



Adaptive Responses of Four Medicinal Plants to High Altitude Oxidative Stresses through the Regulation of Antioxidants and Secondary Metabolites
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Target Species

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Family | Plant Species | Habitat, Status, Uses, and Distribution | Field Photo |
|---|---|---|---|---|
| 1 | Rosaceae | Cotoneaster orbicularis Schltdl. |
|  |
| 2 | Crataegus x sinaica Boiss. |
|  | |
| 3 | Asteraceae | Echinops spinosissimus subsp. Spinosissimus Turra. |
|  |
| 4 | Tanacetum sinaicum Del. |
|  |
2.3. Soil Analysis
2.4. Phytochemical Assay
2.4.1. Extraction and Estimation of Photosynthetic Pigments
2.4.2. Extraction and Estimation of Total Soluble Sugars
2.4.3. Extraction and Estimation of Total Phenolic, Flavonoid, and Tannin Content
2.5. Biochemical Assay
2.5.1. Extraction and Estimation of Total Soluble Proteins and Proline Content
2.5.2. Extraction and Estimation of Malondialdehyde
2.6. Enzymes Extraction and Assays
2.7. Determination of Ascorbic Acid (AsA) and Total Antioxidant Capacity (TAC)
2.8. Statistical Analysis
3. Results and Discussion
3.1. Soil Analysis
3.2. Changes in Photosynthetic Pigments
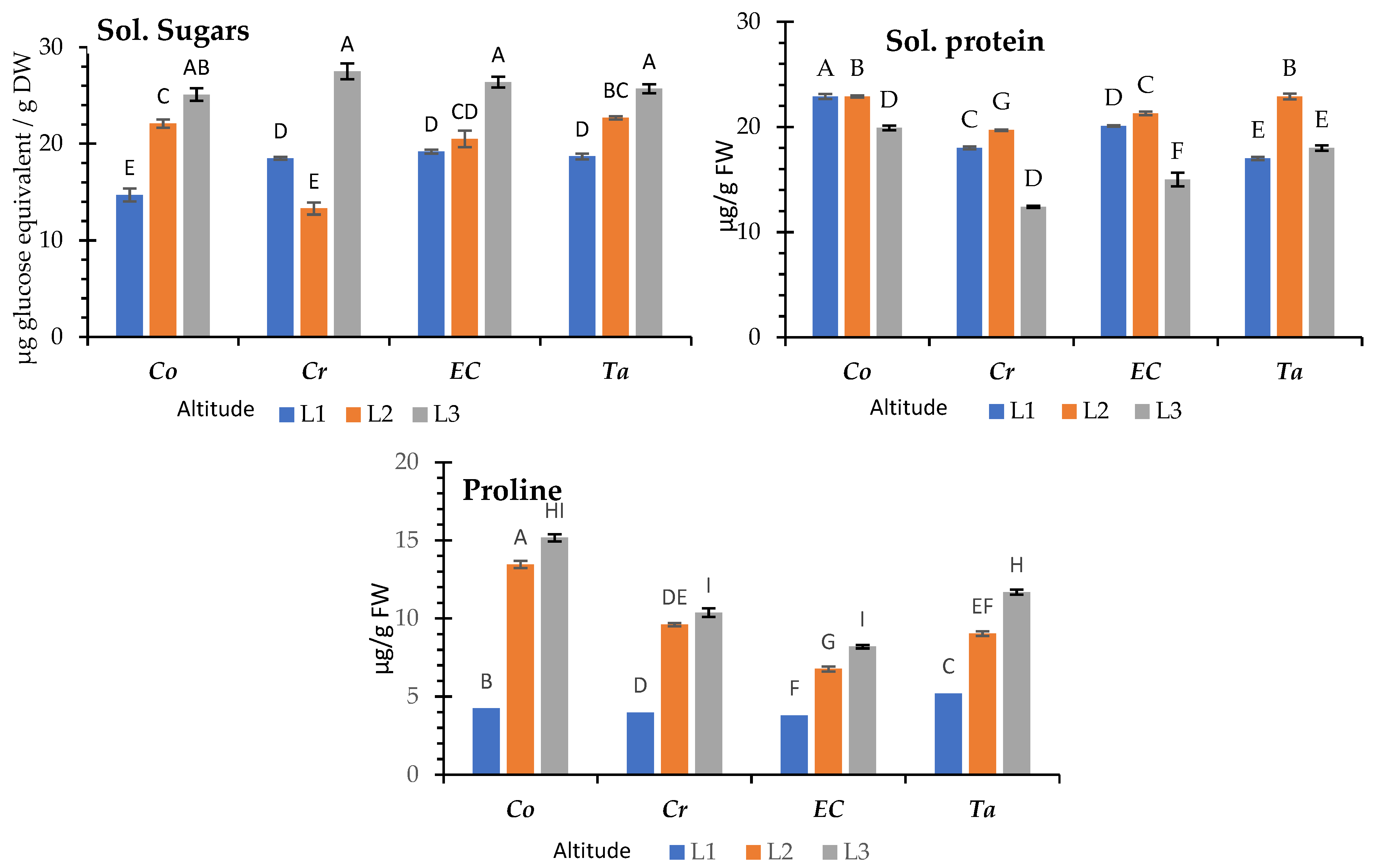
3.3. Variation in Total Soluble Sugar, Total Soluble Proteins, and Proline (Osmolytes)
3.4. Enzymatic Antioxidant System
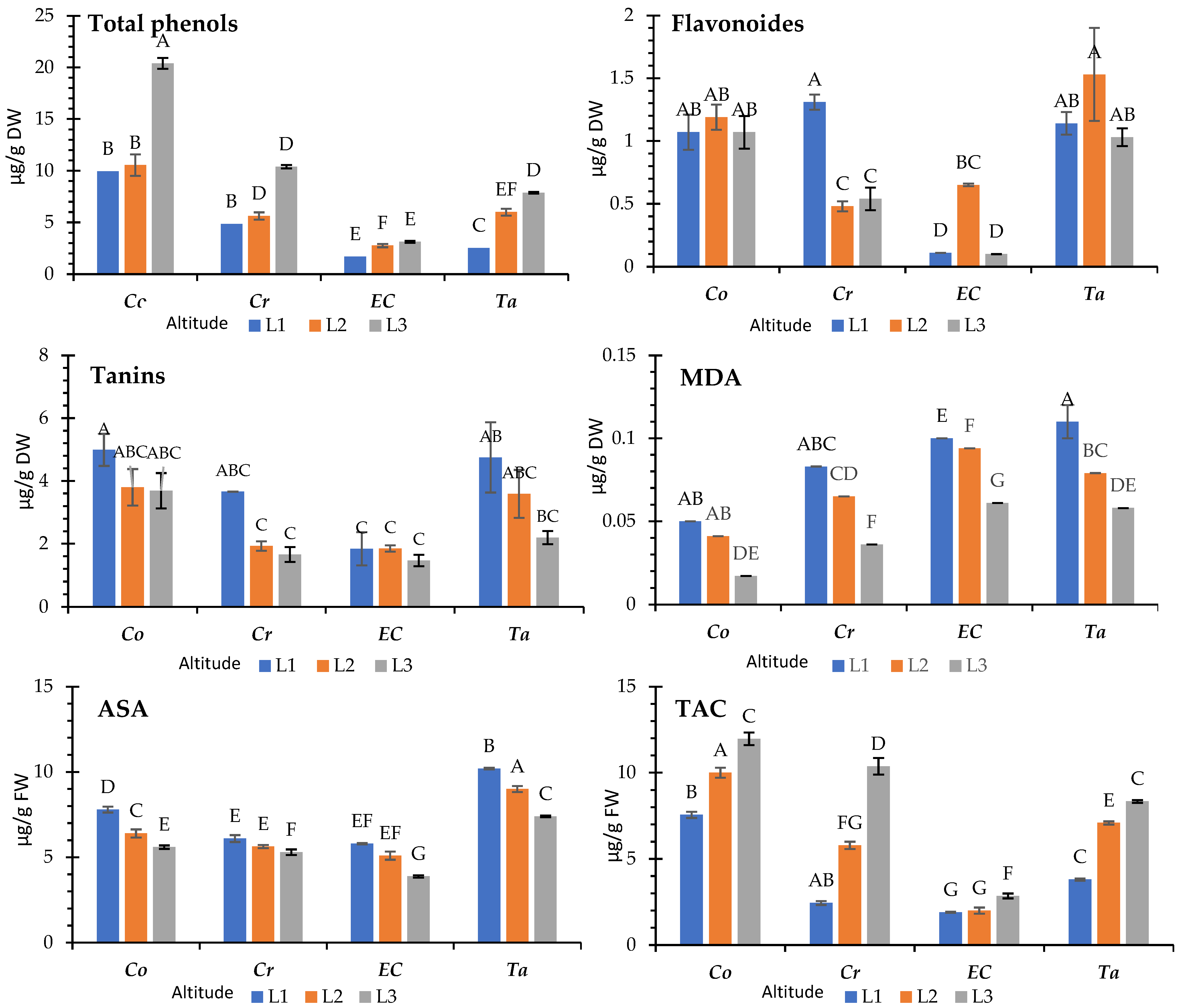
3.5. Non-Enzymatic Antioxidant Systems
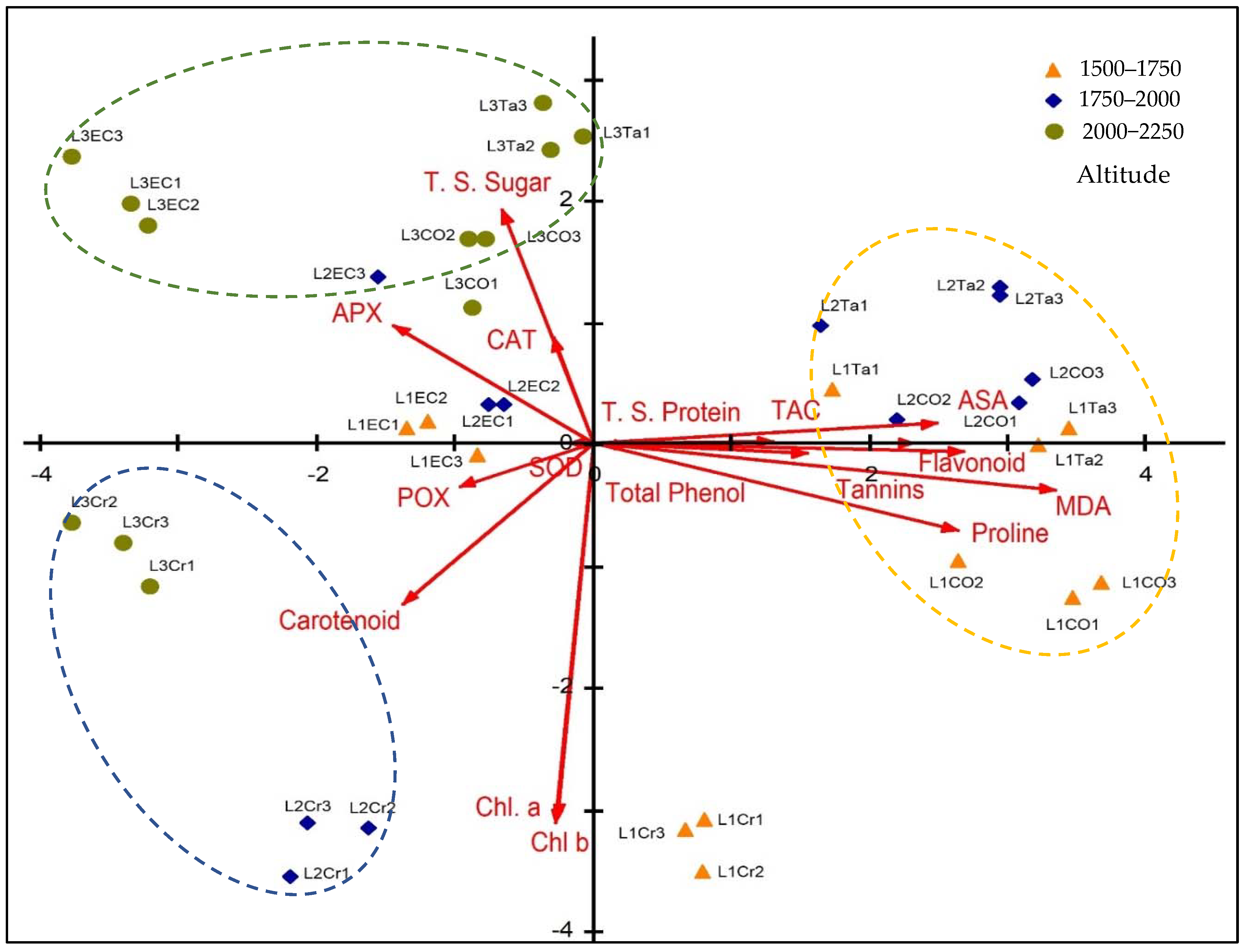
3.6. Principal Component Analysis (PCA)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Devi, N.; Hagedorn, F.; Moiseev, P.; Bugmann, H.; Shiyatov, S.; Mazepa, V.; Rigling, A. Expanding Forests and Changing Growth Forms of Siberian Larch at the Polar Urals Treeline during the 20th Century: TREELINE CHANGES IN THE POLAR URALS. Glob. Change Biol. 2008, 14, 1581–1591. [Google Scholar] [CrossRef]
- Okunlola, G.O.; Olatunji, O.A.; Akinwale, R.O.; Tariq, A.; Adelusi, A.A. Physiological Response of the Three Most Cultivated Pepper Species (Capsicum spp.) in Africa to Drought Stress Imposed at Three Stages of Growth and Development. Sci. Hortic. 2017, 224, 198–205. [Google Scholar] [CrossRef]
- Qian, C.; Yin, H.; Shi, Y.; Zhao, J.; Yin, C.; Luo, W.; Dong, Z.; Chen, G.; Yan, X.; Wang, X.-R.; et al. Population Dynamics of Agriophyllum Squarrosum, a Pioneer Annual Plant Endemic to Mobile Sand Dunes, in Response to Global Climate Change. Sci. Rep. 2016, 6, 26613. [Google Scholar] [CrossRef] [Green Version]
- Jakada, B.H.; Aslam, M.; Fakher, B.; Greaves, J.G.; Li, Z.; Li, W.; Lai, L.; Ayoade, O.A.; Cheng, Y.; Cao, S.; et al. Identification of SWI2/SNF2-Related 1 Chromatin Remodeling Complex (SWR1-C) Subunits in Pineapple and the Role of Pineapple SWR1 COMPLEX 6 (AcSWC6) in Biotic and Abiotic Stress Response. Biomolecules 2019, 9, 364. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Guo, C.; Ahmad, S.; Wang, Q.; Yu, J.; Liu, C.; Guo, Y. Systematic Analysis of MYB Family Genes in Potato and Their Multiple Roles in Development and Stress Responses. Biomolecules 2019, 9, 317. [Google Scholar] [CrossRef] [Green Version]
- ErYuan, L.; YaFeng, W.; Yan, X.; Bo, L.; XueMei, S. Growth Variation in Abies Georgei Var. Smithii along Altitudinal Gradients in the Sygera Mountains, Southeastern Tibetan Plateau. Trees Struct. Funct. 2010, 24, 363–373. [Google Scholar]
- Najafabadi, M.; Ehsanzadeh, P. Photosynthetic and Antioxidative Upregulation in Drought-Stressed Sesame (Sesamum Indicum L.) Subjected to Foliar-Applied Salicylic Acid. Photosynthetica 2017, 55, 611–622. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [Green Version]
- Boulos, L. Flora of Egypt Checklist, Revised Annotated Edition; Al-Hadara Publishing: Cairo, Egypt, 2009; pp. 198–201. [Google Scholar]
- El-Wahab, R.A.; Zaghloul, M.S.; Moustafa, A.A. Conservation of Medicinal Plants in St. Catherine Protectorate, South Sinai, Egypt. In Proceedings of the First International Conference on Strategy of Egyptian Herbaria, Giza, Egypt, 9–11 March 2004; pp. 231–251. [Google Scholar]
- Rabei, S.; Khedr, A.; Elgamal, I. Floristic Study of Saint Katherine Protectorate, Sinai: With One New Record to Flora of Egypt. Taeckholmia 2021, 41, 32. [Google Scholar] [CrossRef]
- Omar, K.; Elgamal, I. Assess the Extinction Risk of Mountain Endemic Plants in Egypt Under the Current Climatic Condition: A Case Study of Endemic Silene Species. Eur. J. Biol. Biotechnol. 2021, 2, 34–47. [Google Scholar] [CrossRef]
- Fakhry, A.M.; El-Keblawy, A.; Shabana, H.A.; Gamal, I.E.; Shalouf, A. Microhabitats Affect Population Size and Plant Vigor of Three Critically Endangered Endemic Plants in Southern Sinai Mountains, Egypt. Land 2019, 8, 86. [Google Scholar] [CrossRef]
- Shaltout, K.H.; Ahmed, D.A.; Shabana, H.A. Population Structure and Dynamics of the Endemic Species Phlomis Aurea Decne in Different Habitats in Southern Sinai Peninsula, Egypt. Glob. Ecol. Conserv. 2015, 4, 505–515. [Google Scholar] [CrossRef] [Green Version]
- El-Keblawy, A.A.; Khedr, A.-H.A. Population Structure and Ecological Role of Moringa Peregrina (Forssk.) Fiori. at Its Northwestern Range Edge in the Hajar Mountains. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2015, 151, 29–38. [Google Scholar] [CrossRef]
- Elkelish, A.A.; Alhaithloul, H.A.S.; Qari, S.H.; Soliman, M.H.; Hasanuzzaman, M. Pretreatment with Trichoderma Harzianum Alleviates Waterlogging-Induced Growth Alterations in Tomato Seedlings by Modulating Physiological, Biochemical, and Molecular Mechanisms. Environ. Exp. Bot. 2020, 171, 103946. [Google Scholar] [CrossRef]
- Hashim, A.M.; Alharbi, B.M.; Abdulmajeed, A.M.; Elkelish, A.; Hozzein, W.N.; Hassan, H.M. Oxidative Stress Responses of Some Endemic Plants to High Altitudes by Intensifying Antioxidants and Secondary Metabolites Content. Plants 2020, 9, 869. [Google Scholar] [CrossRef]
- Wonsick, M.M.; Pinker, R.T. The Radiative Environment of the Tibetan Plateau. Int. J. Climatol. 2014, 34, 2153–2162. [Google Scholar] [CrossRef]
- Harborne, J.B. Classes and Functions of Secondary Products from Plants. In Chemicals from Plants; Institute of Food Research: Norwich, UK, 1999; pp. 1–25. ISBN 978-981-02-2773-9. [Google Scholar]
- Nawrot-Chorabik, K.; Sułkowska, M.; Gumulak, N. Secondary Metabolites Produced by Trees and Fungi: Achievements So Far and Challenges Remaining. Forests 2022, 13, 1338. [Google Scholar] [CrossRef]
- Julkunen-Tiitto, R.; Rousi, M.; Bryant, J.; Sorsa, S.; Keinänen, M.; Sikanen, H. Chemical Diversity of Several Betulaceae Species: Comparison of Phenolics and Terpenoids in Northern Birch Stems. Trees 1996, 11, 16. [Google Scholar] [CrossRef]
- Gong, J.; Zhang, Z.; Zhang, C.; Zhang, J.; Ran, A. Ecophysiological Responses of Three Tree Species to a High-Altitude Environment in the Southeastern Tibetan Plateau. Forests 2018, 9, 48. [Google Scholar] [CrossRef] [Green Version]
- Gea-Izquierdo, G.; Fonti, P.; Cherubini, P.; Martin-Benito, D.; Chaar, H.; Canellas, I. Xylem Hydraulic Adjustment and Growth Response of Quercus Canariensis Willd. to Climatic Variability. Tree Physiol. 2012, 32, 401–413. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gao, S.; He, X.; Li, Y.; Li, P.; Zhang, Y.; Chen, W. Growth, Secondary Metabolites and Enzyme Activity Responses of Two Edible Fern Species to Drought Stress and Rehydration in Northeast China. Agronomy 2019, 9, 137. [Google Scholar] [CrossRef]
- Soliman, M.H.; Alayafi, A.A.M.; El Kelish, A.A.; Abu-Elsaoud, A.M. Acetylsalicylic Acid Enhance Tolerance of Phaseolus Vulgaris L. to Chilling Stress, Improving Photosynthesis, Antioxidants and Expression of Cold Stress Responsive Genes. Bot. Stud. 2018, 59, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleem, M.H.; Ali, S.; Rehman, M.; Rana, M.S.; Rizwan, M.; Kamran, M.; Imran, M.; Riaz, M.; Soliman, M.H.; Elkelish, A.; et al. Influence of Phosphorus on Copper Phytoextraction via Modulating Cellular Organelles in Two Jute (Corchorus Capsularis L.) Varieties Grown in a Copper Mining Soil of Hubei Province, China. Chemosphere 2020, 248, 126032. [Google Scholar] [CrossRef]
- Elkelish, A.; Qari, S.H.; Mazrou, Y.S.A.; Abdelaal, K.A.A.; Hafez, Y.M.; Abu-Elsaoud, A.M.; Batiha, G.E.-S.; El-Esawi, M.A.; El Nahhas, N. Exogenous Ascorbic Acid Induced Chilling Tolerance in Tomato Plants Through Modulating Metabolism, Osmolytes, Antioxidants, and Transcriptional Regulation of Catalase and Heat Shock Proteins. Plants 2020, 9, 431. [Google Scholar] [CrossRef] [Green Version]
- Racchi, M. Antioxidant Defenses in Plants with Attention to Prunus and Citrus spp. Antioxidants 2013, 2, 340–369. [Google Scholar] [CrossRef]
- Keleş, Y.; Everest, A. Relation to Altitude Adaptation and Antioxidant Defence System in Five Shrubs and Trees Species from Middle Taurus Mountains. Int. J. Nat. Eng. Sci. 2008, 2, 45–49. [Google Scholar]
- Catola, S.; Marino, G.; Emiliani, G.; Huseynova, T.; Musayev, M.; Akparov, Z.; Maserti, B.E. Physiological and Metabolomic Analysis of Punica Granatum (L.) under Drought Stress. Planta 2016, 243, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Selmar, D.; Kleinwächter, M. Influencing the Product Quality by Deliberately Applying Drought Stress during the Cultivation of Medicinal Plants. Ind. Crops Prod. 2013, 42, 558–566. [Google Scholar] [CrossRef]
- Cheng, L.; Han, M.; Yang, L.; Li, Y.; Sun, Z.; Zhang, T. Changes in the Physiological Characteristics and Baicalin Biosynthesis Metabolism of Scutellaria Baicalensis Georgi under Drought Stress. Ind. Crops Prod. 2018, 122, 473–482. [Google Scholar] [CrossRef]
- Das, K.; Roychoudhury, A. Reactive Oxygen Species (ROS) and Response of Antioxidants as ROS-Scavengers during Environmental Stress in Plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Bozin, B.; Mimica-Dukic, N.; Samojlik, I.; Goran, A.; Igic, R. Phenolics as Antioxidants in Garlic (Allium Sativum L., Alliaceae). Food Chem. 2008, 111, 925–929. [Google Scholar] [CrossRef]
- Moussa, Z.; Judeh, Z.M.A.; Ahmed, S.A.; Moussa, Z.; Judeh, Z.M.A.; Ahmed, S.A. Nonenzymatic Exogenous and Endogenous Antioxidants; IntechOpen: London, UK, 2019; ISBN 978-1-78985-144-1. [Google Scholar]
- Rahimi, Y.; Taleei, A.; Ranjbar, M. Long-Term Water Deficit Modulates Antioxidant Capacity of Peppermint (Mentha Piperita L.). Sci. Hortic. 2018, 237, 36–43. [Google Scholar] [CrossRef]
- Khan, M.N.; Zhang, J.; Luo, T.; Liu, J.; Ni, F.; Rizwan, M.; Fahad, S.; Hu, L. Morpho-Physiological and Biochemical Responses of Tolerant and Sensitive Rapeseed Cultivars to Drought Stress during Early Seedling Growth Stage. Acta Physiol. Plant. 2019, 41, 1–13. [Google Scholar] [CrossRef]
- Oral, O.; Kutlu, T.; Aksoy, E.; Fıçıcıoğlu, C.; Uslu, H.; Tuğrul, S. The Effects of Oxidative Stress on Outcomes of Assisted Reproductive Techniques. J. Assist. Reprod. Genet. 2006, 23, 81–85. [Google Scholar] [CrossRef]
- Courrau, J. Evaluating Effectivenes: A Framework for Assessing Management Effectiveness of Protected Areas. Available online: https://www.iucn.org/resources/publication/evaluating-effectiveness-framework-assessing-management-effectiveness (accessed on 4 November 2022).
- Moustafa, A.E.-R.A.; Zaghloul, M.S. Environment and Vegetation in the Montane Saint Catherine Area, South Sinai, Egypt. J. Arid Environ. 1996, 34, 331–349. [Google Scholar] [CrossRef]
- Moustafa, A.G. Long Term Monitoring of Rosa Arabica Populations as a Threatened Species in South Sinai, Egypt. Available online: https://www.semanticscholar.org/paper/Long-Term-Monitoring-of-Rosa-arabica-Populations-as-Moustafa/a9209f600f5eef6928a0df021c7866cea10018f1 (accessed on 2 April 2020).
- Moustafa, A.R.A.; Klopatek, J.M. Vegetation and Landforms of the Saint Catherine Area, Southern Sinai, Egypt. J. Arid Environ. 1995, 30, 385–395. [Google Scholar] [CrossRef]
- Danin, A. Desert Rocks—A Habitat Which Supports Many Species That Were New to Science in the Last 40 Years. Turk. J. Bot. 2008, 32, 459–464. [Google Scholar]
- Grainger, J. ‘People Are Living in the Park’. Linking Biodiversity Conservation to Community Development in the Middle East Region: A Case Study from the Saint Katherine Protectorate, Southern Sinai. J. Arid Environ. 2003, 54, 29–38. [Google Scholar] [CrossRef]
- Täckholm, V. Students’ Flora of Egypt, 2nd ed.; Cairo University: Beirut, Lebanon, 1974. [Google Scholar]
- Zohary, M. Flora Palaestina; Israel Academy of Science and Humanities: Jerusalem, Israel, 1966. [Google Scholar]
- POWO Plants of the World Online. Kew Science. Available online: https://powo.science.kew.org/ (accessed on 1 November 2022).
- Boulos, L. Medicinal Plants of North Africa; Reference Publications, Inc.: Algonac, MI, USA, 1983. [Google Scholar]
- El-Hadidi, M.N.; Batanouny, K.H.; Fahmy, A.G. The Egyptian Plant Red Data Book; University of Cairo: Giza, Egypt, 1991. [Google Scholar]
- El-Mousallamy, A.M.; Hussein, S.A.; Merfort, I.; Nawwar, M.A. Unusual Phenolic Glycosides from Cotoneaster Orbicularis. Phytochemistry 2000, 53, 699–704. [Google Scholar] [CrossRef]
- Moustafa, A.; Zaghloul, M.; Mansour, S.; Alotaibi, M. Conservation Strategy for Protecting Crataegus x Sinaica against Climate Change and Anthropologic Activities in South Sinai Mountains, Egypt. Catrina Int. J. Environ. Sci. 2019, 18, 1–6. [Google Scholar] [CrossRef]
- Shaltout, K.H.; Eid, E.M.; Al-Sodany, Y.M.; Heneidy, S.Z.; Shaltout, S.K.; El-Masry, S.A. Effect of Protection of Mountainous Vegetation against Over-Grazing and Over-Cutting in South Sinai, Egypt. Diversity 2021, 13, 113. [Google Scholar] [CrossRef]
- Refaat, A.T.; Shahat, A.A.; Ehsan, N.A.; Yassin, N.; Hammouda, F.; Tabl, E.A.; Ismail, S.I. Phytochemical and Biological Activities of Crataegus Sinaica Growing in Egypt. Asian Pac. J. Trop. Med. 2010, 3, 257–261. [Google Scholar] [CrossRef] [Green Version]
- Murch, S.J.; Wierenga, E.J.; El-Demerdash, M.A.; Saxena, P.K. In Vitro Propagation of the Egyptian Medicinal Plant, Echinops Spinosissimus Turra. Plant Cell Tissue Organ Cult. 2003, 74, 81–86. [Google Scholar] [CrossRef]
- Dawidar, A.M.; Metwally, M.A.; Abou-Elzahab, M.; Abdel-Mogib, M. Sesquiterpene Lactones from Echinops Spinosissimus. Pharmazie 1990, 45, 70–71. [Google Scholar]
- Hegazy, M.-E.F.; Hamed, A.R.; Mohamed, T.A.; Debbab, A.; Nakamura, S.; Matsuda, H.; Paré, P.W. Anti-Inflammatory Sesquiterpenes from the Medicinal Herb Tanacetum Sinaicum. RSC Adv. 2015, 5, 44895–44901. [Google Scholar] [CrossRef]
- El-Dahmy, S.I.; Abdel-Ghani, A.E.; Attia, R.A.; Abouelenein, D.D. Chemical Composition and Biological Activities of the Essential Oil of Tanacetum Sinaicum Del. Grown Egypt. J. Pharmacogn. Phytochem. 2019, 8, 326–331. [Google Scholar]
- Estefan, G. Methods of Soil, Plant, and Water Analysis: A Manual for the West Asia and North Africa Region, 3rd ed.; International Center for Agricultural Research in the Dry Areas (ICARDA): Beirut, Lebanon, 2017. [Google Scholar]
- Dewis, J.; Nations, F.; Freitas, F. Physical and Chemical Methods of Soil and Water Analysis; Food and Agriculture Organization of the United Nations. FAO Soils Bull. 1970, 10, 275. [Google Scholar]
- Johnson, C.M.; Ulrich, A. Analytical Methods for Use in Plant Analysis; University of California: Los Angeles, CA, USA, 1959. [Google Scholar]
- Shapiro, L.; Brannock, W.W. A Field Method for the Determination of Calcium and Magnesium in Limestone and Dolomite; Open-File Report; U.S. Geological Survey: Reston, VA, USA, 1957; Volume 57–99, p. 9. [CrossRef]
- Blume, H.-P. Page, A. L., R. H. Miller and D. R. Keeney (Ed., 1982): Methods of soil analysis; 2. Chemical and microbiological properties, 2. Aufl. 1184 S., American Soc. of Agronomy (Publ.), Madison, Wisconsin, USA, gebunden 36 Dollar. Bodenkd 1985, 148, 363–364. [Google Scholar] [CrossRef]
- Black, C.A. American Society of Agronomy. American Society for Testing and Materials Methods of Soil Analysis Part 1, Part 1; American Society of Agronomy: Madison, WI, USA, 1965. [Google Scholar]
- Piper, C.S. Soil and Plant Analysis. Soil Sci. 1945, 59, 263. [Google Scholar] [CrossRef]
- Metzner, H.; Rau, H.; Senger, H. Untersuchungen zur Synchronisierbarkeit einzelner Pigmentmangel-Mutanten von Chlorella. Planta 1965, 65, 186–194. [Google Scholar] [CrossRef]
- Prud’homme, M.-P.; Gonzalez, B.; Billard, J.-P.; Boucaud, J. Carbohydrate Content, Fructan and Sucrose Enzyme Activities in Roots, Stubble and Leaves of Ryegrass (Lolium Perenne L.) as Affected by Source/Sink Modification after Cutting. J. Plant Physiol. 1992, 140, 282–291. [Google Scholar] [CrossRef]
- Loewus, F.A. Improvement in Anthrone Method for Determination of Carbohydrates. Anal. Chem. 1952, 24, 219. [Google Scholar] [CrossRef]
- Malik, C.P.; Singh, M.B. Plant Enzymology and Histo-Enzymology: A Text Manual; Kalyani Publishers: Delhi, India, 1980. [Google Scholar]
- Harborne, A.J. Phytochemical Methods A Guide to Modern Techniques of Plant Analysis, 3rd ed.; Springer: Dordrecht, The Netherlands, 1998; ISBN 978-0-412-57260-9. [Google Scholar]
- Ejikeme, C.M.; Ezeonu, C.S.; Eboatu, A.N. Determination of Physical and Phytochemical Constituents of Some Tropical Timbers Indigenous to Niger Delta Area of Nigeria. Eur. Sci. J. ESJ 2014, 10, 247–270. [Google Scholar]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts: I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.P.; Choudhuri, M.A. Implications of Water Stress-Induced Changes in the Levels of Endogenous Ascorbic Acid and Hydrogen Peroxide in Vigna Seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation, and Decreased Levels of Superoxide Dismutase and Catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Hermans, C.; Conn, S.J.; Chen, J.; Xiao, Q.; Verbruggen, N. An Update on Magnesium Homeostasis Mechanisms in Plants. Metallomics 2013, 5, 1170–1183. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, Peroxidase, and Polyphenoloxidase Activities during Rice Leaf Senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Koricheva, J.; Roy, S.; Vranjic, J.A.; Haukioja, E.; Hughes, P.R.; Hänninen, O. Antioxidant Responses to Simulated Acid Rain and Heavy Metal Deposition in Birch Seedlings. Environ. Pollut. Barking Essex 1997, 95, 249–258. [Google Scholar] [CrossRef]
- Kampfenkel, K.; Montagu, M.V.; Inze, D. Effects of Iron Excess on Nicotiana Plumbaginifolia Plants (Implications to Oxidative Stress). Plant Physiol. 1995, 107, 725–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric Quantitation of Antioxidant Capacity through the Formation of a Phosphomolybdenum Complex: Specific Application to the Determination of Vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Wahab, R.; Zayed, A.E.-M.; Moustafa, A.E.-R.; Klopatek, J.; Helmy, M. Landforms, Vegetation, and Soil Quality in South Sinai, Egypt. Catrina Int. J. Environ. Sci. 2018, 1, 127–138. [Google Scholar]
- Sharma, P.; Rana, J.C.; Devi, U.; Randhawa, S.S.; Kumar, R. Floristic Diversity and Distribution Pattern of Plant Communities along Altitudinal Gradient in Sangla Valley, Northwest Himalaya. Sci. World J. 2014, 2014, e264878. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Nazim, H.; Liang, Z.; Yang, D. Magnesium Deficiency in Plants: An Urgent Problem. Crop J. 2016, 4, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Salama, F.; Abd El-Ghani, M.; Gadalla, M.; Ramadan, T.; Galal, H.; Gaafar, A. Vegetation Patterns and Floristic Composition along Elevation Gradient on Jabal Musa, South Sinai, Egypt. Catrina Int. J. Environ. Sci. 2018, 17, 41–57. [Google Scholar] [CrossRef]
- Mirzakhaninafchi, H.; Mani, I.; Hasan, M.; Nafchi, A.M.; Parray, R.A.; Kumar, D. Development of Prediction Models for Soil Nitrogen Management Based on Electrical Conductivity and Moisture Content. Sensors 2022, 22, 6728. [Google Scholar] [CrossRef]
- Visconti, F.; de Paz, J.M. Electrical Conductivity Measurements in Agriculture: The Assessment of Soil Salinity. In New Trends and Developments in Metrology; Cocco, L., Ed.; IntechOpen: London, UK, 2016; ISBN 978-953-51-2477-1. [Google Scholar]
- Rhoades, J.D.; Chanduvi, F.; Lesch, S. Soil Salinity Assessment: Methods and Interpretation of Electrical Conductivity Measurements; Food & Agriculture Org: Rome, Italy, 1999. [Google Scholar]
- Zhao, Y.; Han, Q.; Ding, C.; Huang, Y.; Liao, J.; Chen, T.; Feng, S.; Zhou, L.; Zhang, Z.; Chen, Y.; et al. Effect of Low Temperature on Chlorophyll Biosynthesis and Chloroplast Biogenesis of Rice Seedlings during Greening. Int. J. Mol. Sci. 2020, 21, 1390. [Google Scholar] [CrossRef] [Green Version]
- Cui, G.; Li, B.; He, W.; Yin, X.; Liu, S.; Lian, L.; Zhang, Y.; Liang, W.; Zhang, P. Physiological Analysis of the Effect of Altitudinal Gradients on Leymus Secalinus on the Qinghai-Tibetan Plateau. PLoS ONE 2018, 13, e0202881. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, T.B.; Ribas, A.F.; de Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological Responses to Drought, Salinity, and Heat Stress in Plants: A Review. Stresses 2022, 2, 113–135. [Google Scholar] [CrossRef]
- Abd El-Ghani, M.M.; Huerta-Martínez, F.M.; Hongyan, L.; Qureshi, R. Erratum to: Plant Responses to Hyperarid Desert Environments. In Plant Responses to Hyperarid Desert Environments; Abd El-Ghani, M.M., Huerta-Martínez, F.M., Hongyan, L., Qureshi, R., Eds.; Springer International Publishing: Cham, Germany, 2017; p. e1. ISBN 978-3-319-59135-3. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Caverzan, A.; Casassola, A.; Patussi Brammer, S. Reactive Oxygen Species and Antioxidant Enzymes Involved in Plant Tolerance to Stress. In Abiotic and Biotic Stress in Plants-Recent Advances and Future Perspectives; Shanker, A.K., Shanker, C., Eds.; IntechOpen: London, UK, 2016; ISBN 978-953-51-2250-0. [Google Scholar]
- Havaux, M. Carotenoid Oxidation Products as Stress Signals in Plants. Plant J. Cell Mol. Biol. 2014, 79, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Uarrota, V.G.; Stefen, D.L.V.; Leolato, L.S.; Gindri, D.M.; Nerling, D. Revisiting Carotenoids and Their Role in Plant Stress Responses: From Biosynthesis to Plant Signaling Mechanisms during Stress. In Antioxidants and Antioxidant Enzymes in Higher Plants; Springer International Publishing: Cham, Germany, 2018; pp. 207–232. ISBN 978-3-319-75087-3. [Google Scholar]
- Sharma, A.; Kumar, V.; Yuan, H.; Kanwar, M.K.; Bhardwaj, R.; Thukral, A.K.; Zheng, B. Jasmonic Acid Seed Treatment Stimulates Insecticide Detoxification in Brassica Juncea L. Front. Plant Sci. 2018, 9, 1609. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Jiang, J.-G. Osmotic Adjustment and Plant Adaptation to Environmental Changes Related to Drought and Salinity. Environ. Rev. 2010, 18, 309–319. [Google Scholar] [CrossRef]
- Kurutas, E.B. The Importance of Antioxidants Which Play the Role in Cellular Response against Oxidative/Nitrosative Stress: Current State. Nutr. J. 2015, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- Lambers, H.; Oliveira, R.S. Plant Physiological Ecology; Springer International Publishing: Cham, Germany, 2019; ISBN 978-3-030-29638-4. [Google Scholar]
- Guidi, L.; Tattini, M.; Landi, M. How Does Chloroplast Protect Chlorophyll Against Excessive Light? In Chlorophyll; Jacob-Lopes, E., Zepka, L.Q., Queiroz, M.I., Eds.; IntechOpen: London, UK, 2017; ISBN 978-953-51-3107-6. [Google Scholar]
- Kavi Kishor, P.B.; Suravajhala, P.; Rathnagiri, P.; Sreenivasulu, N. Intriguing Role of Proline in Redox Potential Conferring High Temperature Stress Tolerance. Front. Plant Sci. 2022, 13, 867531. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Ghorbani Javid, M.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of Exogenous Proline to Abiotic Stresses Tolerance in Plants: A Review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A Multifunctional Amino Acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Fu, Y.; Ma, H.; Chen, S.; Gu, T.; Gong, J. Control of Proline Accumulation under Drought via a Novel Pathway Comprising the Histone Methylase CAU1 and the Transcription Factor ANAC055. J. Exp. Bot. 2018, 69, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Hakim, M.A.; Juraimi, A.S.; Hanafi, M.M.; Ismail, M.R.; Selamat, A.; Rafii, M.Y.; Latif, M.A. Biochemical and Anatomical Changes and Yield Reduction in Rice (Oryza Sativa L.) under Varied Salinity Regimes. BioMed Res. Int. 2014, 2014, e208584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, J.; Bernal, A.; Pomar, F.; Merino, F. Induction of Shikimate Dehydrogenase and Peroxidase in Pepper (Capsicum Annuum L.) Seedlings in Response to Copper Stress and Its Relation to Lignification. Plant Sci. 2001, 161, 179–188. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First Line Defence Antioxidants-Superoxide Dismutase (SOD), Catalase (CAT) and Glutathione Peroxidase (GPX): Their Fundamental Role in the Entire Antioxidant Defence Grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Mhamdi, A.; Noctor, G.; Baker, A. Plant Catalases: Peroxisomal Redox Guardians. Arch. Biochem. Biophys. 2012, 525, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.-P.; Xiong, Z.-T.; Huang, Y.; Li, M.-J. Cadmium-Induced Changes in Pigments, Total Phenolics, and Phenylalanine Ammonia-Lyase Activity in Fronds of Azolla Imbricata. Environ. Toxicol. 2006, 21, 505–512. [Google Scholar] [CrossRef]
- Krol, A.; Amarowicz, R.; Weidner, S. The Effects of Cold Stress on the Phenolic Compounds and Antioxidant Capacity of Grapevine (Vitis Vinifera L.) Leaves. J. Plant Physiol. 2015, 189, 97–104. [Google Scholar] [CrossRef]
- Koh, Y.S.; Wong, S.K.; Ismail, N.H.; Zengin, G.; Duangjai, A.; Saokaew, S.; Phisalprapa, P.; Tan, K.W.; Goh, B.H.; Tang, S.Y. Mitigation of Environmental Stress-Impacts in Plants: Role of Sole and Combinatory Exogenous Application of Glutathione. Front. Plant Sci. 2021, 12, 791205. [Google Scholar] [CrossRef]
- Ozkur, O.; Ozdemir, F.; Bor, M.; Turkan, I. Physiochemical and Antioxidant Responses of the Perennial Xerophyte Capparis Ovata Desf. to Drought. Environ. Exp. Bot. 2009, 66, 487–492. [Google Scholar] [CrossRef]








| (a) | ||||||||
| Species | pH | E.C. (ds/m) | TDS | O.C. (%) | O.M. (%) | Clay% | Sand% | Silt% |
| L1 CO. | 7.43 ± 0.07 A | 0.94 ± 0.18 A | 408 ± 113 A | 1.23 ± 0.16 A | 2.12 ± 0.28 A | 8.48 ± 0.28 AB | 39.82 ± 4.89 ABC | 51.71 ± 5.18 AB |
| L1 Cr. | 7.53 ± 0.03 A | 1.03 ± 0.18 A | 657 ± 115 A | 0.83 ± 0.13 A | 1.43 ± 0.22 A | 10.65 ± 1.58 AB | 53.64 ± 4.4 ABC | 35.71 ± 3.23 ABC |
| L1 EC. | 7.57 ± 0.09 A | 0.89 ± 0.15 A | 569.6 ± 97.9 A | 1.01 ± 0.19 A | 1.75 ± 0.32 A | 16.31 ± 4.04 AB | 27.02 ± 5.01 C | 56.66 ± 3.41 A |
| L1 Ta. | 7.5 ± 0 A | 0.9 ± 0.11 A | 576.9 ± 71.4 A | 1.21 ± 0.12 A | 2.09 ± 0.2 A | 20.26 ± 4.98 A | 37.66 ± 4.4 BC | 42.08 ± 0.75 ABC |
| L2 CO. | 7.53 ± 0.03 A | 0.88 ± 0.05 A | 501.5 ± 50.6 A | 0.76 ± 0.03 A | 1.31 ± 0.05 A | 7.35 ± 1.33 AB | 64.17 ± 5.41 AB | 28.48 ± 6.33 BC |
| L2 Cr. | 7.57 ± 0.03 A | 0.72 ± 0.04 A | 496 ± 53.1 A | 1.05 ± 0.19 A | 1.81 ± 0.32 A | 8.69 ± 2.98 AB | 61.2 ± 11.7 AB | 30.15 ± 8.76 BC |
| L2 EC. | 7.47 ± 0.03 A | 0.82 ± 0.14 A | 588.4 ± 90.3 A | 0.98 ± 0.05 A | 1.69 ± 0.09 A | 6.93 ± 0.74 AB | 58.69 ± 6.9 AB | 34.38 ± 6.16 ABC |
| L2 Ta. | 7.5 ± 0 A | 0.71 ± 0.04 A | 484.7 ± 23.5 A | 0.93 ± 0.15 A | 1.61 ± 0.25 A | 5.91 ± 0.21 AB | 62.22 ± 1.65 AB | 31.87 ± 1.44 ABC |
| L3 CO. | 7.6 ± 0 A | 0.67 ± 0.04 A | 437.5 ± 24.9 A | 0.87 ± 0.04 A | 1.49 ± 0.08 A | 6.37 ± 3.66 AB | 51.25 ± 6.73 ABC | 25.9. ± 3.28 ABC |
| L3 Cr. | 7.47 ± 0.09 A | 0.68 ± 0.01 A | 540.6 ± 37 A | 0.81 ± 0.03 A | 1.4 ± 0.06 A | 6.58 ± 3.9 AB | 65.22 ± 7.9 AB | 25.2 ± 4.47 C |
| L3 EC. | 7.53 ± 0.03 A | 0.6.9 ± 0.1 A | 533.8 ± 67 A | 0.85 ± 0.08 A | 1.47 ± 0.14 A | 4.03 ± 0.76 B | 65.78 ± 2.93 AB | 26.19 ± 2.2 BC |
| L3 Ta. | 7.53 ± 0.07 A | 0.71 ± 0.07 A | 548.1 ± 43.5 A | 0.8 ± 0.12 A | 1.38 ± 0.2 A | 4.19 ± 1.34 B | 71.75 ± 6.67 A | 24.06 ± 5.4 C |
| (b) | ||||||||
| Species in Different locations | CL (meq/L) | CaCO3 (%) | SO4 (Mg/L) | HCO3 (%) | Ca (meq/L) | Mg (meq/L) | Na (meq/L) | K (meq/L) |
| L1 CO. | 2.66 ± 0.1 B | 9.33 ± 1.17 A | 7.2 ± 1.46 BC | 0.61 ± 0 A | 4.42 ± 0.54 A | 2.16 ± 0.52 AB | 0.62 ± 0.09 A | 0.98 ± 0.3 A |
| L1 Cr. | 2.19 ± 0.19 B | 5.5 ± 1.04 A | 8.44 ± 0.33 ABC | 0.61 ± 0 A | 5.04 ± 0.03 A | 1.69 ± 0.06 AB | 0.47 ± 0.06 A | 3.12 ± 1.79 A |
| L1 EC. | 4.66 ± 0.91 A | 9.5 ± 5.57 A | 11.57 ± 2.06 AB | 0.51 ± 0.1 A | 2.83 ± 0.51 A | 3.3 ± 0.9 AB | 0.6 ± 0.24 A | 2.18 ± 1.3 A |
| L1 Ta. | 2.28 ± 0.17 B | 5 ± 1.32 A | 9.56 ± 0.12 ABC | 0.51 ± 0.1 A | 4.42 ± 0.54 A | 1.76 ± 0.04 AB | 0.71 ± 0.1 A | 2.18 ± 0.54 A |
| L2 CO. | 2.66 ± 0.1 B | 4.5 ± 1.5 A | 11.74 ± 0.32 AB | 0.51 ± 0.1 A | 4.95 ± 0.94 A | 2.78 ± 0.49 AB | 0.33 ± 0.03 A | 0.64 ± 0.08 A |
| L2 Cr. | 2.19 ± 0.1 B | 6.5 ± 1.26 A | 8.77 ± 1.49 ABC | 0.61 ± 0 A | 4.48 ± 0.57 A | 1.59 ± 0.05 B | 0.33 ± 0.03 A | 0.77 ± 0.13 A |
| L2 EC. | 3.14 ± 0.17 AB | 6.83 ± 1.09 A | 7.32 ± 1.37 BC | 0.61 ± 0 A | 4.52 ± 0.59 A | 2.74 ± 0.54 AB | 0.42 ± 0.06 A | 1.54 ± 0.29 A |
| L2 Ta. | 2.47 ± 0.34 B | 6.67 ± 1.42 A | 5.03 ± 0.39 C | 0.61 ± 0 A | 3.9 ± 0.56 A | 1.65 ± 0.02 B | 0.53 ± 0.14 A | 0.99 ± 0.04 A |
| L3 CO. | 2.47 ± 0.1 B | 4.67 ± 0.88 A | 10.15 ± 1.48 ABC | 0.61 ± 0 A | 4.42 ± 0.54 A | 1.71 ± 0.04 AB | 0.48 ± 0.12 A | 0.34 ± 0.04 A |
| L3 Cr. | 2.47 ± 0.25 B | 2.33 ± 0.44 A | 13.45 ± 0.43 A | 0.61 ± 0 A | 4.48 ± 0.57 A | 3.26 ± 0.08 A | 0.68 ± 0.2 A | 0.53 ± 0.31 A |
| L3 EC. | 2.85 ± 0.44 AB | 5.83 ± 1.09 A | 9.45 ± 1.4 ABC | 0.61 ± 0 A | 4.43 ± 1.09 A | 1.69 ± 0.06 AB | 1.36 ± 0.92 A | 1.41 ± 0.45 A |
| L3 Ta. | 1.33 ± 0.53 B | 9.33 ± 1.17 A | 12.31 ± 0.66 AB | 0.61 ± 0 A | 4.39 ± 1.05 A | 1.65 ± 0.02 B | 0.41 ± 0.08 A | 1.24 ± 0.37 A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, I.A.; Jabbour, A.A.; Abdulmajeed, A.M.; Elhady, M.E.; Almaroai, Y.A.; Hashim, A.M. Adaptive Responses of Four Medicinal Plants to High Altitude Oxidative Stresses through the Regulation of Antioxidants and Secondary Metabolites. Agronomy 2022, 12, 3032. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12123032
Ibrahim IA, Jabbour AA, Abdulmajeed AM, Elhady ME, Almaroai YA, Hashim AM. Adaptive Responses of Four Medicinal Plants to High Altitude Oxidative Stresses through the Regulation of Antioxidants and Secondary Metabolites. Agronomy. 2022; 12(12):3032. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12123032
Chicago/Turabian StyleIbrahim, Ibrahim A., A. A. Jabbour, Awatif M. Abdulmajeed, Mohamed E. Elhady, Yaser A. Almaroai, and Ahmed M. Hashim. 2022. "Adaptive Responses of Four Medicinal Plants to High Altitude Oxidative Stresses through the Regulation of Antioxidants and Secondary Metabolites" Agronomy 12, no. 12: 3032. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12123032