
Role of ZmGST Gene Family Involved in Nicosulfuron Stress Tolerance Revealed by Genomic and Transcriptomic Analyses

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
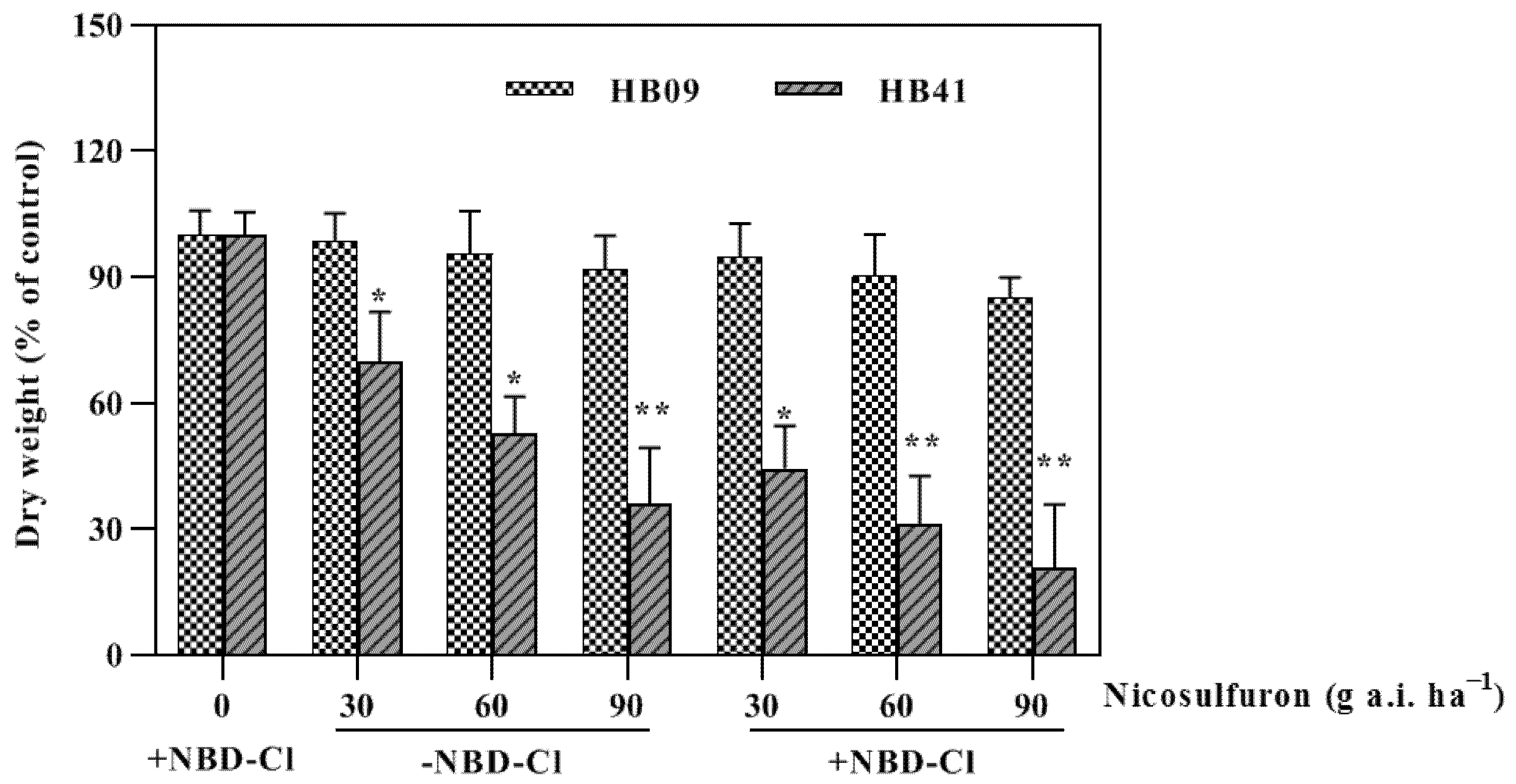
2.1. Effect of NBD-Cl Addition to Nicosulfuron
2.2. Identification, Sequence Analysis, and Nomenclature of ZmGSTs
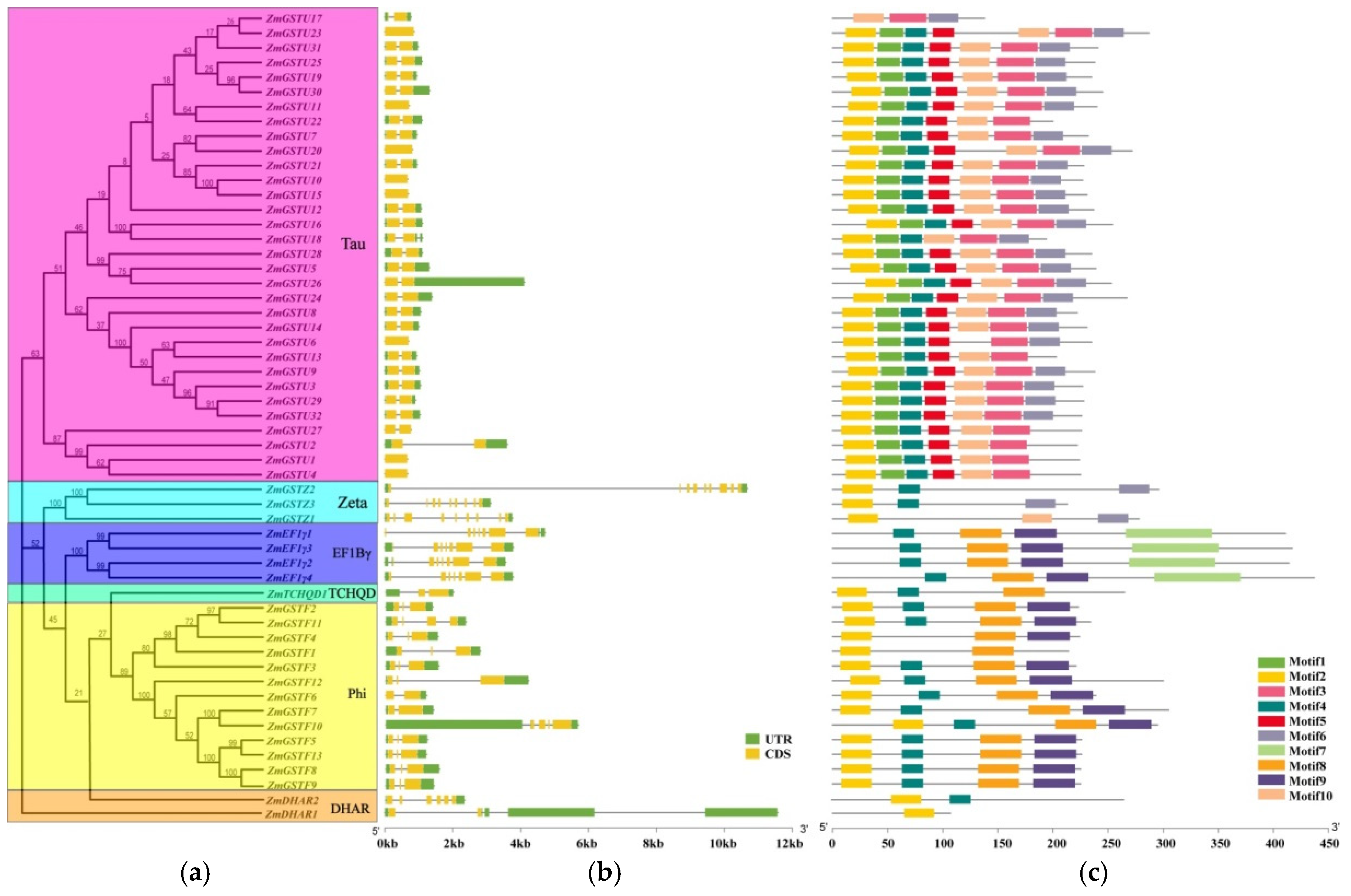
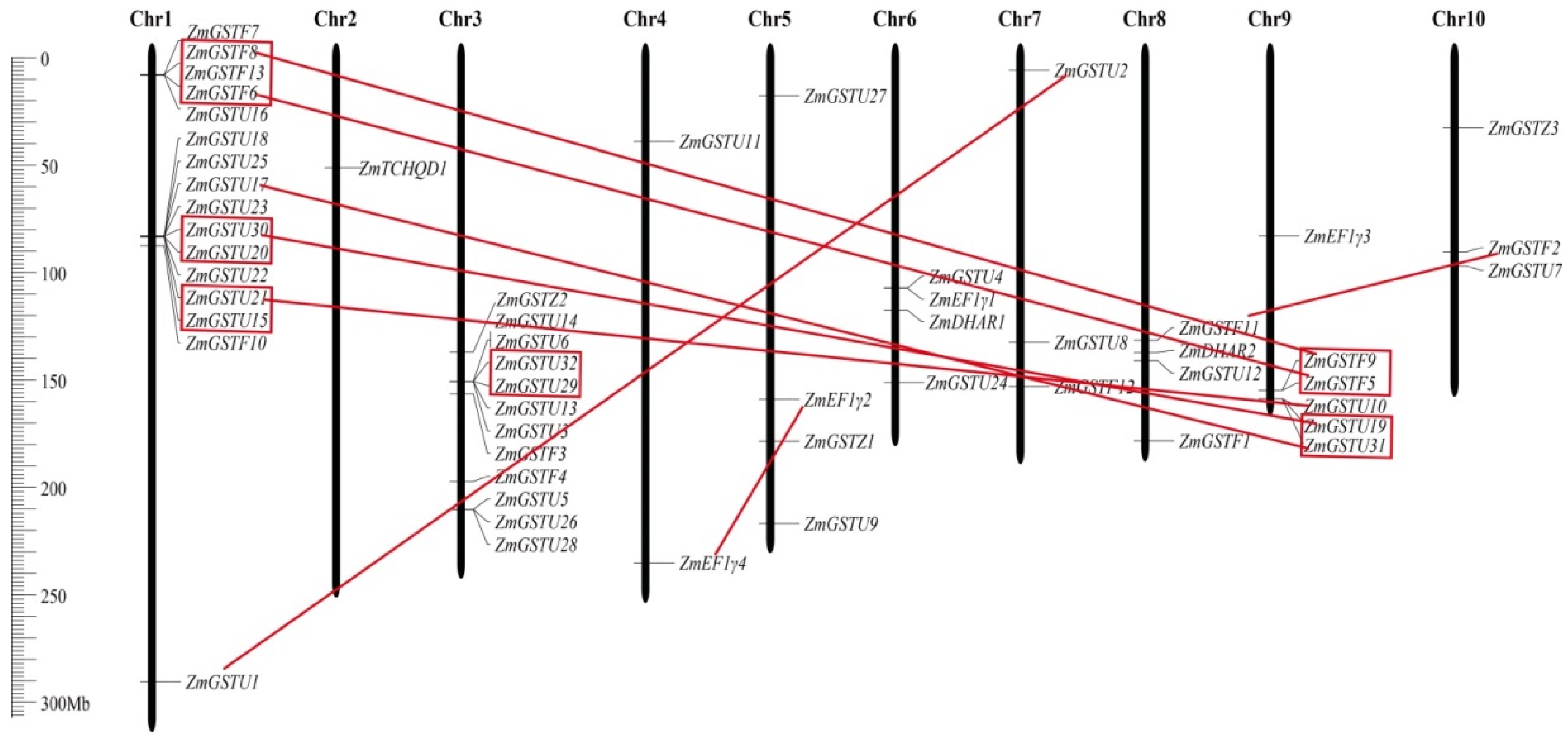
2.3. Gene Structure, Chromosomal Location, and Phylogenetic Analysis of ZmGSTs
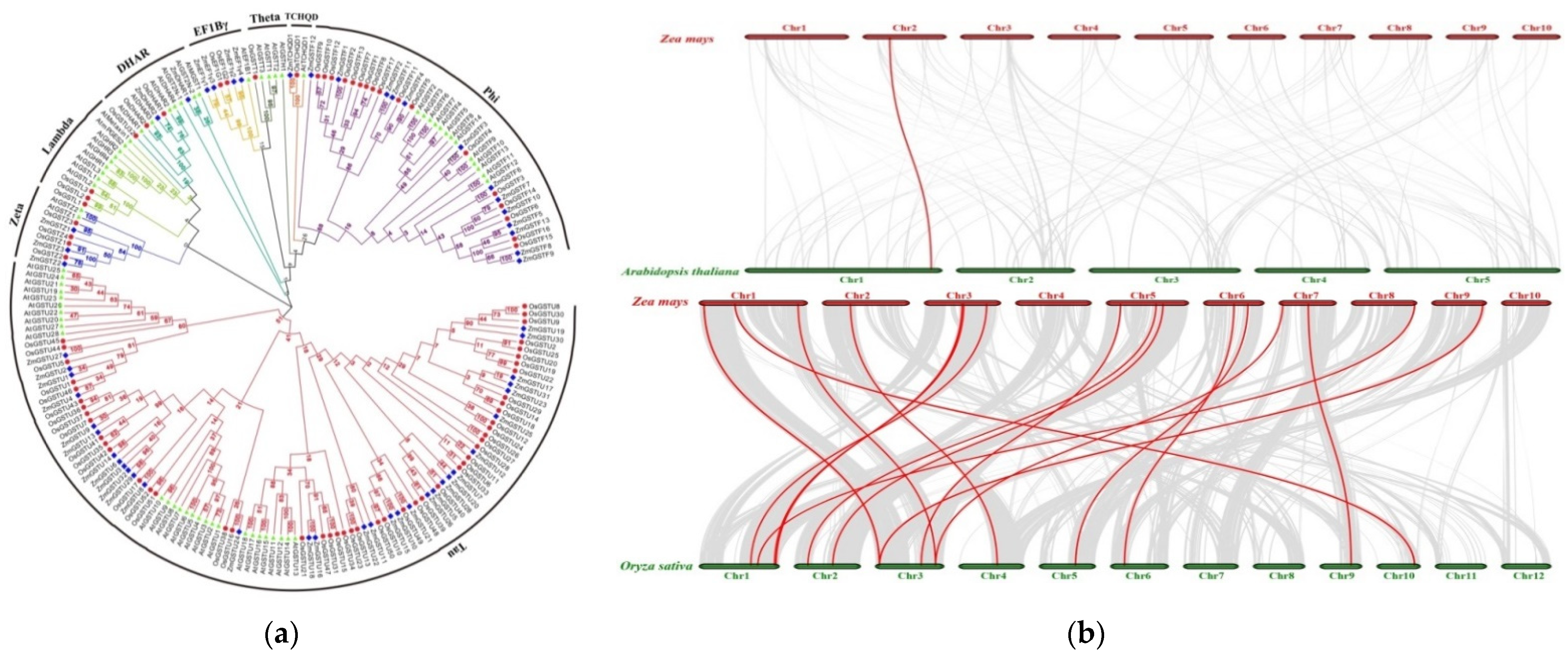
2.4. Phylogenetic and Synteny Analysis of ZmGSTs
2.5. Transcriptome Sequencing, Gene Expression Analysis of ZmGST Genes
2.6. Quantitative RT-PCR Validation Experiments
3. Results
3.1. Effect of GST Inhibitors on Nicosulfuron Photoxicity to Maize Plants
3.2. Identification and Characterization of the GST Gene Family in Zea mays
3.3. Gene Structure, Chromosomal Localization, and Gene Duplication of ZmGST Genes
3.4. Phylogenetic and Synteny Analysis of GSTs in Selected Species
3.5. Expression Profiles of Potential Nicosulfuron Stress-Responsive ZmGST Genes
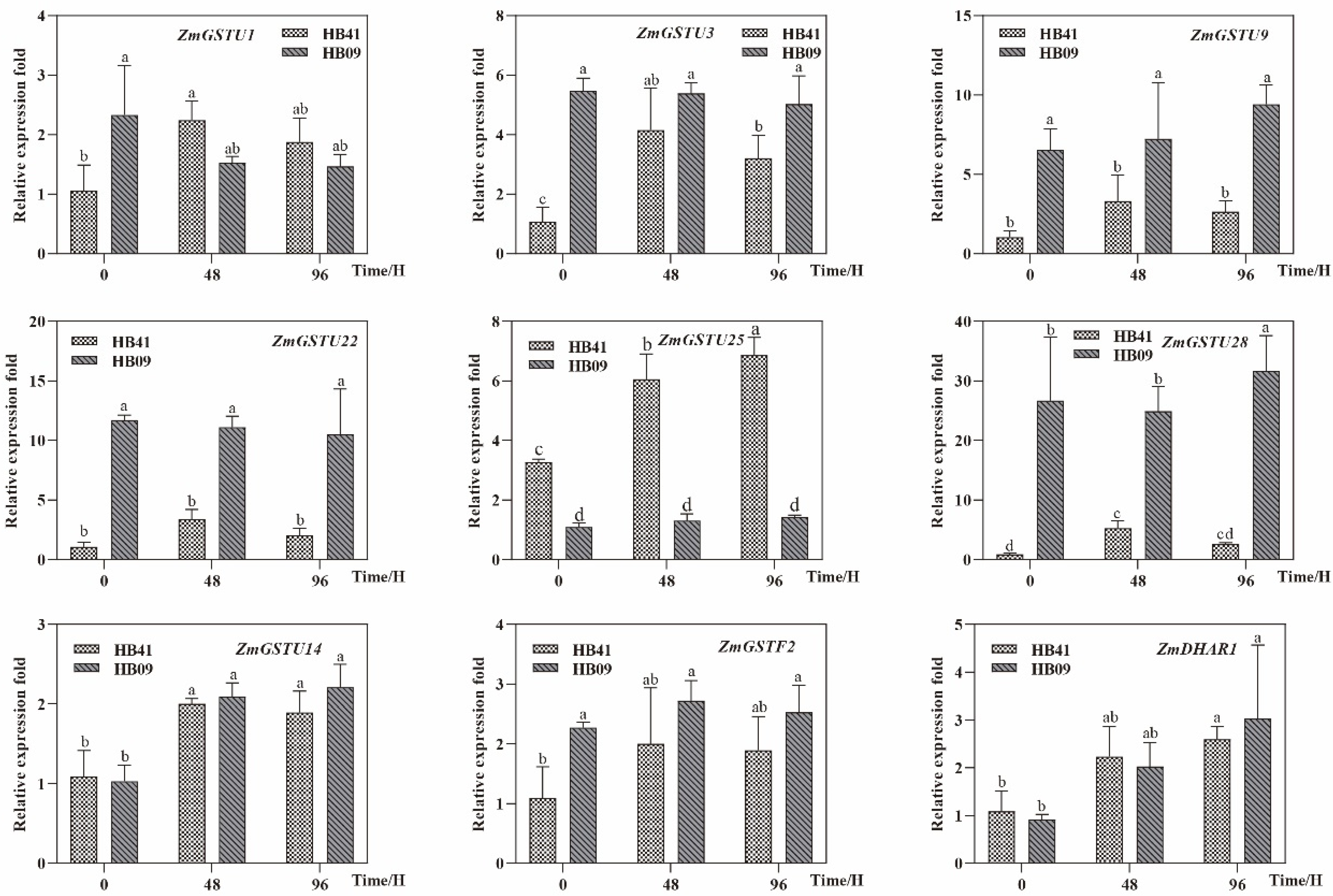
3.6. Expression of ZmGSTs Using qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Zhong, X.; Li, F.; Shi, Z. Effects of nicosulfuron on growth, oxidative damage, and the ascorbate-glutathione pathway in paired nearly isogenic lines of waxy maize (Zea mays L.). Pestic. Biochem. Phys. 2018, 145, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Cummins, I.; Dixon, D.P.; Freitag-Pohl, S.; Skipsey, M.; Edwards, R. Multiple roles for plant glutathione transferases in xenobiotic detoxification. Drug Metab. Rev. 2011, 43, 266–280. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; Edwards, R. Glutathione Transferases. In Arabidopsis Book; The American Society of Plant Biologists: Rockville, MD, USA, 2010; Volume 8, p. e0131. [Google Scholar]
- Jain, M.; Ghanashyam, C.; Bhattacharjee, A. Comprehensive expression analysis suggests overlapping and specific roles of rice glutathione S-transferase genes during development and stress responses. BMC Genom. 2010, 11, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.J.; Tang, Z.X.; Han, X.M.; Yang, Z.L.; Zhang, F.M.; Yang, H.L.; Liu, Y.J.; Zeng, Q.Y. Divergence in Enzymatic Activities in the Soybean GST Supergene Family Provides New Insight into the Evolutionary Dynamics of Whole-Genome Duplicates. Mol. Biol. Evol. 2015, 32, 2844–2859. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Ma, J.; Zhang, Q.; Wu, C.; Zhao, H.; Wu, Y.; Yang, G.; He, G. Genome-wide identification and expression profiling of glutathione transferase gene family under multiple stresses and hormone treatments in wheat (Triticum aestivum L.). BMC Genom. 2019, 20, 986. [Google Scholar] [CrossRef] [Green Version]
- Rezaei, M.K.; Shobbar, Z.-S.; Shahbazi, M.; Abedini, R.; Zare, S. Glutathione S-transferase (GST) family in barley: Identification of members, enzyme activity, and gene expression pattern. J. Plant Physiol. 2013, 170, 1277–1284. [Google Scholar] [CrossRef]
- Islam, M.S.; Choudhury, M.; Majlish, A.-N.K.; Islam, T.; Ghosh, A. Comprehensive genome-wide analysis of Glutathione S-transferase gene family in potato (Solanum tuberosum L.) and their expression profiling in various anatomical tissues and perturbation conditions. Gene 2018, 639, 149–162. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, Y.; Meng, Q.; Shi, F.; Liu, J.; Li, Y. Molecular cloning, identification of GSTs family in sunflower and their regulatory roles in biotic and abiotic stress. World J. Microb. Biot. 2018, 34, 109. [Google Scholar] [CrossRef]
- Lallement, P.A.; Brouwer, B.; Keech, O.; Hecker, A.; Rouhier, N. The still mysterious roles of cysteine-containing glutathione transferases in plants. Front. Pharmacol. 2014, 5, 192. [Google Scholar] [CrossRef] [Green Version]
- Nianiou-Obeidat, I.; Madesis, P.; Kissoudis, C.; Voulgari, G.; Chronopoulou, E.; Tsaftaris, A.; Labrou, N.E. Plant glutathione transferase-mediated stress tolerance: Functions and biotechnological applications. Plant Cell Rep. 2017, 36, 791–805. [Google Scholar] [CrossRef]
- Strom, S.A.; Hager, A.G.; Concepcion, J.C.T.; Seiter, N.J.; Davis, A.S.; Morris, J.A.; Kaundun, S.S.; Riechers, D.E. Metabolic Pathways for S-Metolachlor Detoxification Differ Between Tolerant Corn and Multiple-Resistant Waterhemp. Plant Cell Physiol. 2021, 62, 1770–1785. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; Skipsey, M.; Edwards, R. Roles for glutathione transferases in plant secondary metabolism. Phytochemistry 2010, 71, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Labrou, N.E.; Papageorgiou, A.C.; Pavli, O.; Flemetakis, E. Plant GSTome: Structure and functional role in xenome network and plant stress response. Curr. Opin. Biotech. 2015, 32, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Frear, D.; Swanson, H.R. Biosynthesis of S-(4-ethylamino-6-isopropylamino-2-s-triazino) glutathione: Partial purification and properties of a glutathione S-transferase from corn. Phytochemistry 1970, 9, 2123–2132. [Google Scholar] [CrossRef]
- Zhang, J.J.; Yang, H. Metabolism and detoxification of pesticides in plants. Sci. Total Environ. 2021, 790, 148034. [Google Scholar] [CrossRef]
- Georgakis, N.; Poudel, N.; Papageorgiou, A.C.; Labrou, N.E. Comparative structural and functional analysis of phi class glutathione transferases involved in multiple-herbicide resistance of grass weeds and crops. Plant Physiol. Bioch. 2020, 149, 266–276. [Google Scholar] [CrossRef]
- Chronopoulou, E.; Madesis, P.; Asimakopoulou, B.; Platis, D.; Tsaftaris, A.; Labrou, N.E. Catalytic and structural diversity of the fluazifop-inducible glutathione transferases from Phaseolus vulgaris. Planta 2012, 235, 1253–1269. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.Y.; Kong, K.H. Study on the biochemical characterization of herbicide detoxification enzyme, glutathione S-transferase. Biofactors 2007, 30, 281–287. [Google Scholar] [CrossRef]
- Chronopoulou, E.; Madesis, P.; Tsaftaris, A.; Labrou, N.E. Cloning and Characterization of a Biotic-Stress-Inducible Glutathione Transferase from Phaseolus vulgaris. Appl. Biochem. Biotech. 2014, 172, 595–609. [Google Scholar] [CrossRef] [Green Version]
- Goggin, D.E.; Cawthray, G.R.; Flematti, G.R.; Bringans, S.D.; Lim, H.; Beckie, H.J.; Busi, R. Pyroxasulfone-Resistant Annual Ryegrass (Lolium rigidum) Has Enhanced Capacity for Glutathione Transferase-Mediated Pyroxasulfone Conjugation. J. Agric. Food Chem. 2021, 69, 6414–6422. [Google Scholar] [CrossRef]
- Rangani, G.; Noguera, M.; Salas-Perez, R.; Benedetti, L.; Roma-Burgos, N. Mechanism of Resistance to S-metolachlor in Palmer amaranth. Front. Plant Sci. 2021, 12, 652581. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Gao, H.; Xia, W.; Zhang, T.; Dong, L. Establishing a herbicide-metabolizing enzyme library in Beckmannia syzigachne to identify genes associated with metabolic resistance. J. Exp. Bot. 2016, erv565. [Google Scholar]
- Zhao, N.; Li, W.; Bai, S.; Guo, W.; Yuan, G.; Wang, F.; Liu, W.; Wang, J. Transcriptome Profiling to Identify Genes Involved in Mesosulfuron-Methyl Resistance in Alopecurus aequalis. Front. Plant Sci. 2017, 8, 1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, L.; Guo, Q.; Wang, J.; Shi, L.; Yang, X.; Zhou, Y.; Yu, Q.; Bai, L. CYP81A68 confers metabolic resistance to ALS and ACCase-inhibiting herbicides and its epigenetic regulation in Echinochloa crus-galli. J. Hazard Mater. 2022, 34, 128225. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wu, L.; Wang, J.; Yu, Q.; Bai, L.; Pan, L. Quizalofop-p-ethyl resistance in Polypogon fugax involves glutathione S-transferases. Pest Manag. Sci. 2020, 76, 3800–3805. [Google Scholar] [CrossRef]
- Lin, F.; Jiang, L.; Liu, Y.; Lv, Y.; Dai, H.; Zhao, H. Genome-wide identification of housekeeping genes in maize. Plant Mol. Biol. 2014, 86, 543–554. [Google Scholar] [CrossRef]
- Kõressaar, T.; Lepamets, M.; Kaplinski, L.; Raime, K.; Andreson, R.; Remm, M. Primer3_masker: Integrating masking of template sequence with primer design software. Bioinformatics 2018, 34, 1937–1938. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Bi, B.; Xu, X.; Li, B.; Tian, S.; Wang, J.; Zhang, H.; Wang, G.; Han, Y.; McElroy, J.S. Rapid identification of a candidate nicosulfuron sensitivity gene (Nss) in maize (Zea mays L.) via combining bulked segregant analysis and RNA-seq. Theor. Appl. Genet. 2019, 132, 1351–1361. [Google Scholar] [CrossRef]
- Boulahia, K.; Carol, P.; Planchais, S.; Abrous-Belbachir, O. Phaseolus vulgaris L. seedlings exposed to prometryn herbicide contaminated soil trigger an oxidative stress response. J. Agric. Food Chem. 2016, 64, 3150–3160. [Google Scholar] [CrossRef]
- Yang, G.; Wang, Y.; Xia, D.; Gao, C.; Wang, C.; Yang, C. Overexpression of a GST gene (ThGSTZ1) from Tamarix hispida improves drought and salinity tolerance by enhancing the ability to scavenge reactive oxygen species. Plant Cell Tissue Organ Cult. 2014, 117, 99–112. [Google Scholar] [CrossRef]
- Nandula, V.K.; Riechers, D.E.; Ferhatoglu, Y.; Barrett, M.; Duke, S.O.; Dayan, F.E.; Goldberg-Cavalleri, A.; Tétard-Jones, C.; Wortley, D.J.; Onkokesung, N. Herbicide Metabolism: Crop Selectivity, Bioactivation, Weed Resistance, and Regulation. Weed Sci. 2019, 67, 149–175. [Google Scholar] [CrossRef]
- Shergill, L.S.; Bish, M.D.; Jugulam, M.; Bradley, K.W. Molecular and physiological characterization of six-way resistance in an Amaranthus tuberculatus var. rudis biotype from Missouri. Pest Manag. Sci. 2018, 74, 2688–2698. [Google Scholar] [CrossRef] [PubMed]
- Cummins, I.; Wortley, D.J.; Sabbadin, F.; He, Z.; Coxon, C.R.; Straker, H.E.; Sellars, J.D.; Knight, K.; Edwards, L.; Hughes, D. Key role for a glutathione transferase in multiple-herbicide resistance in grass weeds. Proc. Natl. Acad. Sci. USA 2013, 110, 5812–5817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGonigle, B.; Keeler, S.J.; Lau, S.M.; Koeppe, M.K.; O’Keefe, D.P. A genomics approach to the comprehensive analysis of the glutathione S-transferase gene family in soybean and maize. Plant Physiol. 2000, 124, 1105–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, D.P.; Lapthorn, A.; Edwards, R. Plant glutathione transferases. Genome Biol. 2002, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Edwards, R.; Dixon, D.P.; Walbot, V. Plant glutathione S-transferases: Enzymes with multiple functions in sickness and in health. Trends Plant Sci. 2000, 5, 193–198. [Google Scholar] [CrossRef]
- Monticolo, F.; Colantuono, C.; Chiusano, M.L. Shaping the evolutionary tree of green plants: Evidence from the GST family. Sci. Rep. 2017, 7, 14363. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.J.; Han, X.M.; Ren, L.L.; Yang, H.L.; Zeng, Q.Y. Functional divergence of the glutathione S-transferase supergene family in Physcomitrella patens reveals complex patterns of large gene family evolution in land plants. Plant Physiol. 2013, 161, 773–786. [Google Scholar] [CrossRef] [Green Version]
- Munyampundu, J.P.; Xu, Y.P.; Cai, X.Z. Phi class of glutathione S-transferase gene superfamily widely exists in nonplant taxonomic groups. Evol. Bioinform. 2016, 12, EBO-S35909. [Google Scholar] [CrossRef] [Green Version]
- Frova, C. Glutathione transferases in the genomics era: New insights and perspectives. Biomol. Eng. 2006, 23, 149–169. [Google Scholar] [CrossRef]
- Soranzo, N.; Gorla, M.S.; Mizzi, L.; De Toma, G.; Frova, C. Organisation and structural evolution of the rice glutathione S-transferase gene family. Mol. Genet. Genom. 2004, 271, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Sylvestre-Gonon, E.; Law, S.R.; Schwartz, M.; Robe, K.; Keech, O.; Didierjean, C.; Dubos, C.; Rouhier, N.; Hecker, A. Functional, Structural and Biochemical Features of Plant Serinyl-Glutathione Transferases. Front. Plant Sci. 2019, 10, 608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Gao, Q.; Xu, L.; Pang, S.; Liu, Z.; Wang, C.; Tan, W. Characterization of glutathione S-transferases in the detoxification of metolachlor in two maize cultivars of differing herbicide tolerance. Pestic. Biochem. Phy. 2017, 143, 265–271. [Google Scholar] [CrossRef]
- Karavangeli, M.; Labrou, N.E.; Clonis, Y.D.; Tsaftaris, A. Development of transgenic tobacco plants overexpressing maize glutathione S-transferase I for chloroacetanilide herbicides phytoremediation. Biomol. Eng. 2005, 22, 121–128. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Motif | Width | Best Possible Match |
|---|---|---|
| 1 | 23 | NKSELLLRSNPVHKKVPVLIHGG |
| 2 | 29 | GVWTSPFVIRVRIVLNLKGLAYEYVEEDL |
| 3 | 35 | GKPFFGGDSVGYVDVVLGGLLGWVRASEELHGVRP |
| 4 | 21 | KPVCESQIIVQYIDEAFAGTG |
| 5 | 21 | LPADPYERAVARFWAAYIDDK |
| 6 | 29 | IDAARTPLLAAWMERFCELDAAKAVLQDV |
| 7 | 80 | DWKRLYSNTKTNFREVAIKGFWDMYDPEGYSLWFCDYKYNDENTVSFVTMNKVGGFLQRMDLCRKYAFGKMLVVGSEPPF |
| 8 | 39 | DPAVIAENEDKLKQVLDVYDEILSKNEYLAGDEFTLADL |
| 9 | 40 | SSDRGRKLFTARKHVARWYDKISTRDSWRQVIKMQREHPG |
| 10 | 29 | VFRGRTAEEMAEAARQAVAALETLEQAFR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Zhao, D.; Xu, X.; Yuan, L.; Zhao, B.; Li, B.; Guo, X.; Wang, G. Role of ZmGST Gene Family Involved in Nicosulfuron Stress Tolerance Revealed by Genomic and Transcriptomic Analyses. Agronomy 2022, 12, 2598. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12112598
Liu X, Zhao D, Xu X, Yuan L, Zhao B, Li B, Guo X, Wang G. Role of ZmGST Gene Family Involved in Nicosulfuron Stress Tolerance Revealed by Genomic and Transcriptomic Analyses. Agronomy. 2022; 12(11):2598. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12112598
Chicago/Turabian StyleLiu, Xiaomin, Dan Zhao, Xian Xu, Libing Yuan, Bochui Zhao, Binghua Li, Xinli Guo, and Guiqi Wang. 2022. "Role of ZmGST Gene Family Involved in Nicosulfuron Stress Tolerance Revealed by Genomic and Transcriptomic Analyses" Agronomy 12, no. 11: 2598. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12112598