
Investigation by High-Throughput Sequencing Methods of Microbiota Dynamics in Spontaneous Fermentation of Abruzzo (South Italy) Wines

Abstract
:1. Introduction
2. Materials and Methods
2.1. Alcoholic Fermentation and Sampling
2.2. Chemical Analysis of Must and Wine
2.3. Yeast and Bacteria Cultivation
2.4. DNA Extraction
2.5. Miseq Library Preparation and Illumina Sequencing
2.6. Illumina Data Analysis and Sequences Identification by QIIME2
Statistical Analysis
3. Results and Discussion
3.1. Chemical Composition of the Grape Must and Microbial Evolution of Fermentations
3.2. Microbial Community Dynamic during Montepulciano and Trebbiano Fermentation
3.3. β-Diversity of the Bacterial and Fungal Community
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, H.; Ma, H.; Hao, M.; Pretorius, I.S.; Chen, S. Identification of yeast population dynamics of spontaneous fermentation in Beijing wine region, China. Ann. Microbiol. 2009, 59, 69–76. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Zhang, W.; Yi, H.; Wang, B.; Xiao, J.; Zhou, X.Y.; Xu, J.; Jiang, L.; Shi, X. Microbial community composition and its role in volatile compound formation during the spontaneous fermentation of ice wine made from Vidal grapes. Process Biochem. 2020, 92, 365–377. [Google Scholar] [CrossRef]
- Liu, D.; Legras, J.L.; Zhang, P.; Chen, D.; Howell, K. Diversity and dynamics of fungi during spontaneous fermentations and association with unique aroma profiles in wine. Int. J. Food Microbiol. 2021, 338, 108983. [Google Scholar] [CrossRef]
- Granchi, L.; Ganucci, D.; Messini, A.; Vincenzini, M. Oenological properties of Hanseniaspora osmophila and Kloeckera corticis from wines produced by spontaneous fermentations of normal and dried grapes. FEMS Yeast Res. 2002, 2, 403–407. [Google Scholar]
- Garijo, P.; Gonzalez-Arenzana, L.; Lopez-Alfaro, I.; Garde-Cerdan, T.; Lopez, R.; Santamaria, P.; Gutierrez, A.R. Analysis of grapes and the first stages of the vinification process in wine contamination with Brettanomyces bruxellensis. Eur. Food Res. Technol. 2015, 240, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Zott, K.; Miot-Sertier, C.; Claisse, O.; Lonvaud-Funel, A.; Masneuf-Pomarede, I. Dynamics and diversity of non-Saccharomyces yeasts during the early stages in winemaking. Int. J. Food Microbiol. 2008, 125, 197–203. [Google Scholar] [CrossRef]
- Marzano, M.; Fosso, B.; Manzari, C.; Grieco, F.; Intranuovo, M.; Cozzi, G.; Mulè, G.; Scioscia, G.; Valiente, G.; Tullo, A.; et al. Complexity and dynamics of the winemaking bacterial communities in berries, musts, and wines from apulian grape cultivars through time and space. PLoS ONE 2016, 11, e0157383. [Google Scholar] [CrossRef]
- Costello, P.J.; Siebert, T.E.; Solomon, M.R.; Bartowsky, E.J. Synthesis of fruity ethyl esters by acyl coenzyme A: Alcohol acyltransferase and reverse esterase activities in Oenococcus oeni and Lactobacillus plantarum. J. Appl. Microbiol. 2013, 114, 797–806. [Google Scholar] [CrossRef]
- Guzzon, R.; Roman, T.; Larcher, R. Impact of different temperature profiles on simultaneous yeast and bacteria fermentation. Ann. Microbiol. 2020, 70, 44. [Google Scholar] [CrossRef]
- Guzzon, R.; Malacarne, M.; Larcher, R.; Franciosi, E.; Toffanin, A. The impact of grape processing and carbonic maceration on the microbiota of early stages of winemaking. J. App. Microbiol. 2019, 128, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Yu, L.; Wang, W.; Yan, R.; Zhang, Z.; Yang, H.L.; Zhu, D.; Zhu, B. Comparative Evaluation of Microbiota Dynamics and Metabolites Correlation Between Spontaneous and Inoculated Fermentations of Nanfeng Tangerine Wine. Front. Microbiol. 2014, 12, 649978. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Thorngate, J.H.; Richardson, P.M.; Mills, D.A. Microbial biogeography of wine grapes is conditioned by cultivar, vintage, and climate. Proc. Natl. Acad. Sci. USA 2014, 111, E139–E148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrau, F.; Gaggero, C.; Aguilar, P.S. Yeast diversity and native vigor for flavor phenotypes. Trends Biotechnol. 2015, 33, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Knight, S.; Klaere, S.; Fedrizzi, B.; Goddard, M.R. Regional microbial signatures positively correlate with differential wine phenotypes: Evidence for a microbial aspect to terroir. Sci. Rep. 2015, 5, 14233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hijum, S.A.; Vaughan, E.E.; Vogel, R.F. Application of state-of-art sequencing technologies to indigenous food fermentations. Curr. Opin. Plant Biol. 2013, 24, 178–186. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among wine grape microbiome, metabolome, and fermentation behavior suggest microbial contribution to regional wine characteristics. mBio 2016, 7, e00631-16. [Google Scholar] [CrossRef] [Green Version]
- Stefanini, I.; Albanese, D.; Cavazza, A.; Franciosi, E.; De Filippo, C.; Donati, C.; Cavalieri, D. Dynamic changes in microbiota and mycobiota during spontaneous ‘Vino Santo Trentino’ fermentation. Microb. Biotechnol. 2016, 9, 195–208. [Google Scholar] [CrossRef] [Green Version]
- Belda, I.; Zarraonaindia, I.; Perisin, M.; Palacios, A.; Acedo, A. From vineyard soil to wine fermentation: Microbiome approximations to explain the “terroir” concept. Front. Microbiol. 2017, 8, 821. [Google Scholar] [CrossRef] [Green Version]
- OIV. Update of Compendium of International Methods of Anaysis of Spirit Drinks of Vitiviniculture Origin; Resolution OIV/OENO 381/2009; OIV: Paris, France, 2009. [Google Scholar]
- OIV. Compendium of International Methods of Wine and Must Analysis; OIV: Paris, France, 2022. [Google Scholar]
- Baker, G.C.; Smith, J.J.; Cowan, D.A. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Methods 2003, 55, 541–555. [Google Scholar] [CrossRef] [Green Version]
- Claesson, M.J.; Wang, Q.; O’Sullivan, O.; Greene-Diniz, R.; Cole, J.R.; Ross, R.P.; O’Toole, P.W. Comparison of two next-generation sequencing technologies for resolving highly complex microbiota composition using tandem variable 16S rRNA gene regions. Nucleic Acids Res. 2010, 38, 200. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Vazquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A tool for visualizing high-throughput microbial community data. GigaScience 2013, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Calzarano, F.; Seghetti, L.; Del Carlo, M.; Cichelli, A. Effect of Esca on the Quality of Berries, Musts and Wines. Phytopathol. Mediterr. 2004, 43, 125–135. [Google Scholar]
- Tofalo, R.; Patrignani, F.; Lanciotti, R.; Perpetuini, G.; Schirone, M.; Di Gianvito, P.; Pizzoni, D.; Arfelli, G.; Suzzi, G. Aroma Profile of Montepulciano d’Abruzzo Wine Fermented by Single and Co-culture Starters of Autochthonous Saccharomyces and Non-Saccharomyces Yeasts. Front. Microbiol. 2016, 7, 610. [Google Scholar] [CrossRef]
- Su, Y.; Seguinot, P.; Sanchez, I.; Ortiz-Julien, A.; Heras, J.M.; Querol, A.; Camarasa, C.; Guillamón, J.M. Nitrogen sources preferences of non-Saccharomyces yeasts to sustain growth and fermentation under winemaking conditions. Food Microbiol. 2020, 85, 103287. [Google Scholar] [CrossRef]
- Bell, S.J.; Henschcke, P.A. Implications of nitrogen in grapes, fermentation and wine. Aust. J. Grape Wine Res. 2005, 11, 242–295. [Google Scholar] [CrossRef]
- Bartowsky, E. Bacterial spoilage of wine and approaches to minimize it. Lett. Appl. Microbiol. 2009, 48, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.W. The evolutionary origins of organelles. Trends Genet. 1989, 5, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Stefanini, I.; Cavalieri, D. Metagenomic Approaches to Investigate the Contribution of the Vineyard Environment to the Quality of Wine Fermentation: Potentials and Difficulties. Front. Microbiol. 2018, 9, 991. [Google Scholar] [CrossRef]
- Du Toit, W.J.; Pretorius, I.S. The occurrence, control and esoteric effect of acetic acid bacteria in winemaking. Ann. Microbiol. 2002, 52, 155e179. [Google Scholar]
- Gonzalez, A.; Hierro, N.; Poblet, M.; Mas, A.; Guillamon, J.M. Application of molecular methods to demonstrate species and strain evolution of acetic acid bacteria population during wine production. Int. J. Food Microbiol. 2005, 102, 295e304. [Google Scholar] [CrossRef]
- Lonvaud-Funel, A. Lactic acid bacteria in the quality improvement and depreciation of wine. In Lactic Acid Bacteria: Genetics, Metabolism and Applications ; Springer: Dordrecht, Germany, 1999; Volume 76, pp. 317–331. [Google Scholar] [CrossRef]
- Lu, Y.; Sun, F.; Wang, W.; Liu, Y.; Wang, J.; Sun, J.; Mu, J.; Gao, Z. Effects of spontaneous fermentation on the microorganism diversity and volatile compounds during ‘Marselan’ from grape to wine. LWT 2020, 134, 110193. [Google Scholar] [CrossRef]
- Pinto, C.; Pinho, D.; Cardoso, R.; Custódio, V.; Fernandes, J.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Wine fermentation microbiome: A landscape from different Portuguese wine appellations. Front. Microbiol. 2015, 6, 905. [Google Scholar] [CrossRef] [Green Version]
- Sternes, P.R.; Lee, D.; Kutyna, D.R.; Borneman, A.R. A combined meta-barcoding and shotgun metagenomic analysis of spontaneous wine fermentation. GigaScience 2017, 6, gix040. [Google Scholar] [CrossRef] [Green Version]
- Belda, I.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Improvement of aromatic thiol release through the selection of yeasts with increased β-lyase activity. Int. J. Food Microbiol. 2016, 225, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Merín, M.G.; Morata de Ambrosini, V.I. Highly cold-active pectinases under wine-like conditions from non-Saccharomyces yeasts for enzymatic production during winemaking. Lett. Appl. Microbiol. 2015, 60, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Merín, M.G.; Mendoza, L.M.; Farías, M.E.; Morata de Ambrosini, V.I. Isolation and selection of yeasts from wine grape ecosystem secreting cold-active. Int. J. Food Microbiol. 2011, 147, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Jimenez, J.M.; Mingorance-Cazorla, L.; Martınez-Rodrıguez, S.; Las Heras-Vazquez, F.J.; Rodrıguez-Vico, F. Molecular characterization and oenological properties of wine yeasts isolated during spontaneous fermentation of six varieties of grape must. Food Microbiol. 2004, 21, 149–155. [Google Scholar] [CrossRef]
- Andorrà, I.; Landi, S.; Mas, A.; Esteve-Zarzoso, B.; Guillamon, J.M. Effect of fermentation temperature on microbial population evolution using culture independent and dependent techniques. Int. Food Res. J. 2010, 43, 773–779. [Google Scholar] [CrossRef]
- Benito, S.; Palomero, F.; Calderón, F.; Palmero, D.; Suárez-Lepe, J.A. Selection of appropriate Schizosaccharomyces strains for winemaking. Food Microbiol. 2014, 42, 218–224. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
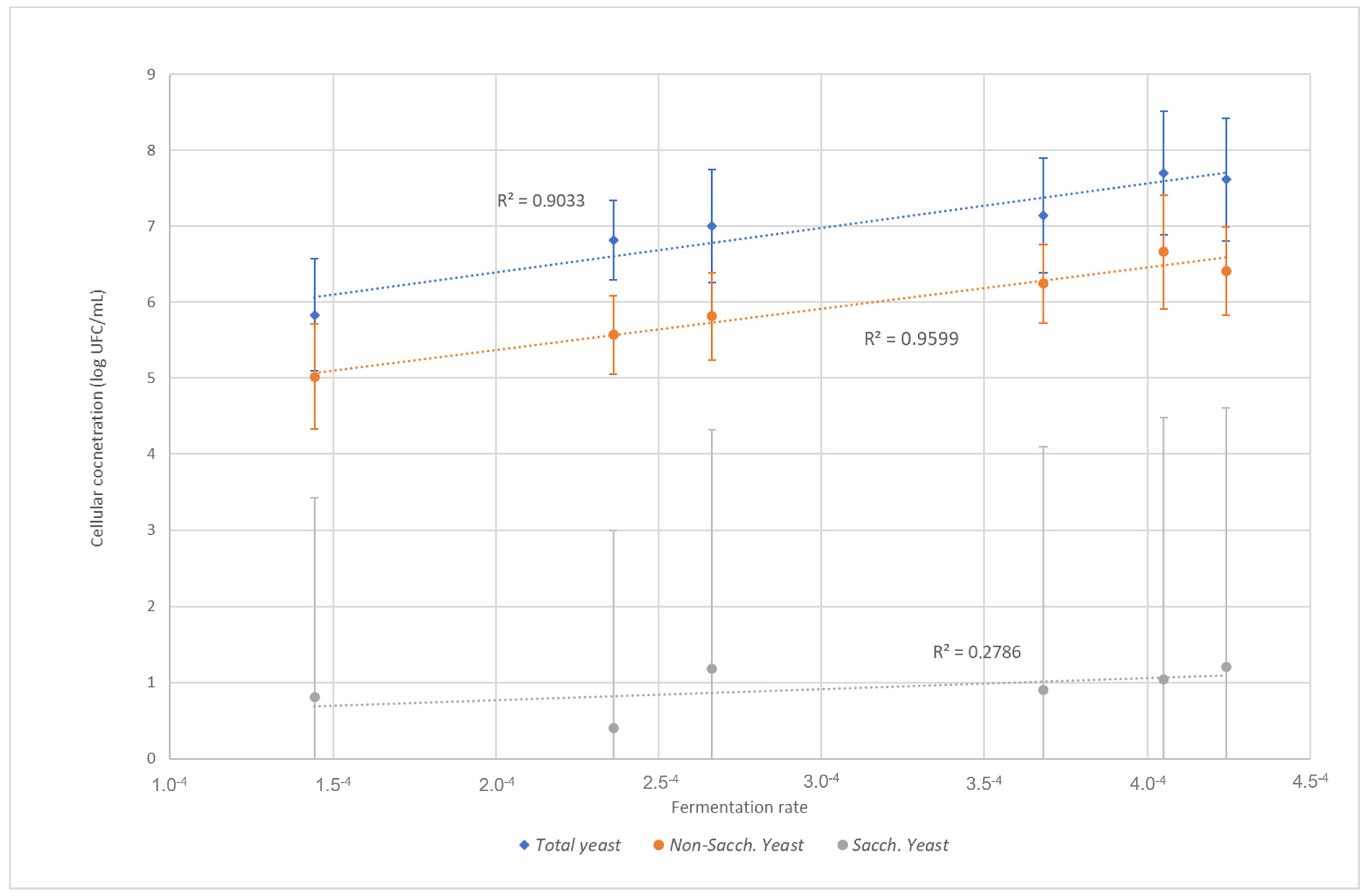{kind=link}
{kind=link}
| Grape Must | ||||||
| Grape variety | Total Sugars g/L | Total Acidity g/L | Malic acid g/L | pH | YAN mg/L | |
| Trebbiano | 232 ± 7 | 5.9 ± 0.7 | 2.1 ± 0.3 | 3.5 ± 0.1 | 54 ± 1 | |
| Montepulciano | 232 ± 9 | 5.7 ± 0.6 | 1.7 ± 0.6 | 3.5 ± 0.1 | 56 ± 4 | |
| Wine | ||||||
| Grape variety | Total Sugars g/L | Total Acidity g/L | Malic acid g/L | pH | Acetic acid g/L | Ethanol (%) |
| Trebbiano | 1.8 ± 0.8 | 4.8 ± 0.7 | N.d. | 3.8 ± 0.1 | 0.4 ± 0.1 | 13.8 ± 0.3 |
| Montepulciano | 2.4 ± 1.1 | 4.8 ± 0.5 | N.d. | 3.8 ± 0.2 | 0.6 ± 0.1 | 13.7 ± 0.6 |
| Must | TY | NSY | LAB |
|---|---|---|---|
| (Log CFU/g) | |||
| Trebbiano | |||
| Day 1 | 5.6 ± 0.6 a | 5.0 ± 0.4 a | N.d. |
| Day 3 | 7.4 ± 0.5 b | 6.5 ± 0.2 ab | N.d. |
| Day 6 | 7.7 ± 0.6 b | 6.4 ± 0.6 ab | 2.5 ± 0.3 a |
| Day 9 | 7.4 ± 0.8 b | 6.3 ± 0.4 ab | 5.8 ± 0.2 b |
| Day 15 | 7.5 ± 0.5 b | 5.7 ± 0.2 a | 6.5 ± 0.6 b |
| Day 30 | 4.9 ± 0.5 a | 4.9 ± 0.6 a | 4.6 ± 0.4 c |
| Montepulciano | |||
| Day 1 | 7.0 ± 0.8 ab | 7.1 ± 1.0 a | 5.7 ± 0.1 b |
| Day 3 | 7.9 ± 0.2 b | 6.8 ± 1.5 a | 6.9 ± 0.5 c |
| Day 6 | 7.0 ± 0.5 ab | 6.2 ± 2.0 a | 5.8 ± 0.2 b |
| Day 9 | 6.6 ± 1.0 a | 6.6 ± 1.0 a | 4.5 ± 1.0 a |
| Day 15 | 7.3 ± 0.4 ab | 6.3 ± 1.1 a | 5.9 ± 0.5 b |
| Day 30 | 7.6 ± 0.3 b | 6.9 ± 1.3 a | 6.8 ± 0.5 c |
| Obs. OTUs | Shannon div. ind. | |
|---|---|---|
| Bacteria | ||
| Wine | ||
| Trebbiano | 88 ± 50 | 5.6 ± 0.6 |
| Montepulciano | 85 ± 26 | 5.7 ± 0.6 |
| Stage of Fermentation | ||
| Day 1 | 93 ± 38 | 5.8 ± 0.6 |
| Day 3 | 107 ± 63 | 5.9 ± 0.4 |
| Day 6 | 81 ± 80 | 5.6 ± 0.2 |
| Day 9 | 87 ± 90 | 5.6 ± 0.3 |
| Day 15 | 69 ± 33 | 5.3 ± 0.9 |
| Day 30 | 78 ± 31 | 5.6 ± 0.7 |
| Fungii | ||
| Wine | ||
| Trebbiano | 23 ± 11 | 2.7 ± 1.2 |
| Montepulciano | 27 ± 12 | 3.0 ± 0.9 |
| Stage of Fermentation | ||
| Day 1 | 39 ± 12 a | 4.0 ± 0.2 a |
| Day 3 | 29 ± 80 a | 3.5 ± 0.4 a |
| Day 6 | 27 ± 8 a | 2.9 ± 0.6 a |
| Day 9 | 21 ± 4 ab | 2.6 ± 0.7 ab |
| Day 15 | 15 ± 4 b | 2.0 ± 0.9 b |
| Day 30 | 12 ± 10 b | 1.8 ± 0.8 b |
| Main Effects | Pseudo-F | p-Value |
|---|---|---|
| Bacteria | ||
| Wine | 2.815 | 0.086 |
| Fermentation day | 6.374 | 0.001 ** |
| Pairwise Comparisons for Fermentation Day | Pseudo-F | p-Value |
| D1 vs. D3 | 1.182 | 0.325 |
| D1 vs. D6 | 8.295 | 0.003 ** |
| D1 vs. D9 | 4.756 | 0.004 ** |
| D1 vs. D15 | 6.684 | 0.003 ** |
| D1 vs. D30 | 16.570 | 0.004 ** |
| D3 vs. D6 | 17.302 | 0.001 ** |
| D3 vs. D9 | 7.030 | 0.003 ** |
| D3 vs. D15 | 9.373 | 0.002 ** |
| D3 vs. D30 | 23.741 | 0.003 ** |
| D6 vs. D9 | 2.999 | 0.061 |
| D6 vs. D15 | 5.523 | 0.020 * |
| D6 vs. D30 | 17.341 | 0.007 ** |
| D9 vs. D15 | 0.311 | 0.556 |
| D9 vs. D30 | 2.207 | 0.181 |
| D15 vs. D30 | 0.742 | 0.423 |
| Fungii | ||
| Wine | 12.875 | 0.001 ** |
| Fermentation day | 1.659 | 0.072 |
| Pairwise Comparisons for Fermentation Day | Pseudo-F | p-Value |
| D1 vs. D3 | 1.085 | 0.360 |
| D1 vs. D6 | 3.195 | 0.010 ** |
| D1 vs. D9 | 4.865 | 0.004 ** |
| D1 vs. D15 | 3.423 | 0.015 * |
| D1 vs. D30 | 3.360 | 0.015 * |
| D3 vs. D6 | 1.222 | 0.371 |
| D3 vs. D9 | 2.247 | 0.080 |
| D3 vs. D15 | 1.800 | 0.112 |
| D3 vs. D30 | 1.892 | 0.082 |
| D6 vs. D9 | 0.340 | 0.659 |
| D6 vs. D15 | 0.437 | 0.527 |
| D6 vs. D30 | 0.674 | 0.489 |
| D9 vs. D15 | 0.334 | 0.717 |
| D9 vs. D30 | 0.425 | 0.728 |
| D15 vs. D30 | 0.712 | 0.251 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guzzon, R.; Franciosi, E.; Toffanin, A. Investigation by High-Throughput Sequencing Methods of Microbiota Dynamics in Spontaneous Fermentation of Abruzzo (South Italy) Wines. Agronomy 2022, 12, 3104. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12123104
Guzzon R, Franciosi E, Toffanin A. Investigation by High-Throughput Sequencing Methods of Microbiota Dynamics in Spontaneous Fermentation of Abruzzo (South Italy) Wines. Agronomy. 2022; 12(12):3104. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12123104
Chicago/Turabian StyleGuzzon, Raffaele, Elena Franciosi, and Annita Toffanin. 2022. "Investigation by High-Throughput Sequencing Methods of Microbiota Dynamics in Spontaneous Fermentation of Abruzzo (South Italy) Wines" Agronomy 12, no. 12: 3104. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12123104