

Genome-Wide Identification, Structural Characterization, and Gene Expression Analysis of BES1 Transcription Factor Family in Tartary Buckwheat (Fagopyrum tataricum)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome-Wide Identification and Sequence Analysis of FtBES1 Family Members
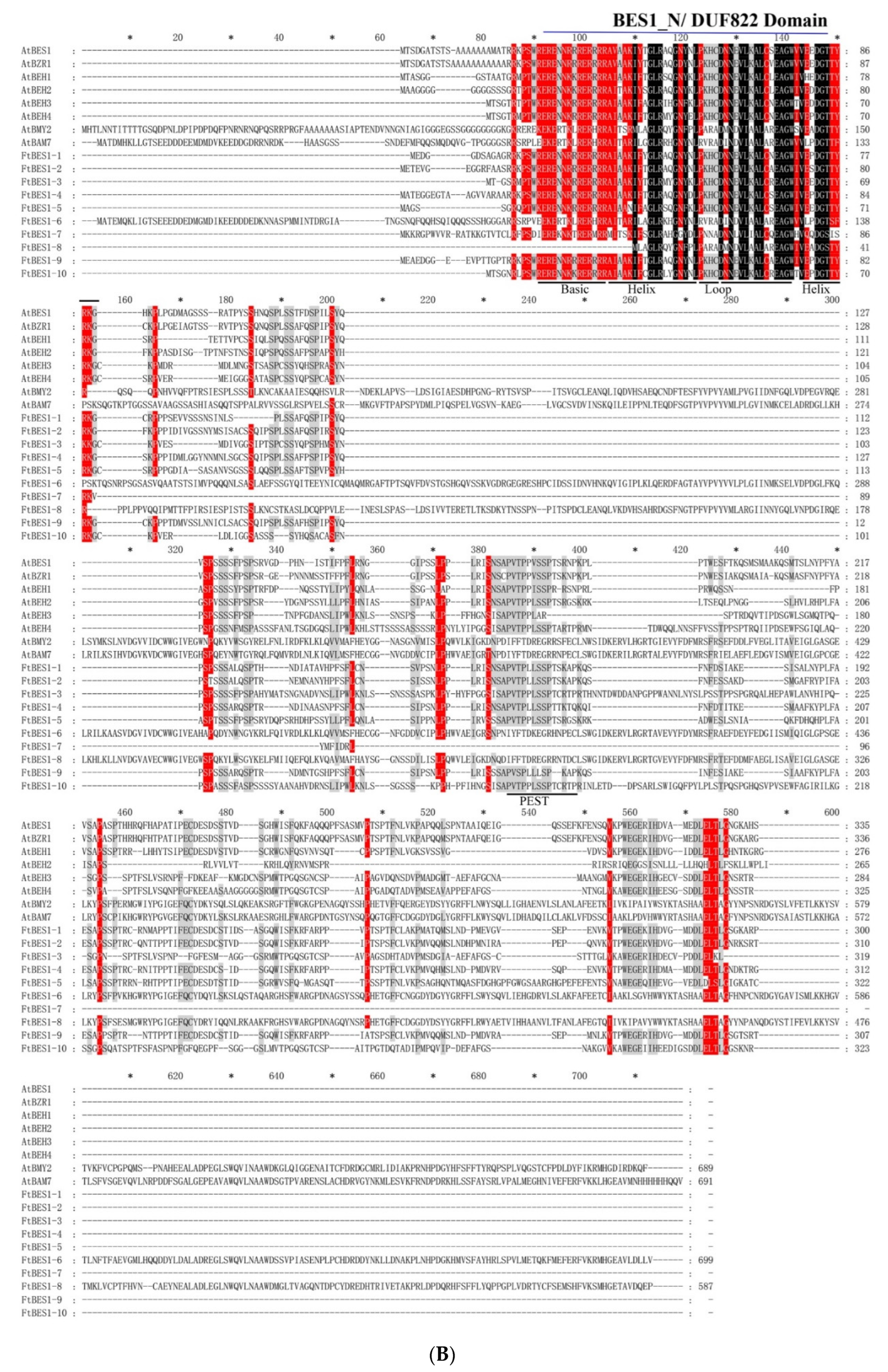
2.2. Phylogenetic Analysis and Multiple Sequence Alignments of FtBES1 Family Members
2.3. Gene Localization and Chromosome Duplication Analysis of FtBES1 Family Members
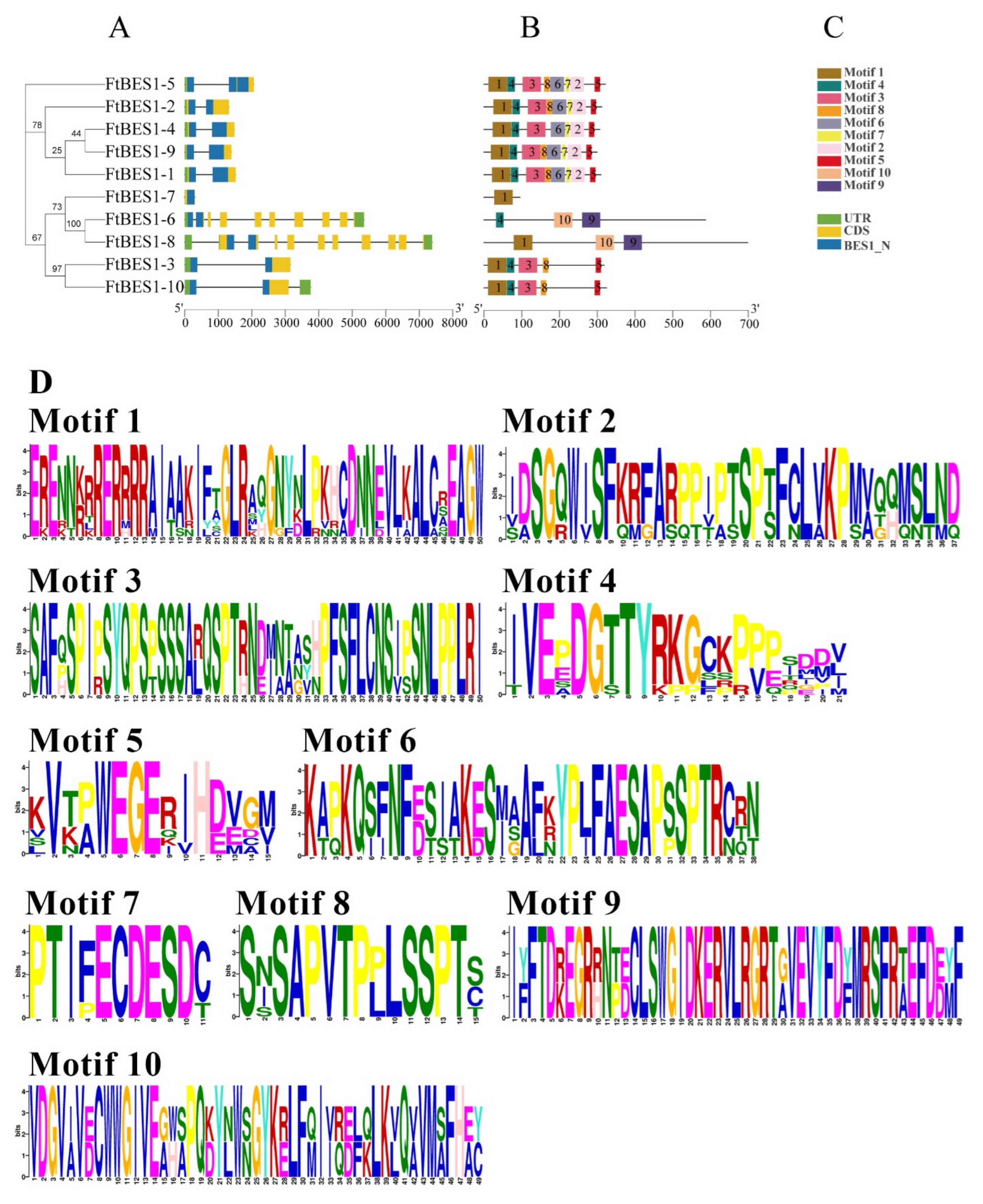
2.4. Analysis of Conserved Motifs and Cis-Acting Elements of FtBES1 Family Members
2.5. Plant Materials and Stress Treatment
2.6. RNA Extraction and Gene Expression Analysis
3. Results
3.1. Identification and Analysis of FtBES1
3.2. Phylogenetic and Multiple Sequence Analysis of FtBES1
3.3. Analysis of FtBES1 Gene Structure and Motif
3.4. Analysis of Cis-Acting Elements in the FtBES1 Promoter
3.5. Chromosome Mapping, Gene Duplication, and Evolutionary Analysis
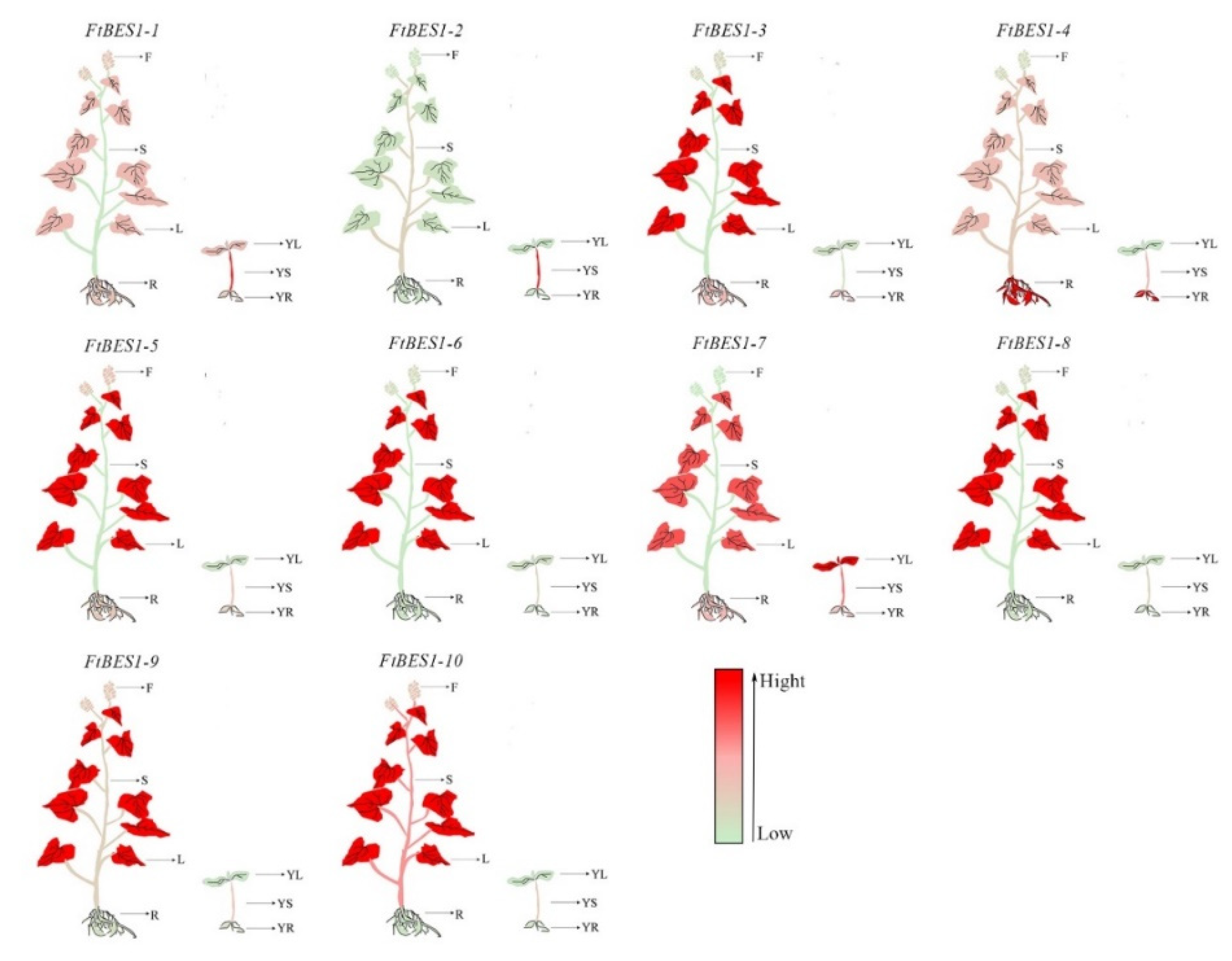
3.6. Expression Pattern of FtBES1 Gene in Different Tissues
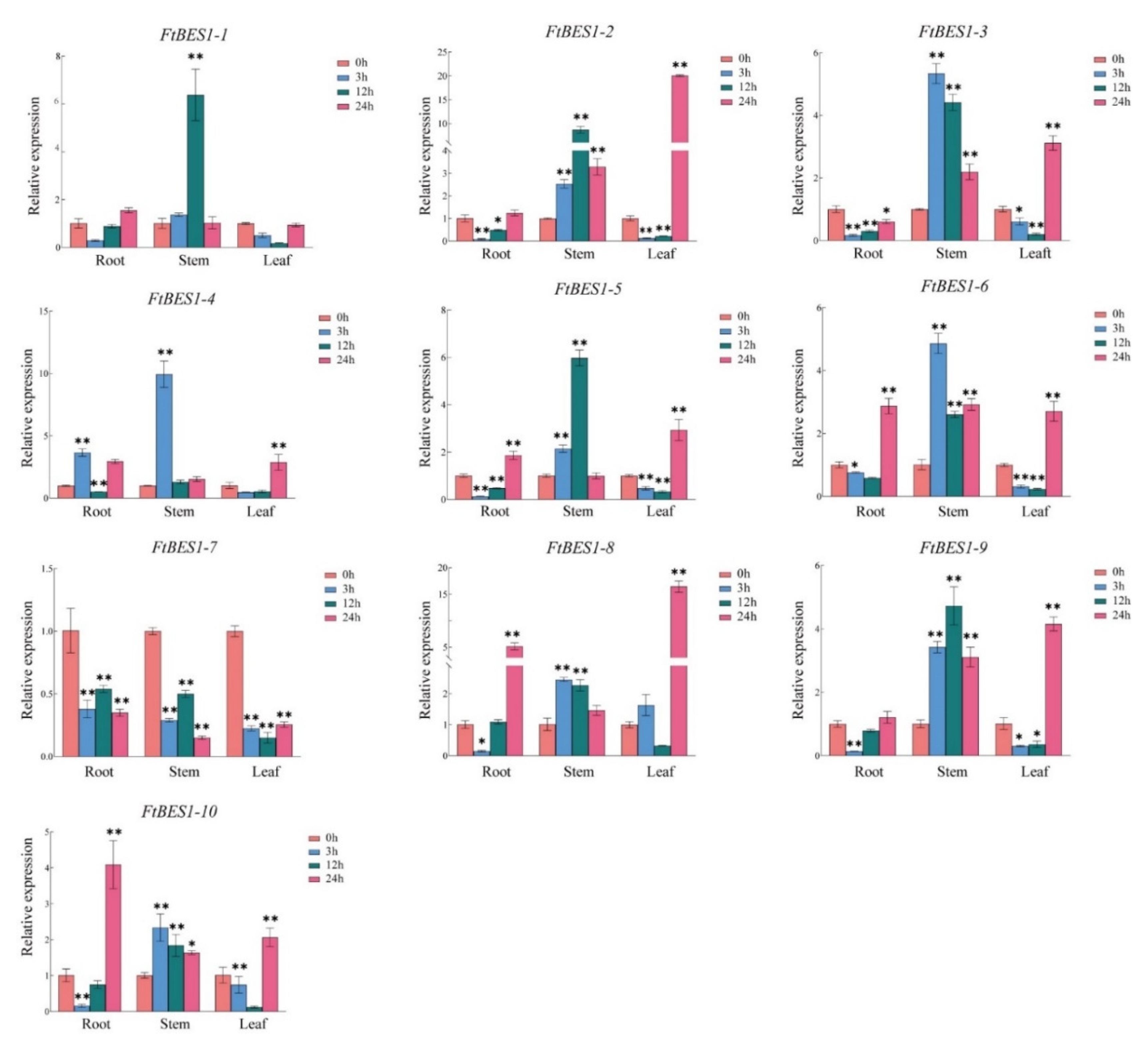
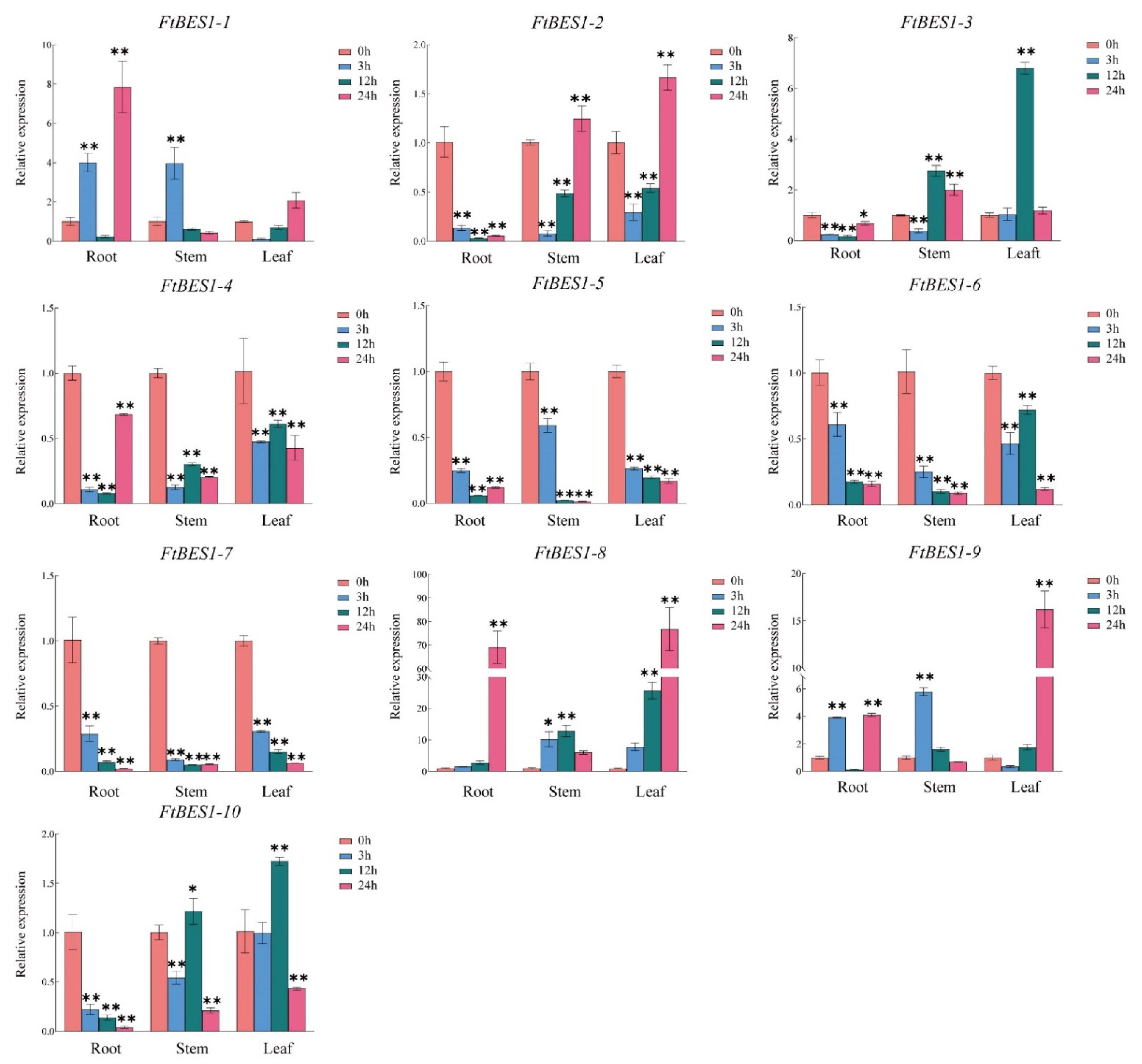
3.7. Expression Analysis of FtBES1 Gene in Response to Hormone Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gonzalez-Garcia, M.P.; Vilarrasa-Blasi, J.; Zhiponova, M.; Divol, F.; Mora-Garcia, S.; Russinova, E.; Cano-Delgado, A.I. Brassinosteroids control meristem size by promoting cell cycle progression in Arabidopsis roots. Development 2011, 138, 849–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.Y.; Bai, M.Y.; Oh, E.; Zhu, J.Y. Brassinosteroid signaling network and regulation of photomorphogenesis. Annu. Rev. Genet. 2012, 46, 701–724. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.F.; Li, L.; Zola, J.; Aluru, M.; Ye, H.X.; Foudree, A.; Guo, H.Q.; Anderson, S.; Aluru, S.; Liu, P.; et al. A brassinosteroid transcriptional network revealed by genome-wide identification of BESI target genes in Arabidopsis thaliana. Plant J. 2011, 65, 634–646. [Google Scholar] [CrossRef]
- Yin, Y.; Vafeados, D.; Tao, Y.; Yoshida, S.; Asami, T.; Chory, J. A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell 2005, 120, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Clouse, S.D. Brassinosteroid signal transduction: From receptor kinase activation to transcriptional networks regulating plant development. Plant Cell 2011, 23, 1219–1230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.J.; Zhang, C.; Lu, Y.N.; Jin, J.Q.; Wang, X.L. The mechanisms of brassinosteroids’ action: From signal transduction to plant development. Mol. Plant 2011, 4, 588–600. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Yu, X.F.; Thompson, A.; Guo, M.; Yoshida, S.; Asami, T.; Chory, J.; Yin, Y. Arabidopsis MYB30 is a direct target of BES1 and cooperates with BES1 to regulate brassinosteroid-induced gene expression. Plant J. 2009, 58, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.F.; Qian, H.M.; Shen, W.S.; Liu, L.H.; Zhang, M.; Cai, C.X.; Zhao, Y.T.; Qiao, J.J.; Wang, Q.M. BZR1 and BES1 participate in regulation of glucosinolate biosynthesis by brassinosteroids in Arabidopsis. J. Exp. Bot. 2013, 64, 2401–2412. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.N.; Nolan, T.M.; Ye, H.X.; Zhang, M.C.; Tong, H.N.; Xin, P.Y.; Chu, J.F.; Chu, C.C.; Li, Z.H.; Yin, Y.H. Arabidopsis WRKY46, WRKY54, and WRKY70 Transcription Factors Are Involved in Brassinosteroid-Regulated Plant Growth and Drought Responses. Plant Cell 2017, 29, 1425–1439. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Xu, P.; Wang, W.; Wang, S.; Caruana, J.C.; Yang, H.Q.; Lian, H. Arabidopsis G-Protein beta Subunit AGB1 Interacts with BES1 to Regulate Brassinosteroid Signaling and Cell Elongation. Front. Plant Sci. 2017, 8, 2225. [Google Scholar] [CrossRef]
- Ye, H.X.; Liu, S.Z.; Tang, B.Y.; Chen, J.N.; Xie, Z.L.; Nolan, T.M.; Jiang, H.; Guo, H.Q.; Lin, H.Y.; Li, L.; et al. RD26 mediates crosstalk between drought and brassinosteroid signalling pathways. Nat. Commun. 2017, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Ryu, H.; Cho, H.; Bae, W.; Hwang, I. Control of early seedling development by BES1/TPL/HDA19-mediated epigenetic regulation of ABI3. Nat. Commun. 2014, 5, 4138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Dou, L.R.; Gong, Z.Z.; Wang, X.F.; Mao, T.L. BES1 hinders ABSCISIC ACID INSENSITIVE5 and promotes seed germination in Arabidopsis. New Phytol. 2019, 221, 908–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Wang, W.X.; Xu, P.B.; Pan, J.; Zhang, T.; Li, Y.; Li, G.W.; Yang, H.Q.; Lian, H.L. phyB Interacts with BES1 to Regulate Brassinosteroid Signaling in Arabidopsis. Plant Cell Physiol. 2019, 60, 353–366. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, S.Y.; Zhu, W.J.; Jia, K.P.; Yang, H.Q.; Wang, X.L. Strigolactone/MAX2-Induced Degradation of Brassinosteroid Transcriptional Effector BES1 Regulates Shoot Branching. Dev. Cell 2013, 27, 681–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albertos, P.; Dündar, G.; Schenk, P.; Carrera, S.; Cavelius, P.; Sieberer, T.; Poppenberger, B. Transcription factor BES1 interacts with HSFA1 to promote heat stress resistance of plants. EMBO J. 2022, 41, e108664. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Jiang, J.J.; Zhang, C.; Wang, X.L. A Recently Evolved Isoform of the Transcription Factor BES1 Promotes Brassinosteroid Signaling and Development in Arabidopsis thaliana. Plant Cell 2015, 27, 361–374. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Wang, Z.Y.; Mora-Garcia, S.; Li, J.; Yoshida, S.; Asami, T.; Chory, J. BES1 accumulates in the nucleus in response to brassinosteroids to regulate gene expression and promote stem elongation. Cell 2002, 109, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.W.; Wu, X.Y.; Qiu, S.Y.; Zheng, H.Y.; Lu, Y.W.; Peng, J.J.; Wu, G.W.; Chen, J.P.; Rao, S.F.; Yan, F. Genome-Wide Identification and Expression Profiling of the BZR Transcription Factor Gene Family in Nicotiana benthamiana. Int. J. Mol. Sci. 2021, 22, 10379. [Google Scholar] [CrossRef]
- Sarwar, R.; Geng, R.; Li, L.; Shan, Y.; Zhu, K.M.; Wang, J.; Tan, X.L. Genome-Wide Prediction, Functional Divergence, and Characterization of Stress-Responsive BZR Transcription Factors in B. napus. Front. Plant Sci. 2021, 12, 790655. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Cui, Y.J.; Zhao, Z.L.; Li, S.Y.; Liang, D.; Wang, C.L.; Feng, G.; Wang, J.H.; Liu, Z.L. Genome-wide identification and characterization of the BES/BZR gene family in wheat and foxtail millet. BMC Genom. 2021, 22, 14. [Google Scholar] [CrossRef] [PubMed]
- Su, D.D.; Xiang, W.; Wen, L.; Lu, W.; Shi, Y.; Liu, Y.D.; Li, Z.G. Genome-wide identification, characterization and expression analysis of BES1 gene family in tomato. BMC Plant Biol. 2021, 21, 17. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Khaliq, A.; Lu, S.; Xie, M.; Ma, Z.; Mao, J.; Chen, B. Genome-wide identification and characterization of the BES1 gene family in apple (Malus domestica). Plant Biol. 2020, 22, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Qanmber, G.; Lu, L.L.; Qin, W.Q.; Liu, J.; Li, J.; Ma, S.Y.; Yang, Z.E.; Yang, Z.R. Genome-wide analysis of BES1 genes in Gossypium revealed their evolutionary conserved roles in brassinosteroid signaling. Sci. China Life Sci. 2018, 61, 1566–1582. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Feng, W.; Sun, F.; Zhang, Y.; Qu, J.; Liu, B.; Lu, F.; Yang, L.; Fu, F.; Li, W. Cloning and characterization of BES1/BZR1 transcription factor genes in maize. Plant Growth Regul. 2018, 86, 235–249. [Google Scholar] [CrossRef]
- Chen, L.-G.; Gao, Z.; Zhao, Z.; Liu, X.; Li, Y.; Zhang, Y.; Liu, X.; Sun, Y.; Tang, W. BZR1 Family Transcription Factors Function Redundantly and Indispensably in BR Signaling but Exhibit BRI1-Independent Function in Regulating Anther Development in Arabidopsis. Mol. Plant 2019, 12, 1408–1415. [Google Scholar] [CrossRef]
- Cui, X.Y.; Gao, Y.; Guo, J.; Yu, T.F.; Zheng, W.J.; Liu, Y.W.; Chen, J.; Xu, Z.S.; Ma, Y.Z. BES/BZR Transcription Factor TaBZR2 Positively Regulates Drought Responses by Activation of TaGST1. Plant Physiol. 2019, 180, 605–620. [Google Scholar] [CrossRef] [Green Version]
- Jia, C.G.; Zhao, S.K.; Bao, T.T.; Zhao, P.Q.; Peng, K.; Guo, Q.X.; Gao, X.; Qin, J.C. Tomato BZR/BES transcription factor SlBZR1 positively regulates BR signaling and salt stress tolerance in tomato and Arabidopsis. Plant Sci. 2021, 302, 110719. [Google Scholar] [CrossRef]
- Shi, Z.M.; Chen, X.; Xue, H.D.; Jia, T.T.; Meng, F.N.; Liu, Y.F.; Luo, X.M.; Xiao, G.H.; Zhu, S.W. GhBZR3 suppresses cotton fiber elongation by inhibiting very-long-chain fatty acid biosynthesis. Plant J. 2022, 111, 785–799. [Google Scholar] [CrossRef]
- Oh, E.; Zhu, J.Y.; Wang, Z.Y. Interaction between BZR1 and PIF4 integrates brassinosteroid and environmental responses. Nat. Cell Biol. 2012, 14, 802–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.; Jeong, Y.J.; Corvalan, C.; Fujioka, S.; Cho, S.; Park, T.; Choe, S. Darkness and gulliver2/phyB mutation decrease the abundance of phosphorylated BZR1 to activate brassinosteroid signaling in Arabidopsis. Plant J. 2014, 77, 737–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.F.; He, J.X. BZR1 Interacts with HY5 to Mediate Brassinosteroid- and Light-Regulated Cotyledon Opening in Arabidopsis in Darkness. Mol. Plant 2016, 9, 113–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, T.; Shi, C.; Peng, Y.; Tan, H.; Xin, P.; Yang, Y.; Wang, F.; Li, X.; Chu, J.; Huang, J.; et al. Brassinosteroid-Activated BRI1-EMS-SUPPRESSOR 1 Inhibits Flavonoid Biosynthesis and Coordinates Growth and UV-B Stress Responses in Plants. Plant Cell 2020, 32, 3224–3239. [Google Scholar] [CrossRef]
- Guo, H.Q.; Li, L.; Aluru, M.; Aluru, S.; Yin, Y.H. Mechanisms and networks for brassinosteroid regulated gene expression. Curr. Opin. Plant Biol. 2013, 16, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Song, J.H.; Park, S.U.; Jeong, Y.S.; Kim, S.H. Brassinosteroid-Induced Transcriptional Repression and Dephosphorylation-Dependent Protein Degradation Negatively Regulate BIN2-Interacting AIF2 (a BR Signaling-Negative Regulator) bHLH Transcription Factor. Plant Cell Physiol. 2017, 58, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Yang, H.; Dai, L.; Zhao, X.; Wang, L.F. Genome-wide identification and response stress expression analysis of the BES1 family in rubber tree (Hevea brasiliensis Muell. Arg.). PeerJ 2022, 10, e13189. [Google Scholar] [CrossRef]
- Lilley, J.L.S.; Gan, Y.B.; Graham, I.A.; Nemhauser, J.L. The effects of DELLAs on growth change with developmental stage and brassinosteroid levels. Plant J. 2013, 76, 165–173. [Google Scholar]
- Unterholzner, S.J.; Rozhon, W.; Papacek, M.; Ciomas, J.; Lange, T.; Kugler, K.G.; Mayer, K.F.; Sieberer, T.; Poppenberger, B. Brassinosteroids Are Master Regulators of Gibberellin Biosynthesis in Arabidopsis. Plant Cell 2015, 27, 2261–2272. [Google Scholar] [CrossRef] [Green Version]
- Tong, H.N.; Xiao, Y.H.; Liu, D.P.; Gao, S.P.; Liu, L.C.; Yin, Y.H.; Jin, Y.; Qian, Q.; Chu, C.C. Brassinosteroid Regulates Cell Elongation by Modulating Gibberellin Metabolism in Rice. Plant Cell 2014, 26, 4376–4393. [Google Scholar] [CrossRef] [Green Version]
- Que, F.; Khadr, A.; Wang, G.L.; Li, T.; Wang, Y.H.; Xu, Z.S.; Xiong, A.S. Exogenous brassinosteroids altered cell length, gibberellin content, and cellulose deposition in promoting carrot petiole elongation. Plant Sci. 2018, 277, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Su, D.D.; Xiang, W.; Liang, Q.; Wen, L.; Shi, Y.; Song, B.Q.; Liu, Y.D.; Xian, Z.Q.; Li, Z.G. Tomato SlBES1.8 Influences Leaf Morphogenesis by Mediating Gibberellin Metabolism and Signaling. Plant Cell Physiol. 2022, 63, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.Q.; He, Y.Q.; Zhang, K.X.; Li, J.B.; Shi, Y.L.; Zhao, M.Y.; Meng, Y.; Georgiev, M.I.; Zhou, M.L. JA-induced FtBPM3 accumulation promotes FtERF-EAR3 degradation and rutin biosynthesis in Tartary buckwheat. Plant J. 2022, 111, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Hou, S.; Yang, W.; Han, Y. Exogenous application of salicylic acid enhanced the rutin accumulation and influenced the expression patterns of rutin biosynthesis related genes in Fagopyrum tartaricum Gaertn leaves. Plant Growth Regul. 2012, 68, 9–15. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Full Length (bp) | CDS (bp) | Amino Acid (aa) | Molecular | pI | GRAVY | Predicted Subcellular Localization | Alpha Helix | Beta Turn | Random Coil |
|---|---|---|---|---|---|---|---|---|---|---|
| FtBES1-1 | 1391 | 903 | 300 | 32,678.69 | 8.1 | −0.573 | Nucleus | 14.33% | 3.33% | 70.67% |
| FtBES1-2 | 1521 | 933 | 310 | 34,426.71 | 9.25 | −0.66 | Nucleus | 13.23% | 4.52% | 73.87% |
| FtBES1-3 | 3153 | 960 | 319 | 34,435.37 | 7.1 | −0.645 | Nucleus | 17.87% | 5.02% | 69.91% |
| FtBES1-4 | 1303 | 939 | 312 | 34,388.88 | 9.03 | −0.604 | Nucleus | 19.87% | 5.77% | 67.31% |
| FtBES1-5 | 2065 | 969 | 322 | 34,830.4 | 8.29 | −0.725 | Nucleus | 14.91% | 6.52% | 66.77% |
| FtBES1-6 | 7386 | 2100 | 699 | 78,544.06 | 5.69 | −0.476 | Nucleus | 34.76% | 6.58% | 45.64% |
| FtBES1-7 | 315 | 291 | 96 | 11,163 | 10.52 | −0.644 | Nucleus | 36.46% | 13.54% | 34.38% |
| FtBES1-8 | 5353 | 1764 | 587 | 66,318.77 | 5.24 | −0.32 | Nucleus | 31.86% | 6.64% | 49.57% |
| FtBES1-9 | 1482 | 924 | 307 | 33,720.08 | 8.36 | −0.585 | Nucleus | 14.98% | 5.21% | 70.68% |
| FtBES1-10 | 3756 | 978 | 325 | 34,807.81 | 8.45 | −5.02 | Nucleus | 16.92% | 4.31% | 68.62% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, A.; Weng, W.; Yao, X.; Wu, W.; Bai, Q.; Xiong, R.; Ma, C.; Cheng, J.; Ruan, J. Genome-Wide Identification, Structural Characterization, and Gene Expression Analysis of BES1 Transcription Factor Family in Tartary Buckwheat (Fagopyrum tataricum). Agronomy 2022, 12, 2729. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12112729
Gao A, Weng W, Yao X, Wu W, Bai Q, Xiong R, Ma C, Cheng J, Ruan J. Genome-Wide Identification, Structural Characterization, and Gene Expression Analysis of BES1 Transcription Factor Family in Tartary Buckwheat (Fagopyrum tataricum). Agronomy. 2022; 12(11):2729. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12112729
Chicago/Turabian StyleGao, Anjing, Wenfeng Weng, Xin Yao, Weijiao Wu, Qing Bai, Ruiqi Xiong, Chao Ma, Jianping Cheng, and Jingjun Ruan. 2022. "Genome-Wide Identification, Structural Characterization, and Gene Expression Analysis of BES1 Transcription Factor Family in Tartary Buckwheat (Fagopyrum tataricum)" Agronomy 12, no. 11: 2729. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12112729