
Correlational Analysis of Agronomic and Seed Quality Traits in Camelina sativa Doubled Haploid Lines under Rain-Fed Condition

 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Conditions
2.2. Experimental Design
2.3. Morphological Traits Measurement
2.4. Oil and Protein Content and Free Fatty Acids Measurement
2.5. Statistical Analysis
3. Results
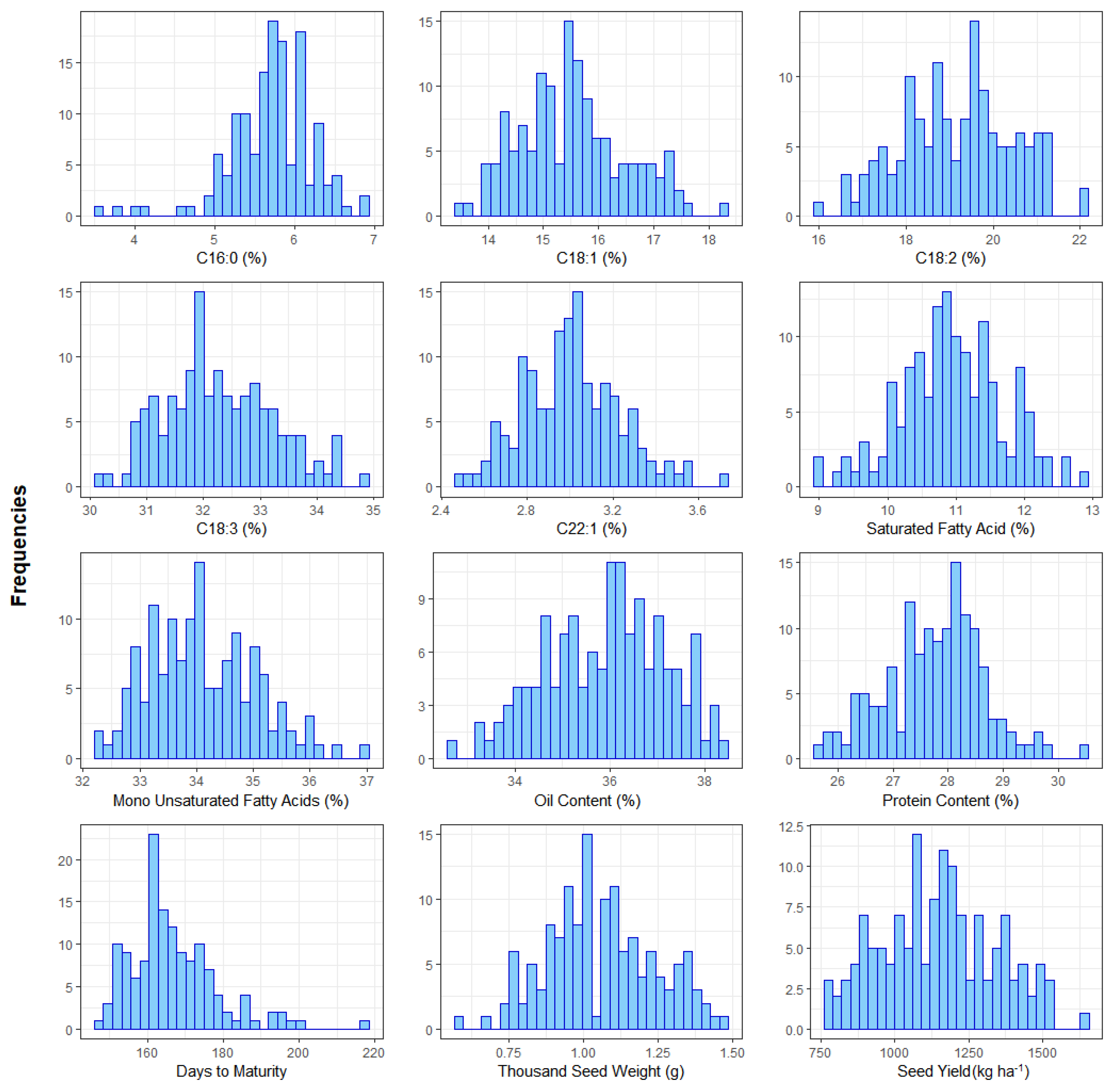
3.1. Comparison of Plant Grwoth and Fatty Acids in DH Lines of Camelina
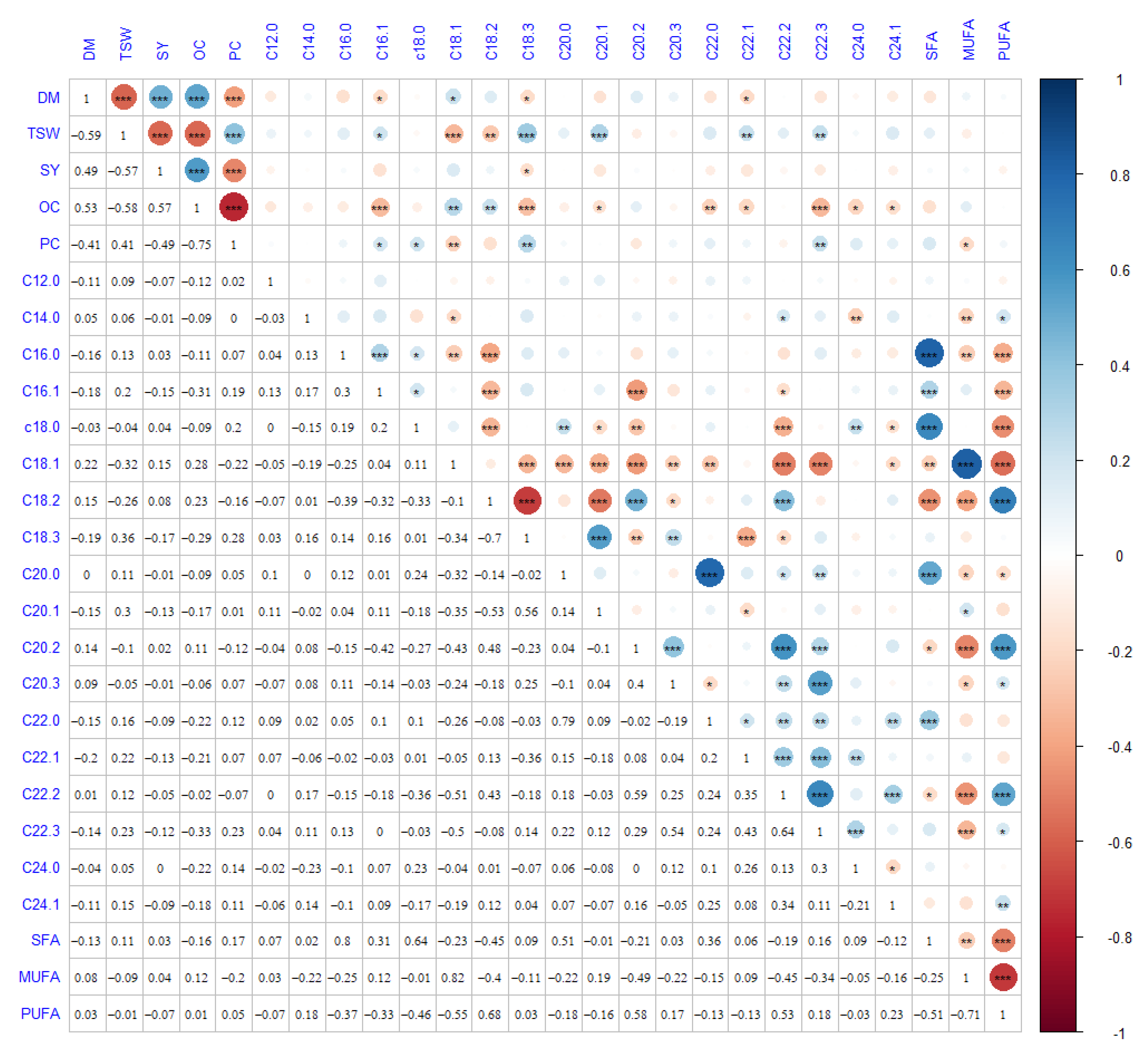
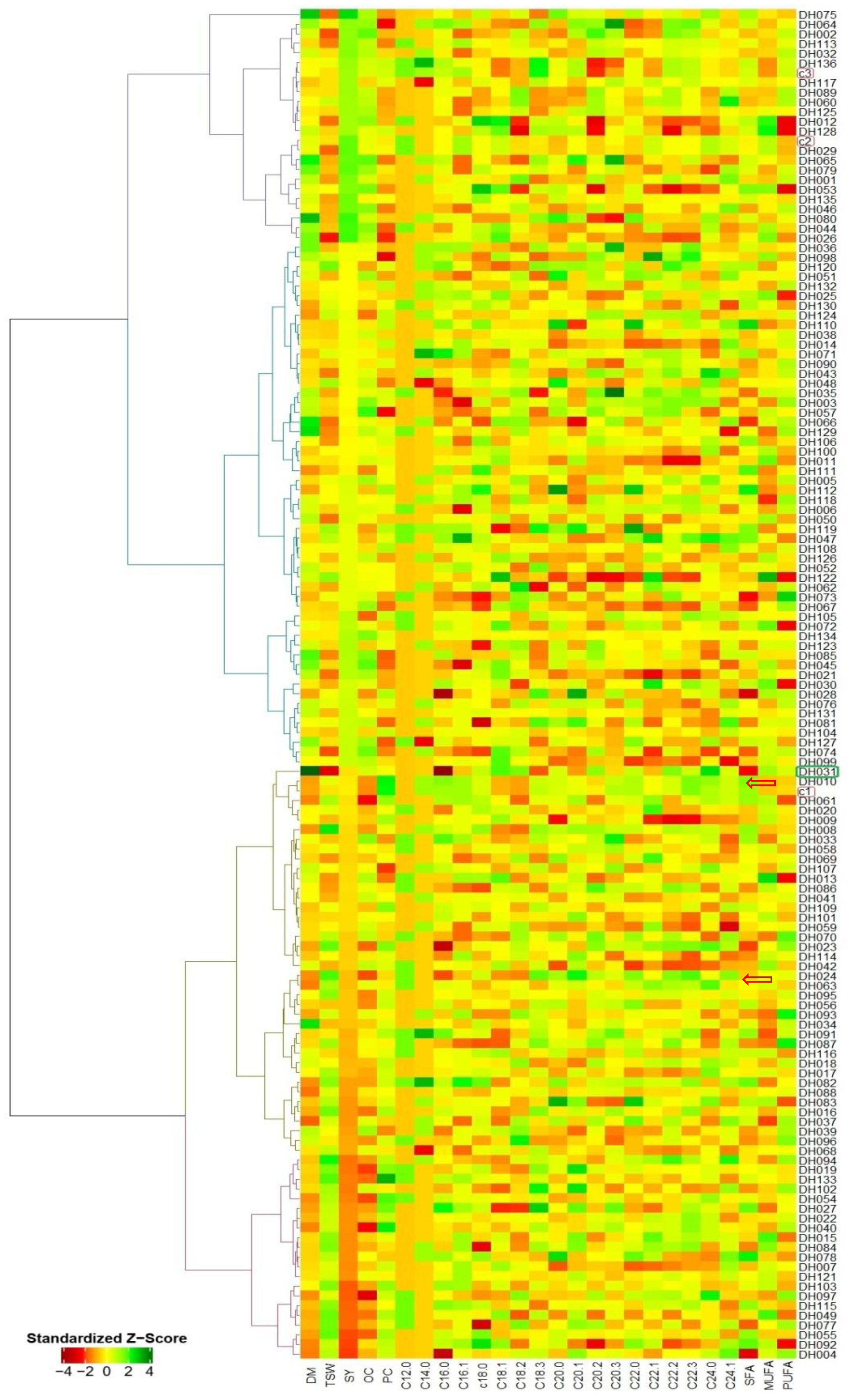
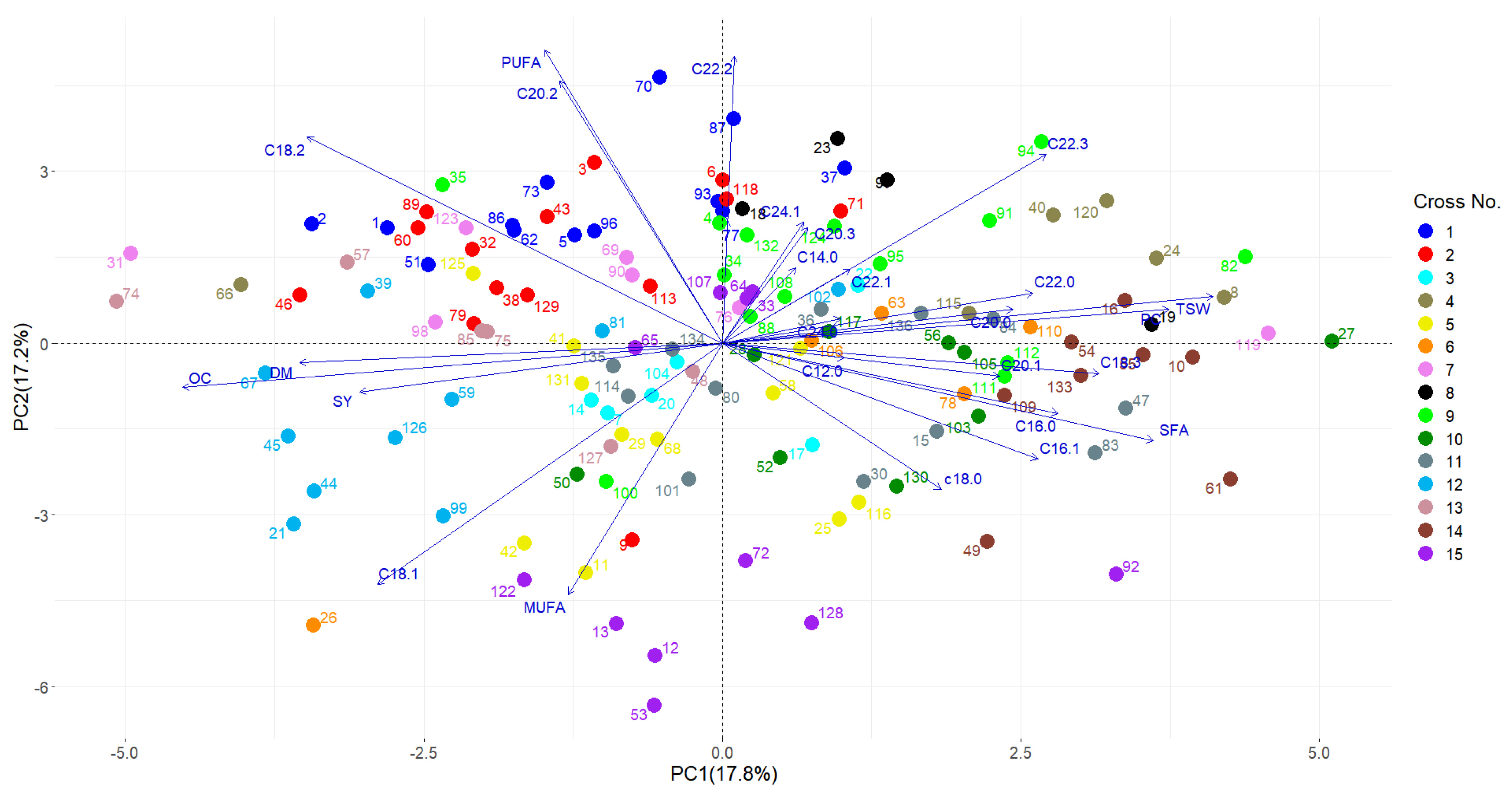
3.2. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vollmann, J.; Eynck, C. Camelina as a sustainable oilseed crop: Contributions of plant breeding and genetic engineering. Biotechnol. J. 2015, 10, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Kawall, K. Genome-edited Camelina sativa with a unique fatty acid content and its potential impact on ecosystems. Environ. Sci. Eur. 2021, 33, 38. [Google Scholar] [CrossRef]
- White, G.; Gardner, J.; Cook, C. Biodiversity for industrial crop development in the United States. Ind. Crops Prod. 1994, 2, 259–272. [Google Scholar] [CrossRef]
- Kon’Kova, N.; Shelenga, T.; Gridnev, G.; Dubovskaya, A.; Malyshev, L. Stability and Variability of Camelina sativa (L.) Crantz Economically Valuable Traits in Various Eco-Geographical Conditions of the Russian Federation. Agronomy 2021, 11, 332. [Google Scholar] [CrossRef]
- Zanetti, F.; Eynck, C.; Christou, M.; Krzyżaniak, M.; Righini, D.; Alexopoulou, E.; Stolarski, M.J.; van Loo, E.N.; Puttick, D.; Monti, A. Agronomic performance and seed quality attributes of Camelina (Camelina sativa L. crantz) in multi-environment trials across Europe and Canada. Ind. Crops Prod. 2017, 107, 602–608. [Google Scholar] [CrossRef] [Green Version]
- Leclère, M.; Loyce, C.; Jeuffroy, M.-H. Growing camelina as a second crop in France: A participatory design approach to produce actionable knowledge. Eur. J. Agron. 2018, 101, 78–89. [Google Scholar] [CrossRef]
- Horvath, D.; Anderson, J.V.; Chao, W.S.; Zheng, P.; Buchwaldt, M.; Parkin, I.A.P.; Dorn, K. Genes associated with chloroplasts and hormone-signaling, and transcription factors other than CBFs are associated with differential survival after low temperature treatments of Camelina sativa biotypes. PLoS ONE 2019, 14, e0217692. [Google Scholar] [CrossRef]
- Krzyżaniak, M.; Stolarski, M.J.; Tworkowski, J.; Puttick, D.; Eynck, C.; Załuski, D.; Kwiatkowski, J. Yield and seed composition of 10 spring camelina genotypes cultivated in the temperate climate of Central Europe. Ind. Crops Prod. 2019, 138, 111443. [Google Scholar] [CrossRef]
- Martin, S.L.; Lujan-Toro, B.E.; Sauder, C.A.; James, T.; Ohadi, S.; Hall, L.M. Hybridization rate and hybrid fitness for Camelina microcarpa Andrz. ex DC (♀) and Camelina sativa (L.) Crantz(Brassicaceae) (♂). Evol. Appl. 2018, 12, 443–455. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.-J.; Auer, C. Overwintering assessment of camelina (Camelina sativa) cultivars and congeneric species in the northeastern US. Ind. Crops Prod. 2019, 139, 111532. [Google Scholar] [CrossRef]
- Anderson, J.V.; Neubauer, M.; Horvath, D.P.; Chao, W.S.; Berti, M.T. Analysis of Camelina sativa transcriptomes identified specific transcription factors and processes associated with freezing tolerance in a winter biotype. Ind. Crops Prod. 2021, 177, 114414. [Google Scholar] [CrossRef]
- Choi, S.-H.; Park, N.; Lee, K.-Y.; Missaoui, A.M.; Lee, G.-J. Novel genes in response to varying water deficit in oil crop Camelina sativa. Euphytica 2019, 215, 86. [Google Scholar] [CrossRef]
- Berti, M.; Gesch, R.; Eynck, C.; Anderson, J.; Cermak, S. Camelina uses, genetics, genomics, production, and management. Ind. Crops Prod. 2016, 94, 690–710. [Google Scholar] [CrossRef]
- Walia, M.K.; Wells, M.S.; Cubins, J.; Wyse, D.; Gardner, R.D.; Forcella, F.; Gesch, R. Winter camelina seed yield and quality responses to harvest time. Ind. Crops Prod. 2018, 124, 765–775. [Google Scholar] [CrossRef]
- Brock, J.R.; Dönmez, A.A.; Beilstein, M.A.; Olsen, K.M. Phylogenetics of Camelina Crantz. (Brassicaceae) and insights on the origin of gold-of-pleasure (Camelina sativa). Mol. Phylogenetics Evol. 2018, 127, 834–842. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.D.; Anderson, J.L. Ruminal degradation and intestinal digestibility of Camelina meal and carinata meal compared with other protein sources. Prof. Anim. Sci. 2018, 34, 10–18. [Google Scholar] [CrossRef]
- Martinez, S.; Alvarez, S.; Capuano, A.; Delgado, M.D.M. Environmental performance of animal feed production from Camelina sativa (L.) Crantz: Influence of crop management practices under Mediterranean conditions. Agric. Syst. 2020, 177, 102717. [Google Scholar] [CrossRef]
- Fröhlich, A.; Rice, B. Evaluation of Camelina sativa oil as a feedstock for biodiesel production. Ind. Crops Prod. 2005, 21, 25–31. [Google Scholar] [CrossRef]
- Zanetti, F.; Alberghini, B.; Jeromela, A.M.; Grahovac, N.; Rajković, D.; Kiprovski, B.; Monti, A. Camelina, an ancient oilseed crop actively contributing to the rural renaissance in Europe. A review. Agron. Sustain. Dev. 2021, 41, 2. [Google Scholar] [CrossRef]
- Eynck, C.; Falk, K.C. Camelina (Camelina sativa). In Biofuel Crops: Production, Physiology and Genetics; Singh, B., Ed.; CABI Publishing: Wallingford, UK, 2013; Volume 1845938852, pp. 369–391. [Google Scholar]
- Francis, A.; Warwick, S.I. The biology of Canadian weeds. 142. Camelina alyssum (Mill.) Thell.; C. microcarpa Andrz. ex DC.; C. sativa (L.) Crantz. Can. J. Plant Sci. 2009, 89, 791–810. [Google Scholar] [CrossRef]
- Righini, D.; Zanetti, F.; Martínez-Force, E.; Mandrioli, M.; Toschi, T.G.; Monti, A. Shifting sowing of camelina from spring to autumn enhances the oil quality for bio-based applications in response to temperature and seed carbon stock. Ind. Crops Prod. 2019, 137, 66–73. [Google Scholar] [CrossRef]
- Soorni, J.; Kazemitabar, S.K.; Kahrizi, D.; Dehestani, A.; Bagheri, N. Genetic analysis of freezing tolerance in camelina [Camelina sativa (L.) Crantz] by diallel cross of winter and spring biotypes. Planta 2021, 253, 9. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Liu, X.; Yiu, S.-M.; Lim, B.L. De novo assembly and characterization of Camelina sativa transcriptome by paired-end sequencing. BMC Genom. 2013, 14, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagale, S.; Koh, C.; Nixon, J.; Bollina, V.; Clarke, W.E.; Tuteja, R.; Spillane, C.; Robinson, S.J.; Links, M.; Clarke, C.; et al. The emerging biofuel crop Camelina sativa retains a highly undifferentiated hexaploid genome structure. Nat. Commun. 2014, 5, 3706. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.R.; Tang, J.; Sharma, N.; Burkitt, C.; Ji, Y.; Mykytyshyn, M.; Bohmert-Tatarev, K.; Peoples, O.; Snell, K.D. Camelina sativa, an oilseed at the nexus between model system and commercial crop. Plant Cell Rep. 2018, 37, 1367–1381. [Google Scholar] [CrossRef] [PubMed]
- Mandáková, T.; Pouch, M.; Brock, J.R.; Al-Shehbaz, I.A.; Lysak, M.A. Origin and Evolution of Diploid and Allopolyploid Camelina Genomes was Accompanied by Chromosome Shattering. Plant Cell 2019, 31, 2596–2612. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Silva, J.E.; Podicheti, R.; Macrander, J.; Yang, W.; Nazarenus, T.J.; Nam, J.-W.; Jaworski, J.G.; Lu, C.; Scheffler, B.E.; et al. Camelina seed transcriptome: A tool for meal and oil improvement and translational research. Plant Biotechnol. J. 2013, 11, 759–769. [Google Scholar] [CrossRef] [Green Version]
- Shahadati-Moghaddam, Z.; Hosseini, A.; Soorni, J.; Trojak-Goluch, A. Development of tobacco (Nicotiana tabacum L.) doubled haploid lines resistant topotato virus Y0. Indian J. Genet. 2016, 76, 333–340. [Google Scholar] [CrossRef]
- Ferrie, A.M.R.; Bethune, T.D. A microspore embryogenesis protocol for Camelina sativa, a multi-use crop. Plant Cell Tissue Organ Cult. (PCTOC) 2011, 106, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Soorni, J.; Kazemitabar, S.K.; Kahrizi, D.; Dehestani, A.; Bagheri, N. Screening of camelina (Camelina sativa L.) doubled haploid lines for freezing tolerance in the seedling stage. Genetika 2017, 49, 173–181. [Google Scholar] [CrossRef]
- Berti, M.; Wilckens, R.; Fischer, S.; Solis, A.; Johnson, B. Seeding date influence on camelina seed yield, yield components, and oil content in Chile. Ind. Crops Prod. 2011, 34, 1358–1365. [Google Scholar] [CrossRef]
- Anderson, J.V.; Wittenberg, A.; Li, H.; Berti, M.T. High throughput phenotyping of Camelina sativa seeds for crude protein, total oil, and fatty acids profile by near infrared spectroscopy. Ind. Crops Prod. 2019, 137, 501–507. [Google Scholar] [CrossRef]
- Wittenberg, A.; Anderson, J.V.; Berti, M.T. Crop growth and productivity of winter camelina in response to sowing date in the northwestern Corn Belt of the USA. Ind. Crops Prod. 2020, 158, 113036. [Google Scholar] [CrossRef]
- Burgueño, J.; Crossa, J.; Rodríguez, F.; Yeater, K.M. Augmented designs-experimental designs in which all treatments are not replicated. In Applied Statistics in Agricultural, Biological, and Environmental Sciences; Glaz, B., Yeater, K.M., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2018; pp. 345–370. [Google Scholar] [CrossRef]
- Piepho, H.; Williams, E.R. Augmented Row–Column Designs for a Small Number of Checks. Agron. J. 2016, 108, 2256–2262. [Google Scholar] [CrossRef]
- Federer, W.T. Augmented (or hoonuiaku) designs. Hawaii. Sugar Plant. Rec. 1956, 55, 191–208. [Google Scholar]
- Scott, R.A.; Milliken, G.A. A SAS Program for Analyzing Augmented Randomized Complete-Block Designs. Crop Sci. 1993, 33, 865–867. [Google Scholar] [CrossRef]
- Federer, W.T.; Reynolds, M.; Crossa, J. Combining Results from Augmented Designs over Sites. Agron. J. 2001, 93, 389–395. [Google Scholar] [CrossRef] [Green Version]
- Haines, L.M. Augmented Block Designs for Unreplicated Trials. J. Agric. Biol. Environ. Stat. 2021, 26, 409–427. [Google Scholar] [CrossRef]
- Aravind, J.; MukeshSankar, S.; Wankhede, D.P.; Kaur, V. Augmented RCBD: Analysis of Augmented Randomised Complete Block Designs. R Package Version 0.1.2.9000. 2020. Available online: https://www.aravind-j.github.io/augmentedRCBD/https://cran.r-project.org/package=augmentedRCBD (accessed on 2 March 2019).
- Neupane, D.; Solomon, J.K.Q.; Mclennon, E.; Davison, J.; Lawry, T. Sowing date and sowing method influence on camelina cultivars grain yield, oil concentration, and biodiesel production. Food Energy Secur. 2019, 8, e00166. [Google Scholar] [CrossRef] [Green Version]
- Bansal, G.; Zhou, W.; Barlow, P.J.; Lo, H.-L.; Neo, F.-L. Performance of palm olein in repeated deep frying and controlled heating processes. Food Chem. 2010, 121, 338–347. [Google Scholar] [CrossRef]
- López-Martínez, J.C.; Campra-Madrid, P.; Guil-Guerrero, J.L. γ-Linolenic acid enrichment from Borago officinalis and Echium fastuosum seed oils and fatty acids by low temperature crystallization. J. Biosci. Bioeng. 2004, 97, 294–298. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Improved recovery of fatty acid through direct transesterification without prior extraction or purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef]
- AOCS. Nitrogen-Ammonia-Protein, Modified Kjeldahl Method; AOCS Recommended Practice Ba 4a-38/Ba 4b-87; AOCS: Champaign, IL, USA, 1987; p. 2. [Google Scholar]
- Čanak, P.; Jeromela, A.M.; Vujošević, B.; Kiprovski, B.; Mitrović, B.; Alberghini, B.; Facciolla, E.; Monti, A.; Zanetti, F. Is Drought Stress Tolerance Affected by Biotypes and Seed Size in the Emerging Oilseed Crop Camelina? Agronomy 2020, 10, 1856. [Google Scholar] [CrossRef]
- Zanetti, F.; Gesch, R.W.; Walia, M.K.; Johnson, J.M.; Monti, A. Winter camelina root characteristics and yield performance under contrasting environmental conditions. Field Crops Res. 2020, 252, 107794. [Google Scholar] [CrossRef]
- Leclère, M.; Lorent, A.-R.; Jeuffroy, M.-H.; Butier, A.; Chatain, C.; Loyce, C. Diagnosis of camelina seed yield and quality across an on-farm experimental network. Eur. J. Agron. 2020, 122, 126190. [Google Scholar] [CrossRef]
- Kurasiak-Popowska, D.; Tomkowiak, A.; Człopińska, M.; Bocianowski, J.; Weigt, D.; Nawracała, J. Analysis of yield and genetic similarity of Polish and Ukrainian Camelina sativa genotypes. Ind. Crops Prod. 2018, 123, 667–675. [Google Scholar] [CrossRef]
- Załuski, D.; Tworkowski, J.; Krzyżaniak, M.; Stolarski, M.J.; Kwiatkowski, J. The Characterization of 10 Spring Camelina Genotypes Grown in Environmental Conditions in North-Eastern Poland. Agronomy 2020, 10, 64. [Google Scholar] [CrossRef] [Green Version]
- Allen, B.L.; Vigil, M.F.; Jabro, J.D. Camelina Growing Degree Hour and Base Temperature Requirements. Agron. J. 2014, 106, 940–944. [Google Scholar] [CrossRef]
- Obeng, E.; Obour, A.K.; Nelson, N.O.; Moreno, J.A.; Ciampitti, I.A.; Wang, D.; Durrett, T.P. Seed yield and oil quality as affected by Camelina cultivar and planting date. J. Crop Improv. 2019, 33, 202–222. [Google Scholar] [CrossRef]
- Kurasiak-Popowska, D.; Stuper-Szablewska, K. The phytochemical quality of Camelina sativa seed and oil. Acta Agric. Scand. Sect. B Plant Soil Sci. 2019, 70, 39–47. [Google Scholar] [CrossRef]
- Kurasiak-Popowska, D.; Graczyk, M.; Przybylska-Balcerek, A.; Stuper-Szablewska, K. Influence of variety and weather conditions on fatty acid composition of winter and spring Camelina sativa varieties in Poland. Eur. Food Res. Technol. 2021, 247, 465–473. [Google Scholar] [CrossRef]
- Gesch, R.W.; Archer, D.W.; Berti, M.T. Dual Cropping Winter Camelina with Soybean in the Northern Corn Belt. Agron. J. 2014, 106, 1735–1745. [Google Scholar] [CrossRef]
- Sydney, E.B.; Sydney, A.C.N.; de Carvalho, J.C.; Soccol, C.R. Microalgal strain selection for biofuel production. In Biomass, Biofuels and Biochemicals: Biofuels from Algae; Panday, A., Chang, J.S., Soccol, C.R., Lee, D.J., Chisti, Y., Elsevier, B.V., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 59–66. [Google Scholar] [CrossRef]
- Luo, Z.; Brock, J.; Dyer, J.M.; Kutchan, T.; Schachtman, D.; Augustin, M.; Ge, Y.; Fahlgren, N.; Abdel-Haleem, H. Genetic Diversity and Population Structure of a Camelina sativa Spring Panel. Front. Plant Sci. 2019, 10, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zubr, J. Qualitative variation of Camelina sativa seed from different locations. Ind. Crops Prod. 2003, 17, 161–169. [Google Scholar] [CrossRef]
- Wittenberg, A.; Anderson, J.V.; Berti, M.T. Winter and summer annual biotypes of camelina have different morphology and seed characteristics. Ind. Crops Prod. 2019, 135, 230–237. [Google Scholar] [CrossRef]
- Augustin, J.M.; Brock, J.R.; Augustin, M.M.; Wellinghoff, R.L.; Shipp, M.; Higashi, Y.; Kumssa, T.T.; Cahoon, E.B.; Kutchan, T.M. Field performance of terpene-producing Camelina sativa. Ind. Crops Prod. 2019, 136, 50–58. [Google Scholar] [CrossRef]
- Stefanoni, W.; Latterini, F.; Ruiz, J.; Bergonzoli, S.; Palmieri, N.; Pari, L. Assessing the Camelina (Camelina sativa (L.) Crantz) Seed Harvesting Using a Combine Harvester: A Case-Study on the Assessment of Work Performance and Seed Loss. Sustainability 2020, 13, 195. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Df | C12.0 | C14.0 | C16.0 | C16.1 | C18.0 | C18.1 | C18.2 | C18.3 | C20.0 | C20.1 | C20.2 | C20.3 | C22.0 | C22.1 | C22.2 | C22.3 | C24.0 | C24.1 | SFA | MUFA | PUFA | OC | PC | DM | TSW | SY |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st Year | |||||||||||||||||||||||||||
| Block | 7 | ** | ns | ns | ns | ns | ns | ns | ns | * | ns | ns | ** | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ** | ns | ns | ns |
| Check Lines | 2 | * | ** | ** | ns | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| DH Lines | 135 | ns | ns | ** | * | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| 2nd Year | |||||||||||||||||||||||||||
| Block | 7 | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns | * | ns | ns | ns | ** | ns | ns | ns | ns |
| Check Lines | 2 | ns | ns | ** | ns | ns | ** | ** | ** | ** | ** | ** | ** | ** | ** | ns | ** | ns | * | ** | ** | ** | ** | ** | ** | ** | ** |
| DH Lines | 135 | ns | ns | ** | ns | ** | ** | ** | ** | ** | ** | ** | * | ns | ** | ** | ** | ns | * | ** | ns | ** | ** | ** | ** | ** | ** |
| Parameters | Minimum | Maximum | Mean ± SD | CV | ||||
|---|---|---|---|---|---|---|---|---|
| 1st Year | 2nd Year | 1st Year | 2nd Year | 1st Year | 2nd Year | 1st Year | 2nd Year | |
| C12:0 (%) | 0.04 | 0.04 | 0.07 | 0.07 | 0.05 ± 0.01 | 0.05 ± 0.01 | 7.1 | 11.6 |
| C14:0 (%) | 0.08 | 0.08 | 0.13 | 0.13 | 0.10 ± 0.01 | 0.10 ± 0.01 | 7.2 | 7.0 |
| C16:0 (%) | 3.53 | 3.59 | 6.8 | 6.58 | 5.69 ± 0.55 | 5.70 ± 0.49 | 3.0 | 3.4 |
| C16:1 (%) | 0.16 | 0.15 | 0.22 | 0.22 | 0.19 ± 0.01 | 0.19 ± 0.01 | 3.6 | 4.4 |
| C18:0 (%) | 1.52 | 1.51 | 3.39 | 3.03 | 2.44 ± 0.33 | 2.42 ± 0.03 | 2.3 | 3.5 |
| C18:1 (%) | 13.36 | 13.45 | 18.26 | 17.75 | 15.53 ± 0.97 | 15.52 ± 0.93 | 0.5 | 1.4 |
| C18:2 (%) | 15.97 | 16.42 | 22.13 | 21.91 | 19.18 ± 1.29 | 19.22 ± 1.24 | 0.5 | 1.6 |
| C18:3 (%) | 30.04 | 30.05 | 34.98 | 34.16 | 32.28 ± 0.96 | 32.14 ± 0.91 | 0.5 | 0.5 |
| C20:0 (%) | 1.45 | 1.53 | 2.69 | 2.77 | 1.92 ± 0.22 | 1.94 ± 0.21 | 2.5 | 3.6 |
| C20:1 (%) | 13.34 | 13.52 | 16.29 | 16.12 | 14.61 ± 0.55 | 14.69 ± 0.55 | 0.5 | 0.8 |
| C20:2 (%) | 1.14 | 1.21 | 1.82 | 1.78 | 1.51 ± 0.15 | 1.51 ± 0.12 | 3.9 | 3.0 |
| C20:3 (%) | 0.94 | 1.03 | 1.62 | 1.50 | 1.22 ± 0.11 | 1.24 ± 0.09 | 3.1 | 4.7 |
| C22:0 (%) | 0.39 | 0.42 | 0.61 | 0.65 | 0.48 ± 0.04 | 0.51 ± 0.04 | 4.4 | 6.1 |
| C22:1 (%) | 2.50 | 2.39 | 3.71 | 3.49 | 3.01 ± 0.23 | 3.05 ± 0.23 | 1.7 | 4.0 |
| C22:2 (%) | 0.13 | 0.15 | 0.23 | 0.26 | 0.19 ± 0.02 | 0.21 ± 0.02 | 4.5 | 5.4 |
| C22:3 (%) | 0.37 | 0.39 | 0.63 | 0.65 | 0.50 ± 0.06 | 0.53 ± 0.05 | 2.8 | 4.0 |
| C24:0 (%) | 0.20 | 0.18 | 0.35 | 0.32 | 0.27 ± 0.03 | 0.25 ± 0.03 | 4.4 | 9.1 |
| C24:1 (%) | 0.60 | 0.53 | 0.92 | 0.90 | 0.78 ± 0.07 | 0.70 ± 0.08 | 2.4 | 5.5 |
| SFA (%) | 8.90 | 9.04 | 12.93 | 13.05 | 10.95 ± 0.78 | 10.97 ± 0.71 | 1.7 | 1.8 |
| MUFA (%) | 32.05 | 32.13 | 36.88 | 36.62 | 34.11 ± 0.94 | 34.15 ± 0.90 | 0.4 | 1.7 |
| PUFA (%) | 52.29 | 51.70 | 57.7 | 56.97 | 54.89 ± 1.06 | 54.84 ± 0.97 | 0.3 | 0.8 |
| OC (%) | 32.76 | 33.56 | 38.37 | 38.99 | 35.91 ± 1.25 | 36.24 ± 1.20 | 0.3 | 0.5 |
| PC (%) | 25.66 | 25.25 | 30.47 | 29.99 | 27.76 ± 0.91 | 27.36 ± 0.89 | 0.3 | 0.6 |
| DM (day) | 144 | 148 | 218 | 221 | 167 ± 11.8 | 168 ± 11.4 | 1.5 | 2.4 |
| TSW (g) | 0.60 | 0.69 | 1.47 | 1.52 | 1.06 ± 0.19 | 1.12 ± 0.18 | 6.8 | 4.7 |
| SY (kg ha−1) | 783 | 831 | 1636 | 1644 | 1149 ± 194 | 1208 ± 187 | 1.0 | 4.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soorni, J.; Shobbar, Z.-S.; Kahrizi, D.; Zanetti, F.; Sadeghi, K.; Rostampour, S.; Kovács, P.G.; Kiss, A.; Mirmazloum, I. Correlational Analysis of Agronomic and Seed Quality Traits in Camelina sativa Doubled Haploid Lines under Rain-Fed Condition. Agronomy 2022, 12, 359. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020359
Soorni J, Shobbar Z-S, Kahrizi D, Zanetti F, Sadeghi K, Rostampour S, Kovács PG, Kiss A, Mirmazloum I. Correlational Analysis of Agronomic and Seed Quality Traits in Camelina sativa Doubled Haploid Lines under Rain-Fed Condition. Agronomy. 2022; 12(2):359. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020359
Chicago/Turabian StyleSoorni, Jahad, Zahra-Sadat Shobbar, Danial Kahrizi, Federica Zanetti, Kaveh Sadeghi, Sara Rostampour, Péter Gergő Kovács, Attila Kiss, and Iman Mirmazloum. 2022. "Correlational Analysis of Agronomic and Seed Quality Traits in Camelina sativa Doubled Haploid Lines under Rain-Fed Condition" Agronomy 12, no. 2: 359. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020359