef]
- Wozniak, E.; Blaszczak, A.; Wiatrak, P.; Canady, M. Mode of Action Biostimulant. In The Chemical Biology of Plant Biostimulants; Geelen, D., Xu, L., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020. [Google Scholar] [CrossRef]
- Fleming, T.R.; Fleming, C.C.; Levy, C.C.B.; Repiso, C.; Hennequart, F.; Nolasco, J.B.; Liu, F. Biostimulants enhance growth and drought tolerance in Arabidopsis thaliana and exhibit chemical priming action. Ann. Appl. Biol. 2019, 174, 153–165. [Google Scholar] [CrossRef]
- Raij, B.V.; Cantarella, H.; Quaggio, J.A.; Furlani, A.M.C. Recomendações de Adubação e Calagem Para o Estado de São Paulo; Instituto Agronômico & Fundação IAC: Campinas, Brasil, 1996; 154p. [Google Scholar]
- Del Longo, O.T.; Gonzáles, C.A.; Pastori, G.M.; Trippi, V.S. Antioxidant defenses under hyperoxygenic and hyperosmotic conditions in leaves of two lines of maize with differential sensitivity to drought. Plant Cell Physiol. 1993, 34, 1023–1028. [Google Scholar] [CrossRef]
- Havir, E.A.; McHale, N.A. Biochemical and Developmental Characterization of Multiple Forms of Catalase in Tobacco Leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, Peroxidase and Polyphenol Oxidase Activities during Rice Leaf Senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. Assay of Catalase and Peroxidase. Methods Enzymol. 1955, 2, 764–775. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Jaworski, E.G. Nitrate reductase assay in intact plant tissues. Biochem. Biophys. Res. Commun. 1971, 43, 1274–1279. [Google Scholar] [CrossRef]
- Ferreira, D.F. Sisvar: A guide for its bootstrap procedures in multiple comparisons. Ciência Agrotecnologia 2014, 38, 109–112. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Aires, E.S.; Ferraz, A.K.L.; Carvalho, B.L.; Teixeira, F.P.; Putti, F.F.; Souza, E.P.; Rodrigues, J.D.; Ono, E.O. Foliar application of salicylic acid to mitigate water stress in tomato. Plants 2022, 11, 1775. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000 Res. 2016, 5, 1554. [Google Scholar] [CrossRef] [PubMed]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef]
- Siddique, Z.; Jan, S.; Imadi, S.R.; Gul, A.; Ahmad, P. Drought stress and photosynthesis in plants. In Water Stress and Crop Plants: A Sustainable Approach; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2016; Volume 10, pp. 1–11. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.; Yang, C.; Du, S.; Yang, W. Photosynthetic performance of soybean plants to water deficit under high and low light intensity. S. Afr. J. Bot. 2016, 105, 279–287. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Arabzadeh, N. The effect of drought stress on soluble carbohydrates (sugars) in two species of Haloxylon persicum and Haloxylon aphyllum. Asian J. Plant Sci. 2012, 11, 44–51. [Google Scholar] [CrossRef]
- Santaniello, A.; Scartazza, A.; Gresta, F.; Loreti, E.; Biasone, A.; Di, T.D.; Piaggesi, A.; Perata, P. Ascophyllum nodosum seaweed extract alleviates drought stress in Arabidopsis by affecting photosynthetic performance and related gene expression. Front. Plant Sci. 2017, 8, 1362. [Google Scholar] [CrossRef] [Green Version]
- Prado, K.; Boursiac, Y.; Tournaire-Roux, C.; Monneuse, J.M.; Postaire, O.; Ines, O.; Maurel, C. Regulation of Arabidopsis leaf hydraulics involves light-dependent phosphorylation of aquaporins in veins. Plant Cell 2013, 25, 1029–1039. [Google Scholar] [CrossRef]
- Cohen, A.S.; Attia, Z.; Moshelion, M. Bundle-sheath cell regulation of xylem-mesophyll water transport via aquaporins under drought stress: A target of xylem-borne ABA? Plant J. 2011, 67, 72–80. [Google Scholar] [CrossRef]
- Silva, M.A.; Santos, C.M.; Vitorino, H.S.; Rhein, A.F.L. Pigmentos fotossintéticos e índice SPAD como descritores de intensidade do estresse por deficiência hídrica em cana-de-açúcar. Biosci. J. 2014, 30, 173–181. [Google Scholar]
- Oliver, M.J.; Farrant, J.M.; Hilhorst, H.W.M.; Mundree, S.; Brett, W.; Bewley, J.D. Desiccation tolerance: Avoiding cellular damage during drying and rehydration. Annu. Rev. Plant Biol. 2020, 71, 435–460. [Google Scholar] [CrossRef]
- Caverzan, A.; Casassola, A.; Brammer, S.P. Reactive oxygen species and antioxidant enzymes involved in plant tolerance to stress. In Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives; Shanker, A.K., Shanker, C., Eds.; InTech: Rijeka, Croatia, 2016; pp. 463–480. [Google Scholar]
- Rivas, R.; Frosi, G.; Ramos, D.G.; Pereira, S.; Benko-Iseppon, A.M.; Santos, M.G. Photosynthetic limitation and mechanisms of photoprotection under drought and recovery of Calotropis procera, an evergreen C3 from arid regions. Plant Physiol. Biochem. 2017, 118, 589–599. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. Oxidative stress and antioxidative systems: Recipes for successful data collection and interpretation. Plant Cell Environ. 2016, 39, 1140–1160. [Google Scholar] [CrossRef]
- Kocira, S. Effect of amino acid biostimulant on the yield and nutraceutical potential of soybean. Chil. J. Agric. Res. 2019, 79, 17–25. [Google Scholar] [CrossRef]
- Dat, J.F.; Foyer, C.H.; Scott, I.M. Changes in salicylic acid and antioxidants during induced thermotolerance in mustard seedlings. Plant Physiol. 1998, 118, 1455–1461. [Google Scholar] [CrossRef]
- Teixeira, W.F.; Soares, L.H.; Fagan, E.B.; Mello, S.C.; Reichardt, K.; Dourado-Neto, D. Amino acids as stress reducers in soybean plant growth under different water-deficit conditions. J. Plant Growth Regul. 2019, 39, 905–919. [Google Scholar] [CrossRef]
- Meena, M.; Davyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, 02952. [Google Scholar] [CrossRef]
- Merward, A.-R.M.A.; Desoky, E.-S.M.; Rady, M.M. Response of water deficit-stressed Vigna unguiculata performances to silicon, proline or methionine foliar application. Sci. Hortic. 2018, 228, 132–144. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, F.; Kato, M.; Hyodo, H.; Ikoma, Y.; Sugiura, M.; Yano, M. Effect of sucrose on ascorbate level and expression of genes involved in the ascorbate biosynthesis and recycling pathway in harvested broccoli florets. J. Exp. Bot. 2005, 56, 65–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | OM | Presin | K | Ca | Mg | H + Al | SB | CEC | V | Clay | Silt | Sand | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CaCl2 | g dm−3 | mg dm−3 | mmolc dm−3 | % | g kg−1 | ||||||||
| 5.4 | 24 | 15 | 6.7 | 36 | 14 | 32 | 57 | 89 | 64 | 614 | 196 | 190 | |
| Composition | (w w−1) |
|---|---|
| Nitrogen (N) | 4.0% |
| Phosphorous (P2O5) | 21.0% |
| Iron (Fe EDTA) | 0.5% |
| Copper (Cu EDTA) | 0.3% |
| Boron (B) | 0.5% |
| Manganese (Mn EDTA) | 3.0% |
| Zinc (Zn EDTA) | 3.0% |
| Glycine betaine | 12.0% |
| pH (1%) | 3.6% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Repke, R.A.; Silva, D.M.R.; dos Santos, J.C.C.; de Almeida Silva, M. Alleviation of Drought Stress in Soybean by Applying a Biostimulant Based on Amino Acids and Macro- and Micronutrients. Agronomy 2022, 12, 2244. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12102244
Repke RA, Silva DMR, dos Santos JCC, de Almeida Silva M. Alleviation of Drought Stress in Soybean by Applying a Biostimulant Based on Amino Acids and Macro- and Micronutrients. Agronomy. 2022; 12(10):2244. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12102244
Chicago/Turabian StyleRepke, Rodrigo Alberto, Dayane Mércia Ribeiro Silva, Jania Claudia Camilo dos Santos, and Marcelo de Almeida Silva. 2022. "Alleviation of Drought Stress in Soybean by Applying a Biostimulant Based on Amino Acids and Macro- and Micronutrients" Agronomy 12, no. 10: 2244. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12102244