
Differences in the Mode of Action of Florpyrauxifen-Benzyl between Barnyardgrass and Yerbadetajo
1
Eco-Environmental Protection Research Institute, Shanghai Academy of Agricultural Sciences, Shanghai 201403, China
2
College of Plant Protection, Nanjing Agricultural University, Nanjing 210095, China
*
Author to whom correspondence should be addressed.
Agronomy 2022, 12(11), 2656; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12112656
Submission received: 21 September 2022
/
Revised: 19 October 2022
/
Accepted: 25 October 2022
/
Published: 27 October 2022
(This article belongs to the Special Issue Herbicides Toxicology and Weeds Herbicide-Resistant Mechanism)
Abstract
:Barnyardgrass (a monocotyledon) and yerbadetajo (a dicotyledon) are the most troublesome weeds in rice fields in China. The synthetic auxin herbicide florpyrauxifen-benzyl can effectively control both weeds. The objective of this research was to clarify modes of action of florpyrauxifen-benzyl in barnyardgrass and yerbadetajo. Our results showed that yerbadetajo was more sensitive to florpyrauxifen-benzyl than barnyardgrass: the 50% growth rate inhibition in barnyardgrass and yerbadetajo was 4.14 and 0.38 g a.i. ha−1, respectively. Florpyrauxifen-benzyl induced the ethylene precursor 1-aminocyclopropane-1-carboxylic acid (ACC) and abscisic acid (ABA) in yerbadetajo within 24 h, while in barnyardgrass, the highest production occurred at 12 h and decreased at 24 h. ACC and ABA levels in yerbadetajo at 24 h of treatment were significantly higher than those in barnyardgrass at any time. There were less differentially expressed genes related to ethylene and ABA synthesis in barnyardgrass than in yerbadetajo. There were five genes induced to increase by florpyrauxifen-benzyl in barnyardgrass and eleven genes in yerbadetajo. More ACC oxidase genes (ACO) were induced in barnyardgrass and more ACC synthesis genes (ACS) in yerbadetajo, especially three ACS3 (>30 fold). We speculated that differences in gene expression caused differences in ethylene and ABA production, leading to differences in phytotoxicity.
Keywords:
florpyrauxifen-benzyl; barnyardgrass; yerbadetajo; mode of action; ethylene; abscisic acid1. Introduction
Rice (Oryza sativa) feeds more people worldwide than any other crop and is now widely cultivated in 95 countries and regions across the world [1]. Rice has high nutritional value that can provide the human body with approximately 27% dietary energy, 20% protein, and 3% fat; it also contains zinc, riboflavin, thiamine, and niacin, which can only be obtained through dietary intake [2]. The yield and quality of rice are affected by many factors, including water, nutrients, temperature, and weeds [3]. In fact, rice has been exposed to the recurrent invasion of weeds since its cultivation, becoming the most critical obstacle in rice production. In China, rice yield loss caused by weeds can exceed 10% every year, and can even lead to a reduction of more than 50% when weeds occur seriously [4].
The genus Echinochloa (barnyardgrass) is among the most pernicious and widely distributed monocotyledonous weeds, including more than 250 annual or perennial species, most of which interfere with crop growth [5,6]. As a C4 photosynthetic plant, barnyardgrass, has strong environmental adaptability and obvious advantages in growth, stress resistance, and competitiveness under living conditions [7,8,9]. Moreover, barnyardgrass and rice are very similar in morphological characteristics, growth and development cycles, physiological and biochemical metabolism, and ecological environment requirements [10,11], making barnyardgrass the most successful competitor of rice (Supplementary Materials Figure S1a). According to different field management levels and models, barnyardgrass alone can cause rice loss of 21–79% [5,12,13]. Barnyardgrass occurs in China’s major rice-producing areas [14,15,16,17]. Another important weed species in rice fields is Eclipta prostata (yerbadetajo) [18,19,20]. Yerbadetajo is an annual dicotyledonous grass belonging to the genus Eclipta in the Compositae family. Yerbadetajo is widely distributed in tropical, subtropical, and temperate regions and is resistant to humidity and drought, with strong environmental adaptability [21,22,23]. The peak of seed germination of yerbadetajo occurs from mid-June to early July, posing a serious threat to rice production in the Yangtze River Basin in China [22], and the frequency of occurrence in some areas is as high as 100% [24]. Yerbadetajo often dominates the growth space of rice as shown in Supplementary Materials Figure S1b.
Presently, weed management in China is mainly dependent on chemical herbicides. However, herbicide resistance in barnyardgrass is widespread, and resistance management is a challenging task. Barnyardgrass has evolved resistance to a variety of herbicides, such as butachlor [25], quinclorac [14], penoxsulam [17,26], bispyribac-sodium [27] and metamifop [28], in paddy fields in China. Several herbicides, such as pyrazosulfuron-ethyl, MCPA, and bentazone, are usually applied to adequately control yerbadetajo. An arylpicolinate synthetic auxin herbicide (Group IV), florpyrauxifen-benzyl [benzyl 4-amino-3-chloro-6-(4-chloro-2-fluoro-3 methoxyphenyl)-5-fluoropicolinate; florpyrauxifen-benzyl], was commercialized to selectively control weeds in paddy fields in China in 2017 under the trade name Rinskor® (Corteva Agriscience (Wilmington, DE, USA)) [29]. It is used as a post-emergence herbicide to control most annual weeds in paddy fields at a dosage of 18–36 g a.i. ha−1 [30] and can provide a broad spectrum of weed control, even weeds that have been proven to be resistant to other herbicides, such as quinclorac-resistant E. crus-galli [31,32] or glyphosate-resistant E. colona [33]. Florpyrauxifen-benzyl may control barnyardgrass and a variety of dicotyledonous weeds, including yerbadetajo, with a single application. In addition, florpyrauxifen-benzyl may replace the herbicides, such as penoxulam and quinclorac, that have been used for a long time and have evolved resistance in barnyardgrass [14,17,26]. Florpyrauxifen-benzyl has the potential to become the main post-emergence herbicides to manage herbicide-resistant weeds in paddy fields in China [29].
Florpyrauxifen-benzyl is a new synthetic auxin herbicide [34,35,36]. The modes of action of many synthetic auxin herbicides have been well studied. It is generally believed that this kind of herbicide causes phytotoxicity mainly through two modes of action [37]. First, synthetic auxin herbicides inducted ethylene that excessively accumulates in sensitive plants. The most typical case is ethylene accumulation by quinclorac in sensitive plants [14,38,39,40]. Cyanide, a by-product of ethylene biosynthesis, is also induced to accumulate by quinclorac, which is an important cause of plant poisoning [41,42,43]. In addition, abscisic acid (ABA) accumulation is over-induced by indoleacetic acid and auxin-type herbicides in sensitive plants [44,45]. Excessive accumulation of ABA may interfere with photosynthesis [45,46] and eventually lead to a burst of reactive oxygen species (ROS) and damage plants [37]. Our previous research found that the mode of action of quinclorac in barnyardgrass includes the above two modes. Furthermore, we also explained the molecular mechanism underlying the changes in the expression of 1-aminocyclopropane-1-carboxylic acid synthase (ACS), 1-aminocyclopropane-1-carboxylic acid oxidase (ACO), and 9-cis-epoxycarotenoid dioxygenase (NCED) genes, which resulted in ethylene and ABA accumulation [38,45,46]. Since florpyrauxifen-benzyl has been used for a short time, there are few studies that investigated its mode of action. A few studies have focused on the biochemical reactions of barnyardgrass caused by florpyrauxifen-benzyl [29]. Our previous study found that florpyrauxifen-benzyl had an excellent control effect on yerbadetajo. However, the molecular mechanism of florpyrauxifen-benzyl in controlling these two weed species and the difference between them has not yet been reported.
In the present study, the representative monocotyledon weed, barnyardgrass, and the representative dicotyledon weed, yerbadetajo, in paddy fields in China were selected as research materials. First, we determined the differences in sensitivity to florpyrauxifen-benzyl between the two weeds. Next, we examined the biosynthesis of ethylene and ABA after florpyrauxifen-benzyl treatment. Finally, we identified the related functional pathways and genes using high-throughput sequencing and quantitative real-time PCR (RT-qPCR). The objective of this study was to clarify the difference in the mode of action of florpyrauxifen-benzyl in barnyardgrass and yerbadetajo and to provide a theoretical basis for the innovation of herbicide application technology and delaying resistance evolution.
2. Materials and Methods
2.1. Chemical
Florpyrauxifen-benzyl, 3% EC (Rinskor®) was obtained from Corteva Agriscience (Wilmington, DE, USA).
2.2. Plant Material
Barnyardgrass and yerbadetajo seeds were collected from the Jinshan District of Shanghai City on October 2020. The weeds were cultivated at 30/25 °C under 14 h of light (20,000 lx) and 10 h of darkness each day in a plant culture room and watered every three days to the 3–4 leaf stage.
2.3. Whole-Plant Bioassay to Determine Herbicide Sensitivity
Whole-plant bioassays were used to determine the susceptibility of barnyardgrass and yerbadetajo to florpyrauxifen-benzyl. The soil was of a middle loam type from farmland in the suburbs of Shanghai, China, where herbicides have not been used for more than five years. The soil was placed in plastic pots (7 × 7 × 7 cm). Water was added until saturation. Twenty seeds were sown in each plastic cup, covered with a fine layer of soil, and cultivated to the 2 leaf stage as described in Section 2.2. After thinning, 15 seedlings of barnyardgrass and ten seedlings of yerbadetajo were selected for continued cultivation (the others were removed) to the 3–4 leaf stage before being treated with a stem and leaf spray. The control plants were sprayed with the same amount of water as the treated plants. The dosages of florpyrauxifen-benzyl were set at 1.125, 2.25, 4.5, 9, 18, and 36 g a.i. ha−1 for barnyardgrass and 0.28125, 0.5625, 1.125, 2.25, 4.5, 9, and 18 g a.i. ha−1 for yerbadetajo. The stems and leaves were sprayed using a 3WP-2000 walking-type spraying system (Nanjing Agricultural Mechanization Research Institute of the Ministry of Agriculture, China). Each treatment was performed with 30 mL of liquid (450 L ha−1 water) using a fan-shaped nozzle. After the liquid on the stems and leaves had dried, the seedlings were placed in a greenhouse for cultivation. After 21 d, the grass was cut and weighed. The inhibition rate of the aboveground fresh weight was calculated. The experiment contained four biological replicates, and the entire experiment was repeated twice. The effective rate of each herbicide causing 50% inhibition in plant height (GR50) was determined using the four-parameter logistic function with the “drc” add-on package [47] in the R 3.1.3 Language and Environment for Statistical Computing [48]. The model was defined as:
The parameter e is also denoted GR50 and is the dose producing a response half-way between the upper limit, d, and the lower limit, c. The parameter b denotes the relative slope around e.
2.4. Liquid Chromatography–Mass Spectrometry Method to Detect 1-Aminocyclopropane-1-Carboxylic Acid (ACC) Content
Entire barnyardgrass and yerbadetajo plants at the 2.5–3.5 leaf stage were sprayed with 18 g a.i. ha−1 of florpyrauxifen-benzyl, as described in Section 2.3. After 0 (untreated control), 3, 6, 12, and 24 h, shoots were collected and immediately frozen in liquid nitrogen for ACC content determination. A sample of each plant was ground into powder in liquid nitrogen, which was accurately weighed (0.2 g) and transferred to a test tube. ddH2O, which was precooled to 4 °C, was then added to the test tube and allowed to stand for 2 h for extraction. The supernatant was retained after centrifuging twice at 10,000× g for 5 min at 4 °C, after which the two extracted supernatants were combined, passed through the McX column, and washed with 5 mL of water. The eluent was analyzed using high-performance liquid chromatography–tandem mass spectrometry (HPLC–MS/MS) after passing through a 0.22 µM filter membrane.
For the HPLC analysis using a reversed-phase chromatography column (Poroshell 120 SB-C18, 2.1 × 150, 2.7 µm), the injection volume was 2 µL, and the column temperature was set to 35 °C. The mobile phase used was A:B = acetonitrile: (water/0.1% formic acid) = 3:7. The elution gradients were equal. The electrospray ionization (ESI) source and positive ion modes (+4500 V) were used for the spectrometry analysis. The temperature was set to 350 °C, the curtain air was set to 15 PSI, the atomization air pressure was set to 65 PSI, and the auxiliary air pressure was set to 70 PSI. The other testing conditions are listed in Table 1.
Each experimental treatment was performed in triplicate, and the experiment was conducted twice. Significant differences in the absolute ACC content of barnyardgrass and yerbadetajo were compared using Duncan’s multiple range test (p < 0.05). Analysis of variance (ANOVA) was performed using the SPSS Statistics (for Windows, Version 20.0. Armonk, NY, USA: IBM Corp.).
2.5. Liquid Chromatography–Mass Spectrometry Method to Detect ABA Content
Entire barnyardgrass and yerbadetajo plants at the 2.5–3.5 leaf stage were sprayed with 18 g a.i. ha−1 of florpyrauxifen-benzyl, as described in Section 2.3. After 0 (untreated control), 3, 6, 12, and 24 h, shoots were collected and immediately frozen in liquid nitrogen for ABA content determination. A sample of each plant was ground into a powder in liquid nitrogen, which was accurately weighed (0.5 g) and transferred to a test tube. Ten times the volume of acetonitrile solution and 4 μL of the internal standard mother liquor were added. The solution was extracted at 4 °C overnight, centrifuged at 12,000× g for 5 min, and the supernatant was collected. Five times the volume of acetonitrile solution was added to the precipitate for extraction, and the supernatants obtained from the two extractions were combined. Then, 15–40 mg of C18 filler was added to the supernatant. After shaking violently for 30 s, the mixture was centrifuged at 10,000× g for 5 min, and the supernatant was collected. The supernatant was dried under nitrogen and added to 400 μL of methanol. The solution was then passed through a 0.22 μM organic phase filter membrane and stored at −20 °C for detection.
For the HPLC analysis using a reversed-phase chromatographic column (Poroshell 120 SB-C18, 2.1 × 150, 2.7 µm), the injection volume was 2 µL, and the column temperature was set to 30 °C. The mobile phase was A:B = (methanol/0.1% formic acid): (water/0.1% formic acid). The elution gradients are listed in Table 2.
For the spectrometry analysis, the ESI and positive and negative ion modes (+4500 V, −4000 V) were used. The scan type was MRM. The temperature was set to 400 °C, the curtain air was set to 15 PSI, the atomization air pressure was set to 65 PSI, and the auxiliary air pressure was set to 70 PSI. The other testing conditions are listed in Table 3.
Each experimental treatment was performed in triplicate, and the experiment was conducted twice. Significant differences in the absolute ABA content of barnyardgrass and yerbadetajo were compared using Duncan’s multiple range test (p < 0.05). ANOVA was performed using the SPSS Statistics (for Windows, Version 20.0. Armonk, NY, USA: IBM Corp.).
2.6. High-Throughput Sequencing to Screen for Differentially Expressed Genes (DEGs)
Entire barnyardgrass and yerbadetajo plants at the 2.5–3.5 leaf stage were sprayed with 18 g ai ha−1 of florpyrauxifen-benzyl, as described in Section 2.3. After 3, 6, 12, and 24 h, shoots were collected, and samples not treated with florpyrauxifen-benzyl were harvested and immediately frozen in liquid nitrogen for RNA extraction. A library of barnyardgrass and yerbadetajo treated with florpyrauxifen-benzyl was constructed with the assistance of Guangzhou Gene Denove Biotechnology Co., Ltd. Clustering of index-coded samples was performed on a cBot Cluster Generation System using a TruSeq PE Cluster Kit v3-cBot-HS (Illumina), according to the manufacturer’s instructions. After cluster generation, libraries were sequenced on an Illumina HiSeq platform, and paired-end reads were generated. Gene function was annotated based on the following databases: Nr (NCBI non-redundant protein sequences), Nt (NCBI non-redundant nucleotide sequences), Pfam (Protein family), KOG/COG (Clusters of Orthologous Groups of proteins), Swiss-Prot (a manually annotated and reviewed protein sequence database), KO (KEGG Ortholog database), and GO (Gene Ontology). Based on the gene function annotation and differential gene expression analysis, DEGs associated with ethylene and ABA biosynthesis after florpyrauxifen-benzyl treatment were screened. Repetitive fragments, fragments whose length is less than 150 bp, and fragments whose fpkm value is less than 1 at all-time points sequenced were eliminated. The screened genes were further confirmed by sequence alignment using the NCBI website (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/, accessed on 22 June 2022).
2.7. Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR) to Verify DEGs
Based on the transcriptional sequencing data of barnyardgrass (GenBank number: PRJNA828271) and yerbadetajo (GenBank number: PRJNA828110), the genes ACS, ACO, and NCED were screened using gene function annotation. The screened gene sequences were compared using the NCBI BLAST function to confirm the names and functions of these genes. RT-qPCR primers for the target genes were designed using an online tool, Primer3 (v. 0.4.0) (http://bioinfo.ut.ee/primer3-0.4.0/, accessed on 8 August 2022). EcActin (GenBank accession number: HQ395760) in barnyardgrass [49] and 18SrRNA gene (Supplementary Materials Table S1) were used as internal reference genes. The primers used are listed in Supplementary Materials Table S1. The dissociation profiles showed that the amplification products (Supplementary Materials Table S1) of each pair of primers have a single peak, and the amplification efficiency values were between 90–110%. Barnyardgrass and yerbadetajo samples were used to detect ACS, ACO, and NCED mRNA levels at 0 (untreated control) and 12 h after florpyrauxifen-benzyl treatment. Total RNA was extracted using the RNAsimple Total RNA Kit (TIANGEN, Beijing, China), according to the manufacturer’s instructions. Then, RNA samples were reverse transcribed to cDNA using the PrimeScript™ RT Reagent Kit with gDNA Eraser (TaKaRa, Otsu, Japan), and the cDNA samples were diluted to a uniform value. The reaction system and procedure were performed according to the manufacturer’s instructions for the TAKARA TB Green Premix Ex Taq II reagent kit. RT-qPCR was performed using the 2−ΔΔCt method based on two repeated techniques and three biological replicates.
All six replicates were analyzed significant difference. To compare differences in gene expression between control (0 h) and treatment (12 h), Student’s t-test (p < 0.05) was performed. Two threshold values, a fold change > 2 and a significant result in the t-test (p < 0.05), were used to determine the up-regulation or down-regulation of genes [50] induced by florpyrauxifen-benzyl.
3. Results
3.1. Sensitivity to Florpyrauxifen-Benzyl
The sensitivity of barnyardgrass and yerbadetajo to florpyrauxifen-benzyl was tested using a whole-plant bioassay (Figure 1). Florpyrauxifen-benzyl caused twisting of the stem bases of the two weeds, and the leaves lost water until they died. The gradient dose of florpyrauxifen-benzyl resulted in the fresh weight reduction in barnyardgrass and yerbadetajo by 12–91% and 37–96%, respectively. The GR50 of florpyrauxifen-benzyl to barnyardgrass was 4.14 ± 0.15 g a.i. ha−1, respectively. The GR50 of florpyrauxifen-benzyl to yerbadetajo was 0.38 ± 0.01 g a.i. ha−1, respectively. The ratio of GR50 of florpyrauxifen-benzyl to barnyardgrass and to yerbadetajo was 10.9.
3.2. ACC Production
The content of the ethylene precursor ACC was regularly monitored over 24 h of florpyrauxifen-benzyl treatment (Figure 2). The results showed that the ACC content in barnyardgrass increased sharply during 12 h after florpyrauxifen-benzyl treatment. The ACC content in untreated barnyardgrass was 32.75 ± 2.78 ng/g but reached 239.02 ± 26.69, 774.56 ± 98.12, and 3094.11 ± 282.03 ng/g at 3, 6, and 12 h, respectively, after florpyrauxifen-benzyl treatment, which was 7.3−94.5 fold that of the control; after florpyrauxifen-benzyl treatment for 24 h, the ACC content decreased, but it was still 2270.58 ± 271.90 ng/g and 69.3 fold that of the untreated control. The results also showed that the ACC content in yerbadetajo increased sharply 24 h after florpyrauxifen-benzyl treatment. The ACC content in untreated yerbadetajo was 5.87 ± 0.13 ng/g but reached 382.98 ± 3.27, 737.21 ± 3.52, 1938.52 ± 11.89, and 4136.60 ± 8.50 ng/g at 3, 6, 12, and 24 h, respectively, after florpyrauxifen-benzyl treatment, which was 65.2−704.7 fold that of the control. The ACC content in yerbadetajo rose continually within 24 h. It was significantly higher than the detection value at any time in both species at 24 h after florpyrauxifen-benzyl treatment (p < 0.05).
3.3. ABA Production
ABA content was regularly monitored within 24 h after florpyrauxifen-benzyl treatment (Figure 3). The results showed that ABA content in barnyardgrass increased 6 and 12 h after florpyrauxifen-benzyl treatment. The ABA content in untreated barnyardgrass was 10.20 ± 0.53 ng/g but was 46.16 ± 2.43 and 67.45 ± 3.91 ng/g at 6 and 12 h, respectively, after florpyrauxifen-benzyl treatment, which was 4.5 and 6.6 fold, respectively, that of the control; after florpyrauxifen-benzyl treatment for 24 h, the ABA content decreased to 44.49 ± 2.10 ng/g, which was still 4.4 fold that of the control. The results also showed that ABA content in yerbadetajo increased sharply 24 h after florpyrauxifen-benzyl treatment. The ABA content in untreated yerbadetajo was 4.77 ± 0.03 ng/g but reached 21.19 ± 0.35, 30.24 ± 0.46, 80.29 ± 1.12, and 176.03 ± 2.69 ng/g at 3, 6, 12, and 24 h, respectively, after florpyrauxifen-benzyl treatment, which was 4.4−36.9 fold that of the control. The ABA content in yerbadetajo rose continually within 24 h and was significantly higher than the detection value at any time point in both species at 24 h after florpyrauxifen-benzyl treatment (p < 0.05).
3.4. DEGs Related to ACC and ABA Biosyntheses
Transcriptome sequencing data showed that the expression levels of many genes differed after florpyrauxifen-benzyl treatment. There were 12729–16339 DEGs (fold change > 2; FDR ≤ 0.05) and 6097–11204 DEGs (fold change > 2; FDR ≤ 0.05) in barnyardgrass and yerbadetajo, respectively (Table 4). In barnyardgrass, the number of up-regulated genes was much higher than that of down-regulated genes, which was not observed in yerbadetajo. Based on the KEGG classification, most DEGs of the two weeds were enriched in metabolic pathways. In the next level category, these DEGs were mainly enriched in global and overview maps (metabolism), carbohydrate metabolism (metabolism), and translation (genetic information processing) in barnyardgrass and global and overview maps (metabolism), signal transduction (environmental information processing), and carbohydrate metabolism (metabolism) in yerbadetajo (Figure 4).
Based on functional annotations and DEG analysis from transcriptome sequencing, we screened nine genes among the unigenes in barnyardgrass after further sequence alignment on the NCBI website (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/, accessed on 22 June 2022). Amplification using the primers in Supplementary Materials Table S1, sequencing, and alignment analysis revealed that all nine genes were present in barnyardgrass. Six of these genes encoded ACO, two encoded ACS, and one encoded NCED (Table 5). Using the same method, amplification using the primers listed in Supplementary Materials Table S1, sequencing, and alignment analysis revealed that all 16 genes were present in yerbadetajo. Six of these genes encoded ACO, seven encoded ACS, and three encoded NCED (Table 5).
3.5. The Expression Profiles of ACS, ACO, and NCED
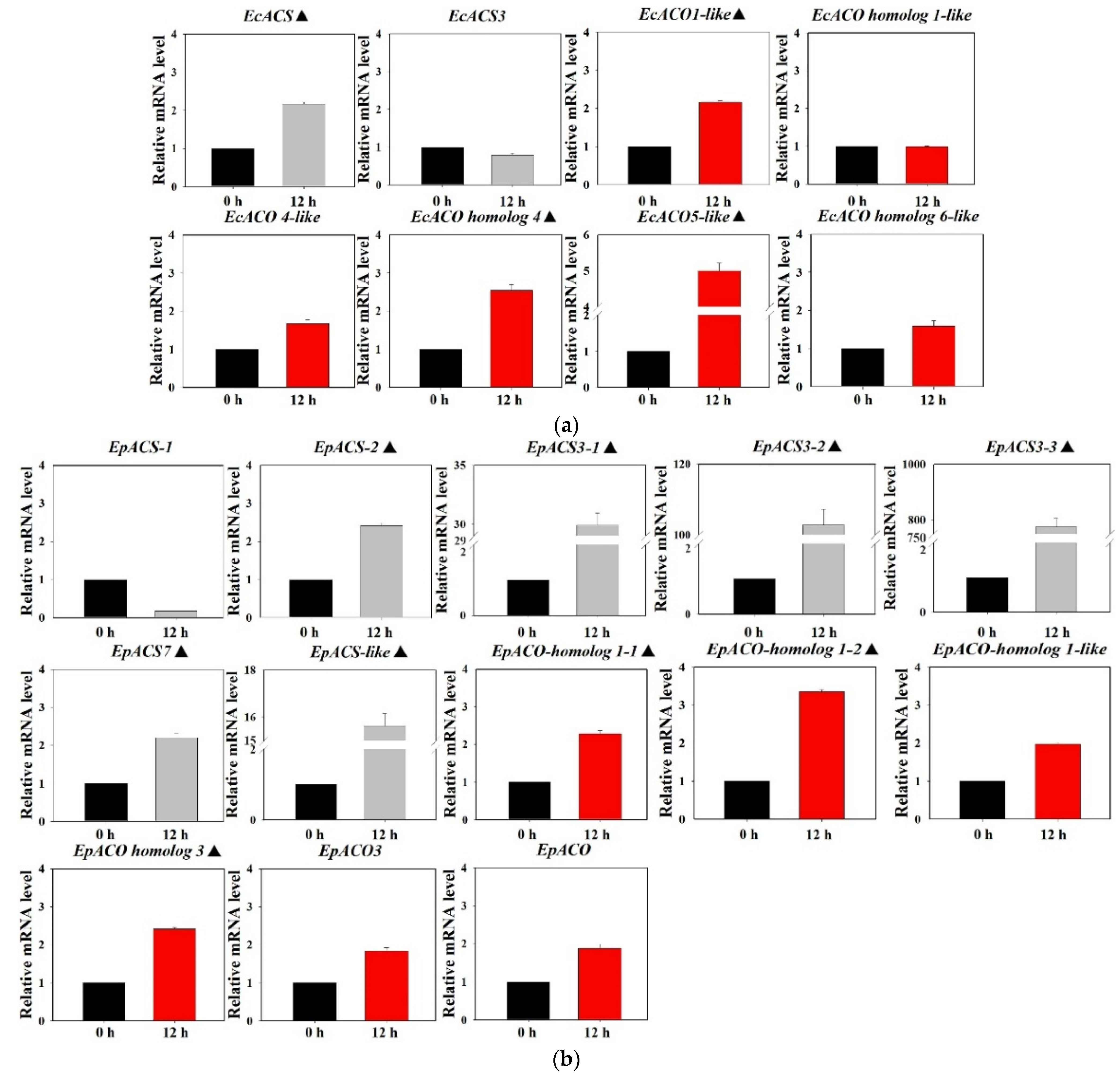
The relative levels of ACS and ACO mRNA in the shoots of barnyardgrass and yerbadetajo at the seedling stage was assessed by RT-qPCR (Figure 5). In barnyardgrass, the expression of EcACS, EcACO1-like, and EcACO homolog 4 increased 2 fold at 12 h after florpyrauxifen-benzoyl treatment and the expression of EcACO5-like increased by approximately 5 fold at 12 h after florpyrauxifen-benzoyl treatment; the expression of these four genes were significantly higher than that of the untreated control (0 h) (p < 0.05) (Figure 5a). In yerbadetajo, there were nine genes whose expression changes were more than 2 fold, which were significantly higher than that in the untreated control (p < 0.05); among these genes, the expression of EpACS3-2 and EpACS3-3 was more than 100 fold and the expression of EpACS3-1 and EpACS-like was more than 10 fold at 12 h after florpyrauxifen-benzoyl treatment compared to the untreated control (Figure 5b). The mRNA level of one gene, EpACS-1, was detected to decrease at 12 h after florpyrauxifen-benzoyl treatment, which was inconsistent with the expression trend of other genes.
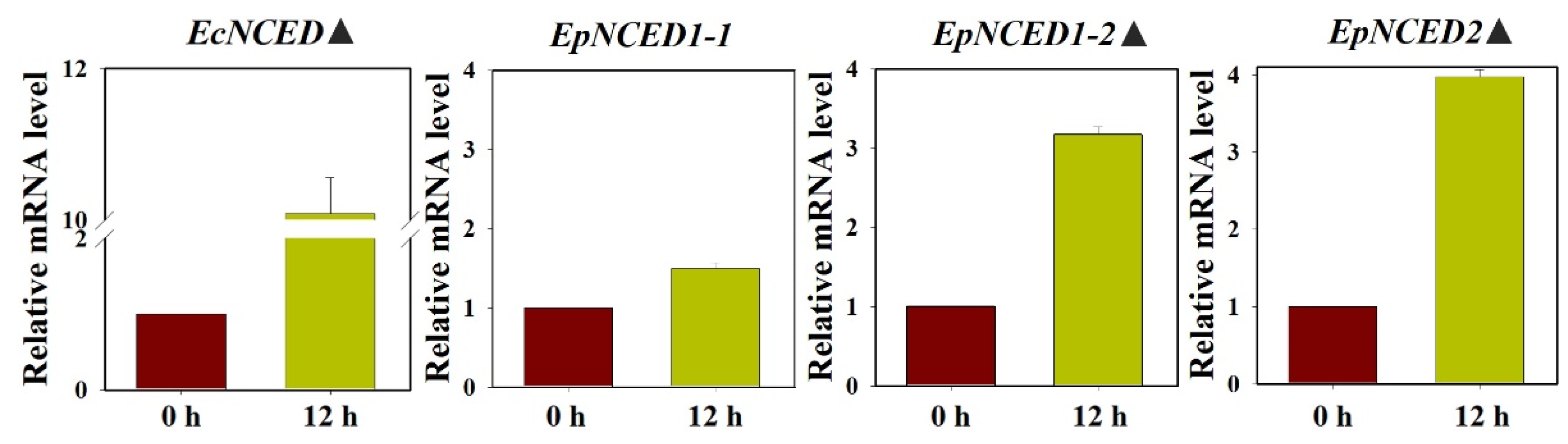
The relative levels of NCED mRNA in the shoots of barnyardgrass and yerbadetajo at the seedling stage was also assessed by RT-qPCR (Figure 6). The expression of EcNCED was more than 10 fold at 12 h after florpyrauxifen-benzoyl treatment compared to the control (0 h) in barnyardgrass. While two of three EpNCED, EpNCED1-2 and EpNCED2, were increased by approximately 3 fold at 12 h after florpyrauxifen-benzoyl treatment compared to the control (0 h) in yerbadetajo.
4. Discussion
Rampant weeds in rice fields have become the most critical factor restricting the yield and quality of rice [3,4,12,13]. In China, rice farmers prefer herbicides to other methods to control weeds because the simplicity of operation in agricultural production is their first choice. The development and application of florpyrauxifen-benzyl filled the gap after barnyardgrass evolved resistance to metamifop, bispyribac-sodium, quinclorac, and penoxsulam in China [14,17,26,27,28]. Similar to quinclorac, florpyrauxifen-benzyl is a synthetic auxin herbicide that controls many dicotyledonous weeds and the troublesome monocotyledonous weed, barnyardgrass [29,34,35,36]. This differs from other auxin herbicides that can only control dicotyledonous weeds [30]. This study showed that yerbadetajo was far more sensitive to florpyrauxifen-benzyl than to barnyardgrass, with a GR50 ratio of 10.9 (Figure 1). It is necessary to clarify the mode of action of florpyrauxifen-benzyl in the two weeds and the reasons for these differences in sensitivity to florpyrauxifen-benzyl.
Synthetic auxin herbicides mimic the effect of plant auxin and specifically initiate ethylene and ABA biosynthesis pathways after binding to the receptor TIR/AFB and the co-receptor IAA/AUX [37,44,51,52,53]. This view was confirmed in the mode of action of quinclorac on barnyardgrass [38] and other synthetic auxins such as florpyrauxifen-benzyl on Galium aparine [54]. Ethylene is a hormone that can accelerate plant senescence [55,56,57,58,59], a precursor of ACC in the biosynthesis process [60]. The ACC content can be used to characterize the ethylene content, which is positively correlated to the amount of ethylene [14]. Ethylene accumulation also results in the excessive accumulation of cyanide, a by-product of phytotoxicity [43,61]. ABA can lead to stomatal closure and interfere with photosynthesis, causing excessive accumulation of ROS, which also have toxic effects on plants [37,62]. Previous studies have shown that ethylene accumulation mainly occurs in monocotyledons, and ABA accumulation mainly occurs in dicotyledons after auxin analog treatment [63]. However, our previous studies confirmed that the accumulation of ethylene and ABA could occur in monocotyledons (barnyardgrass) after quinclorac treatment [38,45]. This study demonstrated that florpyrauxifen-benzyl caused excessive production of ethylene and excessive accumulation of ABA in barnyardgrass (Figure 2 and Figure 3). The same phenomenon was also found in yerbadetajo, but the degree was sharper than that in barnyardgrass and this induction lasted longer (Figure 2 and Figure 3), which undoubtedly caused more serious damage to yerbadetajo. The content of ACC and ABA of barnyardgrass and yerbadetajo after 24 h of florpyrauxen-benzyl treatment was not monitored continuously, because the two weeds of florpyrauxen-benzyl treatment showed severe wilting and chlorosis after 24 h.
Changes in the tertiary structure or expression level of herbicide targets at herbicide binding sites or functional proteins can lead to resistance [38,64,65]. Ethylene biosynthesis is controlled mainly by two gene families, the ACS and ACO gene families, which are expressed in Arabidopsis, tomato, melon, and deep-water rice under synthetic auxin treatment or stress [66,67,68,69,70,71,72,73]. NCED is a key enzyme in ABA biosynthesis [74,75]. Our previous studies confirmed that ACS, ACO, and NCED could be induced by quinclorac in barnyardgrass [38,45]. In particular, the expression of Ec-ACS7 could be strongly induced by quinclorac, which was considered to be the molecular basis for the mode of action of quinclorac in barnyardgrass [38]. In the present study, high-throughput sequencing and RT-qPCR verification all showed that compared with only one ACS induced by florpyrauxen-benzyl, the increase in ACO mRNA levels was more obvious (Figure 5a). At the same time, NCED in barnyardgrass was also found to be strongly induced by florpyrauxen-benzyl (Figure 6). These may be the molecular basis of excessive production of ethylene and ABA, indicating slightly differences in the mechanism of action mode of quinclorac in barnyardgrass [38,45]. Different from barnyardgrass, the increase in ACS mRNA level in yerbadetajo was more obvious, because the expression of two ACSs, ACS 3-2, and ACS 3-3, were even more than 100-fold higher compared to the control (0 h) at 12 h after florpyrauxen-benzyl treatment (Figure 5b). At the same time, mRNA levels of two NCEDs, EpNCED 1-2 and EpNCED 2, were also induced to rise. These may be the molecular basis of excessive production of ethylene and ABA by florpyrauxen-benzyl in yerbadetajo. The reason for the decrease in EcACS-1 mRNA level in yerbadetajo may be that it plays a negative role in regulation, or it may not play a role in florpyrauxen-benzyl signal transduction, just because of plant poisoning.
Yerbadetajo has received little attention previously. However, the damage of yerbadetajo to rice fields in China has become more serious, and it is difficult to control in the late growth stage. Our whole-plant bioassay results showed that the control effect of common herbicides, pyrazosulfuron-ethyl and MCPA-Na, on the yerbadetajo at the 5 leaf stage was lower than that of florpyrauxen-benzyl (data not shown). Therefore, we selected yerbadetajo as the research material of dicotyledon weeds. Based on the results of this study, it is speculated that the overexpression of three ACS3 induced by florpyrauxen-benzyl may be the key to cause yerbadetajo poisoning. This is the first time to find genes strongly expressed in yerbadetajo induced by synthetic auxin herbicide. The difference in the mode of action of florpyrauxen-benzyl in yerbadetajo compared with barnyardgrass is mainly reflected in: (1) the detection of more related overexpression genes; (2) the higher mRNA levels of ACS; (3) more ethylene and ABA production and for longer.
5. Conclusions
This study elucidated modes of action of florpyrauxifen-benzyl in barnyardgrass and yerbadetajo. Although the excessive accumulation of ACC and ABA was found in both barnyardgrass (a monocotyledon) and yerbadetajo (a dicotyledon), florpyrauxifen-benzyl caused differences in degree and response time. Different response genes and mRNA levels were also detected. However, the functions of genes with large changes in mRNA levels such as EcACO5 and EpACS3, including the EpACS-1 gene with interesting decreased expression, need further verification. Additionally, whether florpyrauxifen-benzyl directly induces changes in mRNA levels of ACS, ACO, and NCED or involves other pathways is also unclear and requires further study.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/agronomy12112656/s1, Figure S1: Barnyardgrass (a) and yerbadetajo (b) grow in rice fields; Table S1: Sequences of DEGs related to ethylene and ABA biosynthesis identified in barnyardgrass and primers for RT-qPCR.
Author Contributions
Conceptualization, Y.G.; methodology, Y.G. and J.L.; software, Y.G.; validation, Y.G. and Z.T.; formal analysis, Y.G. and Z.T.; investigation, Y.G. and Z.T.; resources, Y.G. and Z.T.; data curation, Y.G. and G.S.; writing—original draft, Y.G.; writing—review and editing, J.L., Z.T. and G.S.; visualization, Y.G. and J.L.; supervision, Z.T. and G.S.; project administration, Y.G.; funding acquisition, Y.G. and Z.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Shanghai Sailing Program, grant number [22YF1441100], and special research on supporting fields of agricultural science and technology innovation, grant number [Shengkechuang-YJ 2022(11)].
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All transcriptome data were uploaded to the NCBI (Genebank number: PRJNA828271 and PRJNA828110). Other data from this study are contained within this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bhattacharya, K.R. Introduction: Rice in historical and social perspectives. Rice Qual. Woodhead Publ. Ser. Food Sci. Technol. Nutr. Elsevier Sci. 2013, 1–25. [Google Scholar] [CrossRef]
- Philip, D.; Jayeoba, O.; Ndripaya, Y.; Fatunbi, A. Innovation Opportunities in the Rice Value Chain in Nigeria. FARA Res. Rep. 2018, 2, 1–48. [Google Scholar]
- Alibu, S.; Mamadou, F. How Does Water Stress and Nitrogen Fertilizer Affect the Growth and Yield of Upland Rice (Oryza sativa L.). J. Food Nutr. Res. 2021, 9, 215–222. [Google Scholar] [CrossRef]
- Zhang, Z.P. Development of chemical weed control and integrated weed management in China. Weed Biol. Manag. 2003, 3, 197–203. [Google Scholar] [CrossRef]
- Bajwa, A.A.; Jabran, K.; Shahid, M.; Ali, H.H.; Chauhan, B.; Ehsanullah. Eco-biology and management of Echinochloa crus-galli. Crop Prot. 2015, 75, 151–162. [Google Scholar] [CrossRef]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: www.weedscience.org (accessed on 16 May 2022).
- Gealy, D.R.; Fischer, A.J. 13C Discrimination: A Stable Isotope Method to Quantify Root Interactions between C3 Rice (Oryza sativa) and C4 Barnyardgrass (Echinochloa crus-galli) in Flooded Fields. Weed Sci. 2010, 58, 359–368. [Google Scholar] [CrossRef]
- Marambe, B.; Amarasinghe, L. Propanil-resistant barnyardgrass [Echinochloa crus-galli (L.) Beauv.] in Sri Lanka: Seedling growth under different temperatures and control. Weed Biol. Manag. 2002, 2, 194–199. [Google Scholar] [CrossRef]
- Juliano, L.M.; Casimero, M.C.; Llewellyn, R. Multiple herbicide resistance in barnyardgrass (Echinochloa crus-galli) in direct-seeded rice in the Philippines. Int. J. Pest Manag. 2010, 56, 299–307. [Google Scholar] [CrossRef]
- Gibson, K.; Fischer, A.; Foin, T.; Hill, J. Implications of delayed Echinochloa spp. germination and duration of competition for integrated weed management in water-seeded rice. Weed Res. 2002, 42, 351–358. [Google Scholar] [CrossRef]
- Clay, S.A.; Kleinjan, J.; Clay, D.E.; Forcella, F.; Batchelor, W. Growth and Fecundity of Several Weed Species in Corn and Soybean. Agron. J. 2005, 97, 294–302. [Google Scholar] [CrossRef]
- Ottis, B.V.; Talbert, R.E. Barnyardgrass (Echinochloa crus-galli) Control and Rice Density Effects on Rice Yield Components. Weed Technol. 2007, 21, 110–118. [Google Scholar] [CrossRef]
- Wilson, M.J.; Norsworthy, J.K.; Scott, R.C.; Gbur, E.E. Program Approaches to Control Herbicide-Resistant Barnyardgrass (Echinochloa crus-galli) in Midsouthern United States Rice. Weed Technol. 2014, 28, 39–46. [Google Scholar] [CrossRef]
- Xu, J.; Lv, B.; Wang, Q.; Li, J.; Dong, L. A resistance mechanism dependent upon the inhibition of ethylene biosynthesis. Pest Manag. Sci. 2013, 69, 1407–1414. [Google Scholar] [CrossRef]
- Chen, G.; Wang, Q.; Yao, Z.; Zhu, L.; Dong, L. Penoxsulam-resistant barnyardgrass (Echinochloa crus-galli) in rice fields in China. Weed Biol. Manag. 2015, 16, 16–23. [Google Scholar] [CrossRef]
- Liu, H.Y.; Han, H.T.; Zuo, H.L.; Gao, L.; Zhong, G.H. Effect of pyrazosulfuron against Echinochloa crus-galli in rice fields in South China. Guangdong Agric. Sci. 2011, 38, 3. (In Chinese) [Google Scholar]
- Liu, J.; Fang, J.; He, Z.; Li, J.; Dong, L. Target site–based resistance to penoxsulam in late watergrass (Echinochloa phyllopogon) from China. Weed Sci. 2019, 67, 380–388. [Google Scholar] [CrossRef]
- Li, D.; Li, X.; Yu, H.; Cui, H. Detection of resistance of Eclipta prostrata L. to pyrazosulfuron-ethyl in some provinces of China. Plant Prot. 2017, 196–201. (In Chinese) [Google Scholar]
- Zhao, H.Y.; Lu, Z.C.; Qiao, L.; Yang, L.M. Comparative Test of Weed Control Efficacy of Five Herbicides in Dry Direct-seeding Rice Field. Weed Sci. 2012, 48–50. (In Chinese) [Google Scholar]
- Li, R.; Luo, X.; Dong, L.; Zhao, G.; Fu, Y. Sensitivity of Eclipta prostrata populations to herbicides. Jiangsu J. Agric. Sci. 2013, 1514–1516. (In Chinese) [Google Scholar]
- Li, Y.H. Weeds of China; China Agroculture Press: Beijing, China, 1998. (In Chinese) [Google Scholar]
- Zhou, X.G.; Zhang, H. Primary Color Map of Common Weeds in Sichuan Farmland; Sichuan Publishing House of Science & Technology: Chengdu, China, 2006. (In Chinese) [Google Scholar]
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s Worst Weeds; Distribution and biology, University Press of Hawaii: Honolulu, HI, USA, 1977. [Google Scholar]
- Li, S.S.; Zhang, L.J.; Qiang, S. Weed Community Characteristics and Comprehensive Evaluation on Weeds in Paddy Fields under Labour-Saving Cultivations in Middle Jiangsu Province, China. Chin. J. Rice Sci. 2009, 23, 207–214. (In Chinese) [Google Scholar]
- Huang, B.Q.; Lin, S.X. Study on the resistance of the barnyardgrass in the paddy fields to the butachlor in China. J. South China Agric. Univ. 1993, 14, 103–108. (In Chinese) [Google Scholar]
- Fang, J.; Liu, T.; Zhang, Y.; Li, J.; Dong, L. Target site–based penoxsulam resistance in barnyardgrass (Echinochloa crus-galli) from China. Weed Sci. 2019, 67, 281–287. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Z.; Gu, T.; Dong, M.; Peng, Q.; Bai, L.; Li, Y. Quantitative proteomics reveals ecological fitness cost of multi-herbicide resistant barnyardgrass (Echinochloa crus-galli L.). J. Proteom. 2017, 150, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Zuo, P.; Ji, M.; Zang, X.; Chen, S.; Du, Y. The resistance level and acetyl co-enzyme A carboxylase activity of Echinochloa phyllopogon populations to metamifop in paddies. J. Plant Prot. 2017, 44, 1040–1045. (In Chinese) [Google Scholar]
- Wang, H.; Sun, X.; Yu, J.; Li, J.; Dong, L. The phytotoxicity mechanism of florpyrauxifen-benzyl to Echinochloa crus-galli (L.) P. Beauv and weed control effect. Pestic. Biochem. Physiol. 2021, 179, 104978. [Google Scholar] [CrossRef]
- China Pesticide Information Network. 2020. Available online: https://www.chinapesticide.org.cn/ (accessed on 16 May 2022).
- Epp, J.B.; Alexander, A.L.; Balko, T.W.; Buysse, A.M.; Brewster, W.K.; Bryan, K.; Daeuble, J.F.; Fields, S.C.; Gast, R.E.; Green, R.A.; et al. The discovery of Arylex™ active and Rinskor™ active: Two novel auxin herbicides. Bioorg. Med. Chem. 2016, 24, 362–371. [Google Scholar] [CrossRef]
- Miller, M.R.; Norsworthy, J.K.; Scott, R.C. Evaluation of Florpyrauxifen-benzyl on Herbicide-Resistant and Herbicide-Susceptible Barnyardgrass Accessions. Weed Technol. 2017, 32, 126–134. [Google Scholar] [CrossRef]
- Travlos, I.; Kanatas, P.; Tsekoura, A.; Gazoulis, I.; Papastylianou, P.; Kakabouki, I.; Antonopoulos, N. Efficacy of Different Herbicides on Echinochloa colona (L.) Link Control and the First Case of Its Glyphosate Resistance in Greece. Agronomy 2020, 10, 1056. [Google Scholar] [CrossRef]
- Lim, S.-H.; Kim, H.; Noh, T.-K.; Lim, J.-S.; Yook, M.-J.; Kim, J.-W.; Yi, J.-H.; Kim, D.-S. Baseline Sensitivity of Echinochloa crus-gall and E. oryzicola to Florpyrauxifen-Benzyl, a New Synthetic Auxin Herbicide, in Korea. Front. Plant Sci. 2021, 12, 1065. [Google Scholar] [CrossRef]
- Hwang, J.I.; Norsworthy, J.K.; González-Torralva, F.; Priess, G.L.; Barber, L.T.; Butts, T.R. Non-target-site resistance mechanism of barnyardgrass [Echinochloa crus-galli (L.) P. Beauv.] to florpyrauxifen-benzyl. Pest Manag. Sci. 2022, 78, 287–295. [Google Scholar] [CrossRef]
- Velásquez, J.C.; Bundt, A.D.C.; Camargo, E.R.; Andres, A.; Viana, V.E.; Hoyos, V.; Plaza, G.; de Avila, L.A. Florpyrauxifen-Benzyl Selectivity to Rice. Agriculture 2021, 11, 1270. [Google Scholar] [CrossRef]
- Grossmann, K. Auxin herbicides: Current status of mechanism and mode of action. Pest Manag. Sci. Former. Pestic. Sci. 2009, 66, 113–120. [Google Scholar] [CrossRef]
- Gao, Y.; Li, J.; Pan, X.; Liu, D.; Napier, R.; Dong, L. Quinclorac resistance induced by the suppression of the expression of 1-aminocyclopropane-1-carboxylic acid (ACC) synthase and ACC oxidase genes in Echinochloa crus-galli var. zelayensis. Pestic. Biochem. Physiol. 2018, 146, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuor, H.; Milan, M.; Eckert, J.W.; Fischer, A.J. Quinclorac resistance: A concerted hormonal and enzymatic effort in Echinochloa phyllopogon. Pest Manag. Sci. 2011, 68, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, K.; Scheltrup, F. Selective Induction of 1-Aminocyclopropane-1-carboxylic Acid (ACC) Synthase Activity Is Involved in the Selectivity of the Auxin Herbicide Quinclorac between Barnyard Grass and Rice. Pestic. Biochem. Physiol. 1997, 58, 145–153. [Google Scholar] [CrossRef]
- Grossmann, K.; Kwiatkowski, J. Evidence for a Causative Role of Cyanide, Derived from Ethylene Biosynthesis, in the Herbicidal Mode of Action of Quinclorac in Barnyard Grass. Pestic. Biochem. Physiol. 1995, 51, 150–160. [Google Scholar] [CrossRef]
- Abdallah, I.; Fischer, A.J.; Elmore, C.L.; Saltveit, M.E.; Zaki, M. Mechanism of resistance to quinclorac in smooth crabgrass (Digitaria ischaemum). Pestic. Biochem. Physiol. 2006, 84, 38–48. [Google Scholar] [CrossRef]
- Gao, Y.; Pan, L.; Sun, Y.; Zhang, T.; Dong, L.; Li, J. Resistance to quinclorac caused by the enhanced ability to detoxify cyanide and its molecular mechanism in Echinochloa crus-galli var. zelayensis. Pestic. Biochem. Physiol. 2017, 143, 231–238. [Google Scholar] [CrossRef]
- Kraft, M.; Kuglitsch, R.; Kwiatkowski, J.; Frank, M.; Grossmann, K. Indole-3-acetic acid and auxin herbicides up-regulate 9-cis-epoxycarotenoid dioxygenase gene expression and abscisic acid accumulation in cleavers (Galium aparine): Interaction with ethylene. J. Exp. Bot. 2007, 58, 1497–1503. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Xu, J.; Qi, Y.; He, S.; Li, J.; Dong, L. Differences in ABA synthesis and oxidase stress between quinclorac-resistant and-sensitive Echinochloa crus-galli var. zelayensis. Int. J. Pest Manag. 2021, 1–13. [Google Scholar] [CrossRef]
- Gao, Y.; Pan, X.; Sun, X.; Li, J.; Dong, L. Is the protection of photosynthesis related to the mechanism of quinclorac resistance in Echinochloa crus-galli var. zelayensis? Gene 2019, 683, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Ritz, C.; Streibig, J.C. Bioassay analysis using R. J. Stat. Softw. 2005, 12, 1–22. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org (accessed on 3 January 2022).
- Li, G.; Wu, S.; Cai, L.; Wang, Q.; Zhao, X.; Wu, C. Identification and mRNA expression profile of glutamate receptor-like gene in quinclorac-resistant and susceptible Echinochloa crus-galli. Gene 2013, 531, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Wang, Z.; Cai, J.; Gao, H.; Zhao, H.; Dong, L. High-throughput sequencing reveals differential regulation of miRNAs in fenoxaprop-P-ethyl-resistant Beckmannia syzigachne. Sci. Rep. 2016, 6, 28725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderón Villalobos, L.I.A.; Lee, S.; De Oliveira, C.; Ivetac, A.; Brandt, W.; Armitage, L.; Sheard, L.B.; Tan, X.; Parry, G.; Mao, H. A combinatorial TIR1/AFB–Aux/IAA co-receptor system for differential sensing of auxin. Nat. Chem. Biol. 2012, 8, 477–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeClere, S.; Wu, C.; Westra, P.; Sammons, R.D. Cross-resistance to dicamba, 2, 4-D, and fluroxypyr in Kochia scoparia is endowed by a mutation in an AUX/IAA gene. Proc. Natl. Acad. Sci. USA 2018, 115, E2911–E2920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossmann, K.; Kwiatkowski, J. The Mechanism of Quinclorac Selectivity in Grasses. Pestic. Biochem. Physiol. 2000, 66, 83–91. [Google Scholar] [CrossRef]
- Xu, J.; Liu, X.; Napier, R.; Dong, L.; Li, J. Mode of Action of a Novel Synthetic Auxin Herbicide Halauxifen-Methyl. Agronomy 2022, 12, 1659. [Google Scholar] [CrossRef]
- Chutichude, P.; Chutichudet, B.; Boontiang, K. Influence of 1-MCP Fumigation on Flowering Weight Loss, Water Uptake, Longevity, Anthocyanin Content and Colour of Patumma (Curcuma alismatifolia) cv. Chiang Mai Pink. Int. J. Agric. Res. 2010, 6, 29–39. [Google Scholar] [CrossRef]
- Brown, K.M. Ethylene and abscission. Physiol. Plantarum. 1997, 100, 567–576. [Google Scholar] [CrossRef]
- Lin, C.C.; Hsu, Y.T.; Kao, C.H. Ammonium ion, ethylene, and NaCl-induced senescence of detached rice leaves. Plant Growth Regul. 2002, 37, 85–92. [Google Scholar] [CrossRef]
- Morgan, P.W.; Drew, M.C. Ethylene and plant responses to stress. Physiol. Plantarum. 1997, 100, 620–630. [Google Scholar] [CrossRef]
- Woltering, E.J.; Van Doorn, W.G. Role of ethylene in senescence of petals—Morphological and taxonomical relationships. J. Exp. Bot. 1988, 39, 1605–1616. [Google Scholar] [CrossRef]
- Yang, S.F.; Hoffman, N.E. Ethylene Biosynthesis and its Regulation in Higher Plants. Annu. Rev. Plant Physiol. 1984, 35, 155–189. [Google Scholar] [CrossRef]
- Ikegaya, H.; Iwase, H.; Hatanaka, K.; Sakurada, K.; Yoshida, K.-I.; Takatori, T. Diagnosis of cyanide intoxication by measurement of cytochrome c oxidase activity. Toxicol. Lett. 2001, 119, 117–123. [Google Scholar] [CrossRef]
- Dat, J.; Vandenabeele, S.; Vranova, E.; Van Montagu, M.; Van Breusegem, F. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life Sci. 2000, 57, 779–795. [Google Scholar] [CrossRef] [PubMed]
- Fipke, M.; Vidal, R. Integrative Theory of the Mode of Action of Quinclorac: Literature Review1. Planta Daninha 2016, 34, 393–402. [Google Scholar] [CrossRef]
- Iwakami, S.; Uchino, A.; Kataoka, Y.; Shibaike, H.; Watanabe, H.; Inamura, T. Cytochrome P450 genes induced by bispyribac-sodium treatment in a multiple-herbicide-resistant biotype of Echinochloa phyllopogon. Pest Manag. Sci. 2013, 70, 549–558. [Google Scholar] [CrossRef]
- Yuan, J.S.; Tranel, P.J.; Stewart, C.N., Jr. Non-target-site herbicide resistance: A family business. Trends Plant Sci. 2007, 12, 6–13. [Google Scholar] [CrossRef]
- Barry, C.S.; Blume, B.; Bouzayen, M.; Cooper, W.; Hamilton, A.J.; Grierson, D. Differential expression of the 1-aminocyclopropane-1-carboxylate oxidase gene family of tomato. Plant J. 1996, 9, 525–535. [Google Scholar] [CrossRef]
- Bleecker, A.B.; Kende, H. Ethylene: A Gaseous Signal Molecule in Plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botella, J.R.; Schlagnhaufer, C.D.; Arteca, J.M.; Arteca, R.N.; Phillips, A.T. Identification of two new members of the l-aminocyclopropane-l-carboxylate synthase-encoding multigene family in mung bean. Gene 1993, 123, 249–253. [Google Scholar] [CrossRef]
- Kende, H.; Zeevaart, J. The Five "Classical" Plant Hormones. Plant Cell 1997, 9, 1197–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Abel, S.; A Keller, J.; Shen, N.F.; Theologis, A. The 1-aminocyclopropane-1-carboxylate synthase gene family of Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1992, 89, 11046–11050. [Google Scholar] [CrossRef] [Green Version]
- Yip, W.K.; Moore, T.; Yang, S.F. Differential accumulation of transcripts for four tomato 1-aminocyclopropane-1-carboxylate synthase homologs under various conditions. Proc. Natl. Acad. Sci. USA 1992, 89, 2475–2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rottmann, W.H.; Peter, G.F.; Oeller, P.W.; Keller, J.A.; Shen, N.F.; Nagy, B.P.; Taylor, L.P.; Campbell, A.D.; Theologis, A. 1-Aminocyclopropane-1-carboxylate synthase in tomato is encoded by a multigene family whose transcription is induced during fruit and floral senescence. J. Mol. Biol. 1991, 222, 937–961. [Google Scholar] [CrossRef]
- Yamagami, T.; Tsuchisaka, A.; Yamada, K.; Haddon, W.F.; Harden, L.A.; Theologis, A. Biochemical Diversity among the 1-Amino-cyclopropane-1-Carboxylate Synthase Isozymes Encoded by the Arabidopsis Gene Family. J. Biol. Chem. 2003, 278, 49102–49112. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.H.; Qin, X.; Zeevaart, J.A. Elucidation of the Indirect Pathway of Abscisic Acid Biosynthesis by Mutants, Genes, and Enzymes. Plant Physiol. 2003, 131, 1591–1601. [Google Scholar] [CrossRef]
- Tan, B.-C.; Joseph, L.M.; Deng, W.-T.; Liu, L.; Li, Q.-B.; Cline, K.; McCarty, D.R. Molecular characterization of the Arabidopsis 9-cis epoxycarotenoid dioxygenase gene family. Plant J. 2003, 35, 44–56. [Google Scholar] [CrossRef]
Figure 1.
Dose–response analyses for response of barnyardgrass and yerbadetajo to florpyrauxifen-benzyl. The X-axis represents the dose (g a.i. ha−1). The Y-axis represents percentages of fresh weight (% untreated control).
Figure 1.
Dose–response analyses for response of barnyardgrass and yerbadetajo to florpyrauxifen-benzyl. The X-axis represents the dose (g a.i. ha−1). The Y-axis represents percentages of fresh weight (% untreated control).

Figure 2.
ACC content in barnyardgrass and yerbadetajo after florpyrauxifen-benzyl treatment. The X-axis represents the treatment time; the Y-axis represents ACC content. a–d represents the significance level (p < 0.05) (the significance mark of barnyardgrass is above the broken line, and the significance mark of yerbadetajo is below the broken line). The standard errors of the means are described by vertical bars.
Figure 2.
ACC content in barnyardgrass and yerbadetajo after florpyrauxifen-benzyl treatment. The X-axis represents the treatment time; the Y-axis represents ACC content. a–d represents the significance level (p < 0.05) (the significance mark of barnyardgrass is above the broken line, and the significance mark of yerbadetajo is below the broken line). The standard errors of the means are described by vertical bars.

Figure 3.
ABA content in barnyardgrass and yerbadetajo after florpyrauxifen-benzyl treatment. The X-axis represents the treatment time; the Y-axis represents ABA content. a–f represents the significance level (p < 0.05) (the significance mark of barnyardgrass is above the broken line, and the significance mark of yerbadetajo is below the broken line). Vertical bars represent standard errors of the means.
Figure 3.
ABA content in barnyardgrass and yerbadetajo after florpyrauxifen-benzyl treatment. The X-axis represents the treatment time; the Y-axis represents ABA content. a–f represents the significance level (p < 0.05) (the significance mark of barnyardgrass is above the broken line, and the significance mark of yerbadetajo is below the broken line). Vertical bars represent standard errors of the means.

Figure 4.
KEGG Ortholog database (KO) analysis. The vertical axis represents the entries of the pathways, and the horizontal axis represents the number of genes. Columns of the same color represent the same first class. B3-B24 means barnyardgrass was treated with florpyrauxifen-benzoyl for 3, 6, 12, or 24 h. Y3-Y24 means yerbadetajo was treated with florpyrauxifen-benzoyl for 3, 6, 12, or 24 h. (a). KO analysis for barnyardgrass after florpyrauxifen-benzoyl treatment. (b). KO analysis for yerbadetajo after florpyrauxifen-benzoyl treatment.
Figure 4.
KEGG Ortholog database (KO) analysis. The vertical axis represents the entries of the pathways, and the horizontal axis represents the number of genes. Columns of the same color represent the same first class. B3-B24 means barnyardgrass was treated with florpyrauxifen-benzoyl for 3, 6, 12, or 24 h. Y3-Y24 means yerbadetajo was treated with florpyrauxifen-benzoyl for 3, 6, 12, or 24 h. (a). KO analysis for barnyardgrass after florpyrauxifen-benzoyl treatment. (b). KO analysis for yerbadetajo after florpyrauxifen-benzoyl treatment.

Figure 5.
ACS and ACO gene expression patterns after florpyrauxifen-benzoyl treatment. The X-axis represents the time after florpyrauxifen-benzoyl treatment. The Y-axis represents transcript levels relative to 0 h untreated with florpyrauxifen-benzoyl. Data are the mean values of six technical and biological replicates. The standard errors of the means are described by Vertical bars represent standard errors of the means. The icon “▲” represents genes that were up-regulated more than 2 fold by florpyrauxifen-benzoyl and significant high than the untreated control group (0 h). (a) ACS and ACO gene expression patterns in barnyardgrass. (b) ACS and ACO gene expression patterns in yerbadetajo.
Figure 5.
ACS and ACO gene expression patterns after florpyrauxifen-benzoyl treatment. The X-axis represents the time after florpyrauxifen-benzoyl treatment. The Y-axis represents transcript levels relative to 0 h untreated with florpyrauxifen-benzoyl. Data are the mean values of six technical and biological replicates. The standard errors of the means are described by Vertical bars represent standard errors of the means. The icon “▲” represents genes that were up-regulated more than 2 fold by florpyrauxifen-benzoyl and significant high than the untreated control group (0 h). (a) ACS and ACO gene expression patterns in barnyardgrass. (b) ACS and ACO gene expression patterns in yerbadetajo.

Figure 6.
NCED gene expression patterns in barnyardgrass (EcNCED) and yerbadetajo (EpNCED) after florpyrauxifen-benzoyl treatment. The X-axis represents the time after florpyrauxifen-benzoyl treatment. The Y-axis represents transcript levels relative to 0 h untreated with florpyrauxifen-benzoyl. Data are the mean values of six technical and biological replicates. The standard errors of the means are described by vertical bars. The icon “▲” represents genes that were up-regulated more than 2 fold by florpyrauxifen-benzoyl and significant high than the untreated control group (0 h).
Figure 6.
NCED gene expression patterns in barnyardgrass (EcNCED) and yerbadetajo (EpNCED) after florpyrauxifen-benzoyl treatment. The X-axis represents the time after florpyrauxifen-benzoyl treatment. The Y-axis represents transcript levels relative to 0 h untreated with florpyrauxifen-benzoyl. Data are the mean values of six technical and biological replicates. The standard errors of the means are described by vertical bars. The icon “▲” represents genes that were up-regulated more than 2 fold by florpyrauxifen-benzoyl and significant high than the untreated control group (0 h).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Selected reaction monitoring conditions for protonated or deprotonated plant hormones ([M+H] + or [M−H]−).
Table 1.
Selected reaction monitoring conditions for protonated or deprotonated plant hormones ([M+H] + or [M−H]−).
| Substance | Polarity | Parent Ion (m/z) | Daughter Ion (m/z) | De Clustering Voltage (V) | Collision Energy (V) |
|---|---|---|---|---|---|
| ACC | + | 102.0 | 55.9 */84.0 | 40 | 19/42 |
* Quantitative ion.
Table 2.
Gradient Parameters of HPLC.
| Time (min) | Flow Velocity (mL/min) | A% |
|---|---|---|
| 0–1 | 0.3 | 20 |
| 1–3 | 0.3 | Increase from 20 to 50 |
| 3–9 | 0.3 | Increase from 50 to 80 |
| 9–10.5 | 0.3 | 80 |
| 10.5–10.6 | 0.3 | Decrease from 80 to 20 |
| 10.6–13.5 | 0.3 | 20 |
Table 3.
Selected reaction monitoring conditions for protonated or deprotonated plant hormones ([M+H] + or [M−H]−).
Table 3.
Selected reaction monitoring conditions for protonated or deprotonated plant hormones ([M+H] + or [M−H]−).
| Substance | Polarity | Parent Ion (m/z) | Daughter Ion (m/z) | De Clustering Voltage (V) | Collision Energy (V) |
|---|---|---|---|---|---|
| ABA | - | 263.1 | 153.1 */204.2 | −60 | −14/−27 |
* Quantitative ion.
Table 4.
Differentially expressed genes compared to untreated control after florpyrauxifen-benzyl treatment in barnyardgrass and yerbadetajo.
Table 4.
Differentially expressed genes compared to untreated control after florpyrauxifen-benzyl treatment in barnyardgrass and yerbadetajo.
| Weed Species | Gene Expression Trend | Time after Florpyrauxifen-Benzyl Treatment | |||
|---|---|---|---|---|---|
| 3 h | 6 h | 12 h | 24 h | ||
| Barnyardgrass | Up-regulated genes | 9162 | 11,545 | 12,150 | 9440 |
| Down-regulated genes | 4148 | 4047 | 4189 | 3289 | |
| All Differentially expressed genes | 13,310 | 15,592 | 16,339 | 12,729 | |
| Yerbadetajo | Up-regulated genes | 2172 | 3589 | 3860 | 4567 |
| Down-regulated genes | 3925 | 3427 | 5718 | 6637 | |
| All Differentially expressed genes | 6097 | 7016 | 9578 | 11,204 | |
Table 5.
Candidate DEGs related to ethylene and ABA biosynthesis identified from transcriptome data of barnyardgrass and yerbadetajo.
Table 5.
Candidate DEGs related to ethylene and ABA biosynthesis identified from transcriptome data of barnyardgrass and yerbadetajo.
| Genes | Best BLASTX Match | |
|---|---|---|
| Name and Accession Number | Ident (%) | |
| EcACS | ACS (Echinochloa crus-galli) (KT245168.1) | 96.95 |
| EcACS3 | ACS 3 (E. crus-galli var. zelayensis) (KY963551) | 100.00 |
| EcACO1-like | ACO 1-like (E. crus-galli var. zelayensis) (KY963553.1) | 96.64 |
| EcACO homolog 1 | EcACO homolog 1 (Setaria italica) (XM_004975083.3) | 89.89 |
| EcACO 4-like | ACO 4-like (E. crus-galli var. zelayensis) (KY963555) | 100.00 |
| EcACO homolog 4 | ACO 4-like (E. crus-galli var. zelayensis) (KY963556) | 100.00 |
| EcACO5-like | ACO 5-like (E. crus-galli var. zelayensis) (KY963548) | 100.00 |
| EcACO homolog 6-like | ACO 6-like (E. crus-galli var. zelayensis) (KY963557) | 100.00 |
| EcNCED | NCED (E. crus-galli var. zelayensis) [45] | 100.00 |
| EpACS-1 | ACS (Helianthus annuus) (XM_022113477.2) | 85.06 |
| EpACS-2 | ACS (Helianthus annuus) (XM_022144560.2) | 88.86 |
| EpACS3-1 | ACS 3 (Helianthus annuus) (XM_022154426.2) | 88.42 |
| EpACS3-2 | ACS 3 (Helianthus annuus) (XM_022142807.2) | 88.39 |
| EpACS3-3 | ACS 3 (Helianthus annuus) (XM_022179292.2) | 86.04 |
| EpACS7 | ACS 7 (Helianthus annuus) (XM_022131322.1) | 91.77 |
| EpACS-like | ACS-like (Helianthus annuus) (XM_035986598.1) | 86.21 |
| EpACO-homolog 1-1 | ACO homolog 1 (Helianthus annuus) (XM_035987786.1) | 85.21 |
| EpACO-homolog 1-2 | ACO homolog 1 (Helianthus annuus) (XM_022146678.2) | 81.67 |
| EpACO-homolog 1-like | ACO homolog 1-like (Helianthus annuus) (XM_022174330.2) | 85.69 |
| EpACO homolog 3 | ACO homolog 3 (Helianthus annuus) (XM_022170011.2) | 88.71 |
| EpACO3 | ACO 3 (Helianthus annuus) (XM_022155132.2) | 88.06 |
| EpACO | ACO (Helianthus annuus) (XM_022155410.2) | 84.07 |
| EpNCED1-1 | NCED1 (Helianthus annuus) (XM_022157417.2) | 85.73 |
| EpNCED1-2 | NCED1 (Helianthus annuus) (XM_022133305.2) | 85.99 |
| EpNCED2 | NCED2 (Helianthus annuus) (XM_022179446.2) | 83.52 |
The prefix “Ec”, the abbreviation of Echinochloa crus-galli represents the genes in barnyardgrass, and the prefix “Ep”, the abbreviation of Eclipta prostrata represents the genes in yerbadetajo.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gao, Y.; Li, J.; Shen, G.; Tian, Z. Differences in the Mode of Action of Florpyrauxifen-Benzyl between Barnyardgrass and Yerbadetajo. Agronomy 2022, 12, 2656. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12112656
AMA Style
Gao Y, Li J, Shen G, Tian Z. Differences in the Mode of Action of Florpyrauxifen-Benzyl between Barnyardgrass and Yerbadetajo. Agronomy. 2022; 12(11):2656. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12112656
Chicago/Turabian StyleGao, Yuan, Jun Li, Guohui Shen, and Zhihui Tian. 2022. "Differences in the Mode of Action of Florpyrauxifen-Benzyl between Barnyardgrass and Yerbadetajo" Agronomy 12, no. 11: 2656. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12112656
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.