
Polyphasic Characterization of Acidovorax citrulli Strains Originating from Serbia

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation of Bacterial Strains
2.2. Phenotypic Characterization
2.3. Genomic DNA Extraction and PCR Identification
2.4. 16S rRNA Sequence Analysis
2.5. Pathogenicity Test
2.6. Host Preference
2.7. Watermelon Cultivar’s Susceptibility to A. citrulli
2.8. Strains Sensitivity to Bactericides
2.9. Differentiation of Strains by Rep-PCR
2.10. PCR Assays for Distinguishing Group I and II A. citrulli Strains
2.11. Multilocus Sequence Analysis (MLSA)
3. Results
3.1. Isolation and Phenotypic Characterization of Bacterial Strains
3.2. PCR Identification and 16S rRNA Sequence Analysis
3.3. Pathogenicity Test and Host Preference
3.4. Cultivars Susceptibility to A. citrulli
3.5. Susceptibility to Bactericides
3.6. Differentiation of Strains by Rep-PCR
3.7. Distinguishing Group I and II of A. citrulli Strains
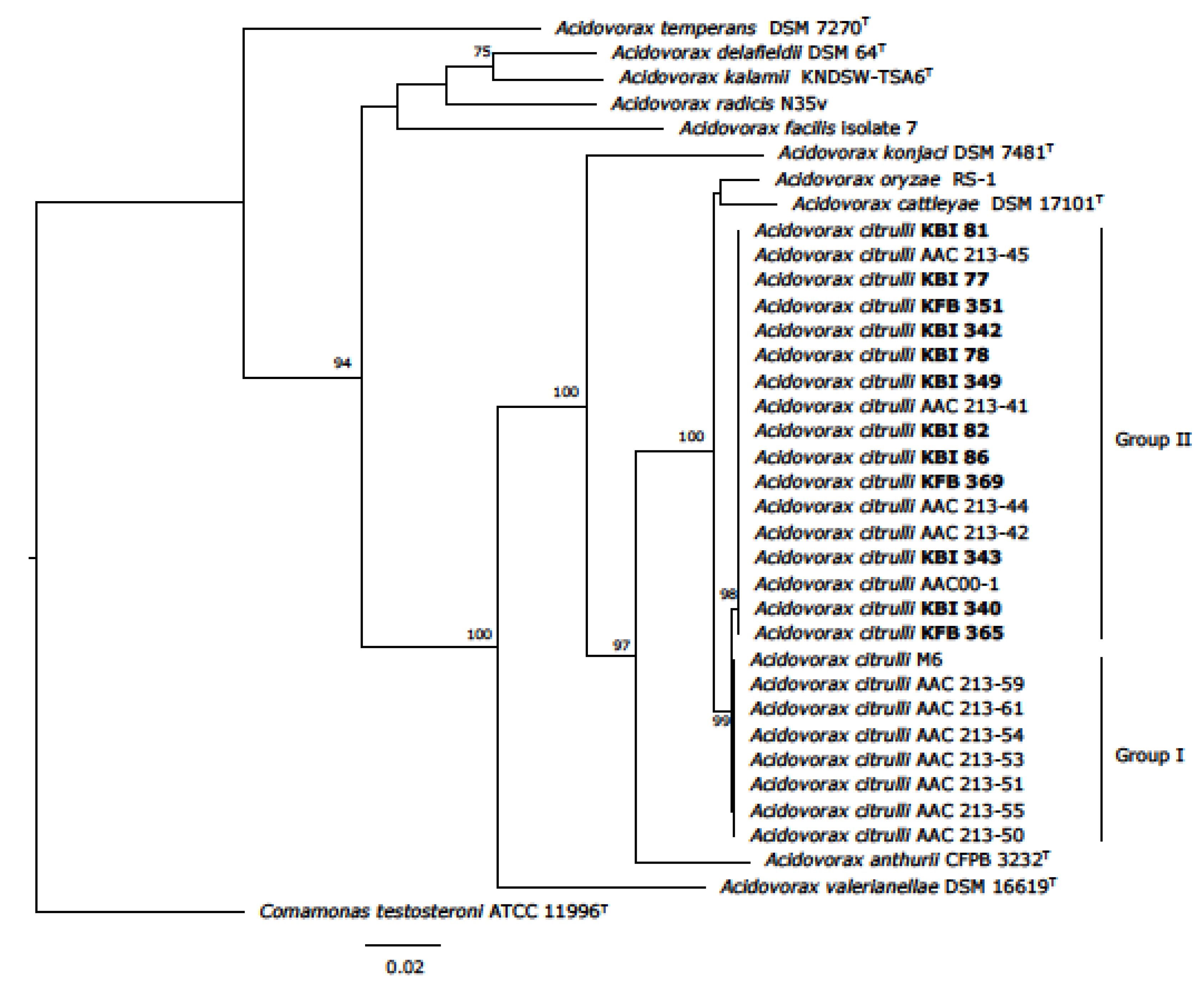
3.8. Multilocus Sequence Analysis (MLSA)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization of the United Nations. Available online: www.fao.ogr/faostat (accessed on 25 September 2021).
- Webb, R.; Goth, R. A seedborne bacterium isolated from watermelon. Plant Dis. Rep. 1965, 49, 818–821. [Google Scholar]
- Schaad, N.W.; Sowell, G.; Goth, R.W.; Colwell, R.R.; Webb, R.E. Pseudomonas pseudoalcaligenes subsp. citrulli subsp. nov. Int. J. Syst. Evol. Microbiol. 1978, 28, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.J.; Li, J.Q.; Walcott, R.R.; Zhang, G.M.; Luo, L.X.; Kang, L.; Zheng, Y.; Schaad, N.W. Advances in detection of Acidovorax citrulli, the causal agent of bacterial fruit blotch of cucurbits. Seed Sci. Technol. 2013, 41, 1–15. [Google Scholar] [CrossRef]
- Schaad, N.W.; Postnikova, E.; Sechler, A.; Claflin, L.E.; Vidaver, A.K.; Jones, J.B.; Agarkova, I.; Ignatov, A.; Dickstein, E.; Ramundo, B.A. Reclassification of subspecies of Acidovorax avenae as A. avenae (Manns 1905) emend., A. cattleyae (Pavarino 1911) comb. nov., A. citrulli (Schaad et al., 1978) comb. nov., and proposal of A. oryzae sp. nov. Syst. Appl. Microbiol. 2008, 31, 434–446. [Google Scholar] [CrossRef] [Green Version]
- Bahar, O.; Burdman, S. Bacterial fruit blotch: A threat to the cucurbit industry. Isr. J. Plant Sci. 2010, 58, 19–31. [Google Scholar] [CrossRef]
- Burdman, S.; Walcott, R. Acidovorax citrulli: Generating basic and applied knowledge to tackle a global threat to the cucurbit industry. Mol. Plant Pathol. 2012, 13, 805–815. [Google Scholar] [CrossRef]
- Feng, J.; Schuenzel, E.L.; Li, J.; Schaad, N.W. Multilocus sequence typing reveals two evolutionary lineages of Acidovorax avenae subsp. citrulli. Phytopathology 2009, 99, 913–920. [Google Scholar] [CrossRef] [Green Version]
- Mirik, M.; Aysan, Y.; Sahin, Y. Occurrence of bacterial fruit blotch of watermelon caused by Acidvorax avenae subsp. citrulli in the eastern Mediterranean region of Turkey. Plant Dis. 2006, 90, 829. [Google Scholar] [CrossRef]
- Palkovics, L.; Petróczy, M.; Kertész, B.; Németh, J.; Bársony, C.; Mike, Z.; Hevesi, M. First Report of Bacterial Fruit Blotch of Watermelon Caused by Acidovorax avenae subsp. citrulli in Hungary. Plant Dis. 2008, 92, 834. [Google Scholar] [CrossRef] [Green Version]
- Holeva, M.C.; Karafla, C.D.; Glynos, P.E.; Alivizatos, A.S. Acidovorax avenae subsp. citrulli newly reported to cause bacterial fruit blotch of watermelon in Greece. Plant Pathol. 2010, 59, 797. [Google Scholar] [CrossRef]
- Walcott, R.R.; Fessehaie, A.; Castro, A.C. Differences in Pathogenicity between two Genetically Distinct Groups of Acidovorax avenae subsp. citrulli on Cucurbit Hosts. J. Phytopathol. 2004, 152, 277–285. [Google Scholar] [CrossRef]
- Zhao, M.; Walcott, R. Acidovorax citrulli: History, Epidemiology, and Management of Bacterial Fruit Blotch of Cucurbits. In Plant Pathogenic Acidovorax Species; Burdman, S., Walcott, R., Eds.; APS Press: Saint Paul, MN, USA, 2018; pp. 39–57. [Google Scholar] [CrossRef]
- Assouline, I.; Milshtein, H.; Mizrachi, M.; Levy, E.; Ben-Ze’ev, I.S. Acidovorax avenae subsp. citrulli transmitted by Solanaceous seeds. Phytoparasitica 1997, 25, 2. [Google Scholar] [CrossRef]
- Burdman, S.; Kots, N.; Kritzman, G.; Kopelowitz, J. Molecular, physiological, and host-range characterization of Acidovorax avenae subsp. citrulli isolates from watermelon and melon in Israel. Plant Dis. 2005, 89, 1339–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalupowicz, L.; Reuven, M.; Dror, O.; Sela, N.; Burdman, S.; Manulis-Sasson, S. Characterization of Acidovorax citrulli strains isolated from solanaceous plants. Plant Pathol. 2020, 69, 1787–1797. [Google Scholar] [CrossRef]
- Malliarakis, D.; Mpalantinaki, E.; Pagoulatou, M.G.; Lorenzou, K.; Goumas, D. First report of Acidovorax citrulli causing a leaf spot disease on tomato plants in Greece. J. Plant Pathol. 2021, 103, 353. [Google Scholar] [CrossRef]
- Popović, T.; Ivanović, Ž. Occurrence of Acidovorax citrulli Causing Bacterial Fruit Blotch of Watermelon in Serbia. Plant Dis. 2015, 99, 886. [Google Scholar] [CrossRef]
- Zivanovic, M.; Walcott, R. Further characterization of genetically distinct groups of Acidovorax citrulli strains. Phytopathology 2017, 107, 29–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walcott, R.R.; Langston, D.B.; Sanders, F.H.; Gitaitis, R.D. Investigating Intraspecific Variation of Acidovorax avenae subsp. citrulli Using DNA Fingerprinting and Whole Cell Fatty Acid Analysis. Phytopathology 2000, 90, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Dutta, B.; Luo, X.; Burdman, S.; Walcott, R. Genetically Distinct Acidovorax citrulli Strains Display Cucurbit Fruit Preference under Field Conditions. Phytopathology 2020, 110, 973–980. [Google Scholar] [CrossRef]
- O’Brien, R.G.; Martin, H.L. Bacterial blotch of melons caused by strains of Acidovorax avenae subsp. citrulli. Aust. J. Exp. Agric. 1999, 39, 479–485. [Google Scholar] [CrossRef]
- Glaeser, S.P.; Kämpfer, P. Multilocus sequence analysis (MLSA) in prokaryotic taxonomy. Syst. Appl. Microbiol. 2015, 38, 237–245. [Google Scholar] [CrossRef]
- Silva, G.M.; Souza, R.M.; Yan, L.; Junior, R.S.; Medeiros, F.H.; Walcott, R.R. Strains of the group I lineage of Acidovorax citrulli, the causal agent of bacterial fruit blotch of cucurbitaceous crops, are predominant in Brazil. Phytopathology 2016, 106, 1486–1494. [Google Scholar] [CrossRef] [Green Version]
- King, E.O.; Ward, M.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescein. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar]
- Lelliott, R.A.; Stead, D.E. Methods for the diagnosis of bacterial diseases of plants. In Methods in Plant Pathology; Blackwell Scientific Publications: Oxford, UK, 1987; Volume 2. [Google Scholar]
- Jones, J.B.; Gitaitis, R.D.; Schaad, N.W. Gram-negative bacteria: Acidovorax and Xylophilus. In Laboratory Guide for Identification of Plant Pathogenic Bacteria, 3rd ed.; Schaad, N.W., Jones, J.B., Chen, W., Eds.; APS Press: Saint Paul, MN, USA, 2001; pp. 121–138. [Google Scholar]
- Lelliott, R.A.; Billing, E.; Hayward, A.C. A determinative scheme for the fluorescent plant pathogenic pseudomonads. J. Appl. Microbiol. 1966, 29, 470–489. [Google Scholar] [CrossRef] [PubMed]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef] [PubMed]
- Bahar, O.; Efrat, M.; Hadar, E.; Dutta, B.; Walcott, R.R.; Burdman, S. New subspecies-specific polymerase chain reaction-based assay for the detection of Acidovorax avenae subsp. citrulli. Plant Pathol. 2008, 57, 754–763. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Versalovic, J.; Schneider, M.; De Bruijn, F.J.; Lupski, J.R. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction. Methods Mol. Cell Biol. 1994, 5, 25–40. [Google Scholar]
- Higgins, D.G.; Thompson, J.D.; Gibson, T.J. Using CLUSTAL for multiple sequence alignments. Methods Enzymol. 1996, 266, 383–402. [Google Scholar] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.T.; Von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.F.K.; Von Haeseler, A.; Jermiin, L.S. Model Finder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Rambaut, A. Figtree v1.4.4. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 10 September 2021).
- Zlatković, N.; Prokić, A.; Kuzmanović, N.; Gašić, K.; Šević, M.; Ivanović, M.; Obradović, A. Bakteriozna mrljavost plodova lubenice u Srbiji. Biljni Lekar 2015, 43, 265–272. (In English) [Google Scholar]
- Eckshtain-Levi, N.; Munitz, T.; Živanović, M.; Traore, S.M.; Spröer, C.; Zhao, B.; Welbaum, G.; Walcott, R.; Sikorski, J.; Burdman, S. Comparative analysis of type III secreted effector genes reflects divergence of Acidovorax citrulli strains into three distinct lineages. Phytopathology 2014, 104, 1152–1162. [Google Scholar] [CrossRef] [Green Version]
- Schaad, N.W.; Jones, J.B.; Lacy, G.H. Laboratory Guide for Identification of Plant Pathogenic Bacteria, 3rd ed.; Schaad, N.W., Jones, J.B., Chun, W., Eds.; APS Press: Saint Paul, MN, USA, 2001; p. 373. [Google Scholar]
- Johnson, K.L.; Minsavage, G.V.; Le, T.; Jones, J.B.; Walcott, R.R. Efficacy of a Nonpathogenic Acidovorax citrulli Strain as a Biocontrol Seed Treatment for Bacterial Fruit Blotch of Cucurbits. Plant Dis. 2011, 95, 697–704. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, F.C.Q.; Santos, L.A.; Dias, R.C.S.; Mariano, R.L.R.; Souza, E.B. Selection of watermelon genotypes for resistance to bacterial fruit blotch. Euphytica 2013, 190, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Wechter, W.P.; Levi, A.; Ling, K.S.; Kousik, C.; Block, C.C. Identification of resistance to Acidovorax avenae subsp. citrulli among melon (Cucumis spp.) plant introductions. HortScience 2011, 46, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.R.; Hossain, M.R.; Jesse, D.M.I.; Jung, H.J.; Kim, H.T.; Park, J.I.; Nou, I.S. Development of Molecular Marker Linked with Bacterial Fruit Blotch Resistance in Melon (Cucumis melo L.). Genes 2020, 11, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šević, M.; Gašić, K.; Ignjatov, M.; Mijatović, M.; Prokić, A.; Obradović, A. Integration of biological and conventional treatments in control of pepper bacterial spot. Crop Prot. 2019, 119, 46–51. [Google Scholar] [CrossRef]
- Kan, Y.; Jiang, N.; Xu, X.; Lyu, Q.; Gopalakrishnan, V.; Walcott, R.; Burdman, S.; Li, J.; Luo, L. Induction and Resuscitation of the Viable but Non-culturable (VBNC) State in Acidovorax citrulli, the Causal Agent of Bacterial Fruit Blotch of Cucurbitaceous Crops. Front. Microbiol. 2019, 10, 1081. [Google Scholar] [CrossRef] [PubMed]
- Walcott, R.; Gitaitis, R.D.; Castro, A.C. Role of blossoms in watermelon seed Infestation by Acidovorax avenae subsp. citrulli. Phytopathology 2003, 93, 528–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Strain Data | ||||
|---|---|---|---|---|
| Strain | Host (Variety) | Isolation Source | Locality | Year |
| KFB a 340 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KFB 341 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KFB 342 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KFB 343 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KFB 344 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KFB 345 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KFB 346 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KFB 347 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KFB 348 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KFB 349 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KFB 350 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KFB 351 | C. lanatus, (Unknow) | Fruit | Čelarevo | 2014 |
| KFB 352 | C. lanatus, (Unknow) | Fruit | Čelarevo | 2014 |
| KFB 365 | C. lanatus, (Top Gun) | Fruit | Rečka | 2015 |
| KFB 366 | C. lanatus, (Top Gun) | Fruit | Rečka | 2015 |
| KFB 367 | C. lanatus, (Top Gun) | Fruit | Rečka | 2015 |
| KFB 368 | C. lanatus, (Top Gun) | Fruit | Rečka | 2015 |
| KFB 369 | C. lanatus, (Eleta) | Fruit | Šabac | 2016 |
| KFB 370 | C. lanatus, (Eleta) | Fruit | Šabac | 2016 |
| KFB 371 | C. lanatus, (Eleta) | Fruit | Šabac | 2016 |
| KFB 372 | C. lanatus, (Eleta) | Fruit | Šabac | 2016 |
| KBI b 76 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KBI 77 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KBI 78 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KBI 79 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KBI 80 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KBI 81 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KBI 82 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KBI 83 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KBI 84 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KBI 85 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KBI 86 | C. lanatus, (Farao) | Fruit | Ašanja | 2014 |
| KBI 340 | C. lanatus, (Unknow) | Fruit | Leskovac | 2018 |
| KBI 341 | C. lanatus, (Unknow) | Fruit | Leskovac | 2018 |
| KBI 342 | C. lanatus, (Unknow) | Leaf | Leskovac | 2018 |
| KBI 343 | C. lanatus, (Unknow) | Fruit | Šabac | 2018 |
| KBI 344 | C. lanatus, (Unknow) | Fruit | Šabac | 2018 |
| KBI 345 | C. lanatus, (Unknow) | Fruit | Šabac | 2018 |
| KBI 346 | C. lanatus, (Unknow) | Fruit | Šabac | 2018 |
| KBI 347 | C. lanatus, (Unknow) | Fruit | Šabac | 2018 |
| KBI 348 | C. lanatus, (Unknow) | Fruit | Šabac | 2018 |
| KBI 349 | C. lanatus, (Unknow) | Fruit | Šabac | 2018 |
| KBI 350 | C. lanatus, (Unknow) | Fruit | Šabac | 2018 |
| Test | Results a | ||
|---|---|---|---|
| Investigated Strains b | Control Strain c | ||
| KFB 340–352, KFB 365–372, KBI 76–81, KBI 83–85, KBI 340–350 | KBI 82, KBI 86 | NCPPB 3679T | |
| Gram reaction | − | − | − |
| Catalase activity | + | + | + |
| Growth on 41 °C | + | + | + |
| Fluorescens on Kings medium B | − | − | − |
| Levan production | − | − | − |
| Oxidase production | + | + | + |
| Pectinolytic activity | − | − | − |
| Arginine dihydrolase production | − | − | − |
| Tobacco hypersensitivity | + | − | + |
| Oxidative–fermentative (O/F) test | (O) | (O) | (O) |
| Nitrate reduction | − | − | − |
| Acid production from: | |||
| Arabinose | + | + | + |
| Sucrose | − | − | − |
| Sorbitol | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zlatković, N.; Gašić, K.; Kuzmanović, N.; Prokić, A.; Ivanović, M.; Živković, S.; Obradović, A. Polyphasic Characterization of Acidovorax citrulli Strains Originating from Serbia. Agronomy 2022, 12, 235. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020235
Zlatković N, Gašić K, Kuzmanović N, Prokić A, Ivanović M, Živković S, Obradović A. Polyphasic Characterization of Acidovorax citrulli Strains Originating from Serbia. Agronomy. 2022; 12(2):235. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020235
Chicago/Turabian StyleZlatković, Nevena, Katarina Gašić, Nemanja Kuzmanović, Anđelka Prokić, Milan Ivanović, Svetlana Živković, and Aleksa Obradović. 2022. "Polyphasic Characterization of Acidovorax citrulli Strains Originating from Serbia" Agronomy 12, no. 2: 235. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020235