
A New Calcium Vectoring Technology: Concentration and Distribution of Ca and Agronomic Efficiency in Pepper Plants

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Localization
2.2. Plant Material and Treatments Description
2.3. Sampling of Plant and Fruit
2.4. Analysis of Plant Material
2.4.1. Chl a Fluorescence Analysis
2.4.2. Determination of the Concentration of Total Ca and Ca Bound to Pectins
2.4.3. Fruit Quality Parameters
2.5. Statistical Analysis
3. Results and Discussion
3.1. Plant Biomass and Photosynthesis
3.2. Ca Concentration in the Different Plant Organs
3.3. Production and Fruit Characteristics
3.4. Fruits Physiopathies
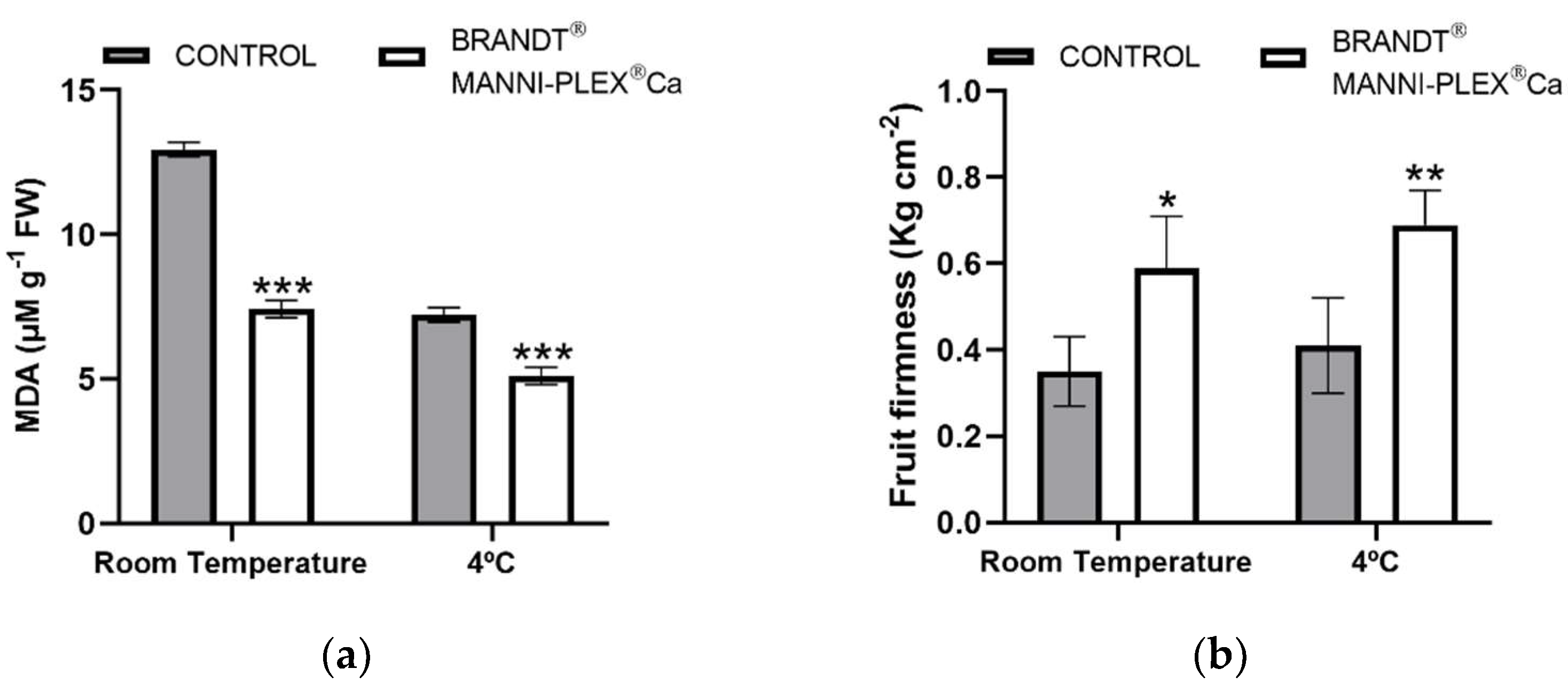
3.5. Post-Harvest Study
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Epstein, E.; Bloom, A.J. Mineral Nutrition of Plants; Principles and Perspective; Sinauer Associates, Inc.: Sunderland, MA, USA, 2005. [Google Scholar]
- De Bang, T.C.; Husted, S.; Laursen, K.H.; Persson, D.P.; Schjoerring, J.K. The Molecular–Physiological Functions of Mineral Macronutrients and Their Consequences for Deficiency Symptoms in Plants. New Phytol. 2021, 229, 2446–2469. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Broadley, M.R. Calcium in Plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Song, W.-P.; Chen, W.; Yi, J.-W.; Wang, H.-C.; Huang, X.-M. Ca Distribution Pattern in Litchi Fruit and Pedicel and Impact of Ca Channel Inhibitor, La3+. Front. Plant Sci. 2018, 8, 2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagassou, D.; Francia, E.; Ronga, D.; Buti, M. Blossom End-Rot in Tomato (Solanum Lycopersicum L.): A Multi-Disciplinary Overview of Inducing Factors and Control Strategies. Sci. Hortic. 2019, 249, 49–58. [Google Scholar] [CrossRef]
- Souri, M.K.; Sooraki, F.Y. Benefits of Organic Fertilizers Spray on Growth Quality of Chili Pepper Seedlings under Cool Temperature. J. Plant Nutr. 2019, 42, 650–656. [Google Scholar] [CrossRef]
- Bonomelli, C.; Fernández, V.; Martiz, J.; Videla, X.; Arias, M.I.; Rojas-Silva, X.; Nario, A. Absorption and Distribution of Root, Fruit, and Foliar-applied 45Ca in ‘Clemenules’ Mandarin Trees. J. Sci. Food Agric. 2020, 100, 4643–4650. [Google Scholar] [CrossRef]
- Bonomelli, C.; Alcalde, C.; Aguilera, C.; Videla, X.; Rojas-Silva, X.; Nario, A.; Fernandez, V. Absorption and Mobility of Radio-Labelled Calcium in Chili Pepper Plants and Sweet Cherry Trees. Sci. Agric. 2021, 78, 1–7. [Google Scholar] [CrossRef]
- Castro, M.J.L.; Ojeda, C.; Cirelli, A.F. Advances in Surfactants for Agrochemicals. Environ. Chem. Lett. 2014, 12, 85–95. [Google Scholar] [CrossRef]
- Larsen, B.; Xu, D.; Halkier, B.A.; Nour-Eldin, H.H. Advances in Methods for Identification and Characterization of Plant Transporter Function. J. Exp. Bot. 2017, 68, 4045–4056. [Google Scholar] [CrossRef]
- Wu, H.; Xu, H.; Marivingt-Mounir, C.; Bonnemain, J.; Chollet, J. Vectorizing Agrochemicals: Enhancing Bioavailability via Carrier-mediated Transport. Pest Manag. Sci. 2019, 75, 1507–1516. [Google Scholar] [CrossRef]
- Nestby, R.; Lieten, F.; Pivot, D.; Lacroix, C.R.; Tagliavini, M. Influence of Mineral Nutrients on Strawberry Fruit Quality and Their Accumulation in Plant Organs. Int. J. Fruit Sci. 2005, 5, 139–156. [Google Scholar] [CrossRef]
- Brown, P.H.; Bellaloui, N.; Wimmer, M.A.; Bassil, E.S.; Ruiz, J.; Hu, H.; Pfeffer, H.; Dannel, F.; Römheld, V. Boron in Plant Biology. Plant Biol. 2002, 4, 205–223. [Google Scholar] [CrossRef]
- Ali, Q.; Shahid, S.; Ali, S.; El-Esawi, M.A.; Hussain, A.I.; Perveen, R.; Iqbal, N.; Rizwan, M.; Nasser Alyemeni, M.; El-Serehy, H.A.; et al. Fertigation of Ajwain (Trachyspermum Ammi L.) with Fe-Glutamate Confers Better Plant Performance and Drought Tolerance in Comparison with FeSO4. Sustainability 2020, 12, 7119. [Google Scholar] [CrossRef]
- Miguel, M.G.; Lourenço, J.P.; Faleiro, M.L. Superparamagnetic Iron Oxide Nanoparticles and Essential Oils: A New Tool for Biological Applications. Int. J. Mol. Sci. 2020, 21, 6633. [Google Scholar] [CrossRef] [PubMed]
- Minchin, P.E.H.; Thorp, T.G.; Boldingh, H.L.; Gould, N.; Cooney, J.M.; Negm, F.B.; Focht, E.; Arpaia, M.L.; Hu, H.; Brown, P. A Possible Mechanism for Phloem Transport of Boron in ‘Hass’ Avocado (Persea Americana Mill.) Trees. J. Hortic. Sci. Biotechnol. 2012, 87, 23–28. [Google Scholar] [CrossRef]
- Hu, H.; Penn, S.G.; Lebrilla, C.B.; Brown, P.H. Isolation and Characterization of Soluble Boron Complexes in Higher Plants (The Mechanism of Phloem Mobility of Boron). Plant Physiol. 1997, 113, 649–655. [Google Scholar] [CrossRef] [Green Version]
- Casas, A. Cultivos Sin Suelo II Curso Superior de Especialización; Fernández, M., Cuadrado, I., Eds.; Dirección General de Investigación y Formación Agraria: Almería, Spain, 1999; pp. 527–566. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Wolf, B. A Comprehensive System of Leaf Analyses and Its Use for Diagnosing Crop Nutrient Status. Commun. Soil Sci. Plant Anal. 1982, 13, 1035–1059. [Google Scholar] [CrossRef]
- Su, Y.; Liu, J.; Lu, Z.; Wang, X.; Zhang, Z.; Shi, G. Effects of Iron Deficiency on Subcellular Distribution and Chemical Forms of Cadmium in Peanut Roots in Relation to Its Translocation. Environ. Exp. Bot. 2014, 97, 40–48. [Google Scholar] [CrossRef]
- Fu, J.; Huang, B. Involvement of Antioxidants and Lipid Peroxidation in the Adaptation of Two Cool-Season Grasses to Localized Drought Stress. Environ. Exp. Bot. 2001, 45, 105–114. [Google Scholar] [CrossRef]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and Ascorbic Acid in Spinach (Spinacia Oleracea) Chloroplasts. The Effect of Hydrogen Peroxide and of Paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef] [Green Version]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Aeasure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Luo, Q.; Sun, M.; Corke, H. Antioxidant Activity and Phenolic Compounds of 112 Traditional Chinese Medicinal Plants Associated with Anticancer. Life Sci. 2004, 74, 2157–2184. [Google Scholar] [CrossRef]
- Maathuis, F.J. Physiological Functions of Mineral Macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Parađiković, N.; Vinković, T.; Vinković Vrček, I.; Žuntar, I.; Bojić, M.; Medić-Šarić, M. Effect of Natural Biostimulants on Yield and Nutritional Quality: An Example of Sweet Yellow Pepper (Capsicum Annuum L.) Plants. J. Sci. Food Agric. 2011, 91, 2146–2152. [Google Scholar] [CrossRef] [PubMed]
- Hochmal, A.K.; Schulze, S.; Trompelt, K.; Hippler, M. Calcium-Dependent Regulation of Photosynthesis. Biochimica et Biophysica Acta—Bioenerg. 2015, 1847, 993–1003. [Google Scholar] [CrossRef] [Green Version]
- Strasser, R.; Srivastava, A.; Tsimilli-Michael, M. The Fluorescence Transient as a Tool to Characterize and Screen Photosynthetic Samples. In Probing Photosynthesis: Mechanism, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor & Francis: London, UK, 2000; pp. 443–480. [Google Scholar]
- Mwije, A.; Hoffman, E.W.; Lötze, E. Apple Peel Biochemical Changes after Foliar Application of Combined Boron and Calcium II. Photosynthetic Pigments, Total Peroxides and Photochemical Efficiency. Am. J. Plant Sci. 2020, 11, 939–964. [Google Scholar] [CrossRef]
- Lester, G.E.; Grusak, M.A. Field Application of Chelated Calcium: Postharvest Effects on Cantaloupe and Honeydew Fruit Quality. HortTechnology 2004, 14, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Lötze, E.; Turketti, S. Efficacy of Foliar Application of Calcium Products on Tomatoes as Defined by Penetration Depth of and Concentration within Fruit Tissues. J. Plant Nutr. 2015, 38, 2112–2125. [Google Scholar] [CrossRef]
- Gigli-Bisceglia, N.; Engelsdorf, T.; Hamann, T. Plant Cell Wall Integrity Maintenance in Model Plants and Crop Species-Relevant Cell Wall Components and Underlying Guiding Principles. Cell. Mol. Life Sci. 2020, 77, 2049–2077. [Google Scholar] [CrossRef] [Green Version]
- Reem, N.T.; Chambers, L.; Zhang, N.; Abdullah, S.F.; Chen, Y.; Feng, G.; Gao, S.; Soto-Burgos, J.; Pogorelko, G.; Bassham, D.C.; et al. Post-Synthetic Reduction of Pectin Methylesterification Causes Morphological Abnormalities and Alterations to Stress Response in Arabidopsis Thaliana. Plants 2020, 9, 1558. [Google Scholar] [CrossRef]
- De Freitas, S.T.; Shackel, K.A.; Mitcham, E.J. Abscisic Acid Triggers Whole-Plant and Fruit-Specific Mechanisms to Increase Fruit Calcium Uptake and Prevent Blossom End Rot Development in Tomato Fruit. J. Exp. Bot. 2011, 62, 2645–2656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, A.; Knoche, M. Calcium and the Physiology of Sweet Cherries: A Review. Sci. Hortic. 2019, 245, 107–115. [Google Scholar] [CrossRef]
- Bai, Q.; Shen, Y.; Huang, Y. Advances in Mineral Nutrition Transport and Signal Transduction in Rosaceae Fruit Quality and Postharvest Storage. Front. Plant Sci. 2021, 12, 68. [Google Scholar] [CrossRef] [PubMed]
- De Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; de Abreu, C.E.B.; Gomes-Filho, E. Effect of Salt Stress on Antioxidative Enzymes and Lipid Peroxidation in Leaves and Roots of Salt-Tolerant and Salt-Sensitive Maize Genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, E.; Rubio-Wilhelmi, M.; Cervilla, L.M.; Blasco, B.; Rios, J.J.; Rosales, M.A.; Romero, L.; Ruiz, J.M. Genotypic Differences in Some Physiological Parameters Symptomatic for Oxidative Stress under Moderate Drought in Tomato Plants. Plant Sci. 2010, 178, 30–40. [Google Scholar] [CrossRef]
- De la Torre-González, A.; Navarro-León, E.; Albacete, A.; Blasco, B.; Ruiz, J.M. Study of Phytohormone Profile and Oxidative Metabolism as Key Process to Identification of Salinity Response in Tomato Commercial Genotypes. J. Plant Physiol. 2017, 216. [Google Scholar] [CrossRef]
- Chen, T.; Ji, D.; Zhang, Z.; Li, B.; Qin, G.; Tian, S. Advances and Strategies for Controlling the Quality and Safety of Postharvest Fruit. Engineering 2021, 7, 1177–1184. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxidant and Antioxidant Signalling in Plants: A Re-Evaluation of the Concept of Oxidative Stress in a Physiological Context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Buettner, G.R.; Jurkiewicz, B.A. Catalytic Metals, Ascorbate and Free Radicals: Combinations to Avoid. Radiat. Res. 1996, 145, 532. [Google Scholar] [CrossRef] [Green Version]
- Eichholzer, M.; Lüthy, J.; Gutzwiller, F.; Stähelin, H.B. The Role of Folate, Antioxidant Vitamins and Other Constituents in Fruit and Vegetables in the Prevention of Cardiovascular Disease: The Epidemiological Evidence. Int. J. Vitam. Nutr. Res. 2001, 71, 5–17. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Control | BRANDT® MANNI-PLEX® Ca | p-Value | HSD0.05 | |
|---|---|---|---|---|
| Fv/Fm | 0.827 ± 0.004 | 0.824 ± 0.004 | NS | 0.03 |
| RC/ABS | 0.86 ± 0.03 | 1.31 ± 0.09 | ** | 0.21 |
| PIABS | 8.36 ± 1.09 | 11.74 ± 1.18 | * | 2.57 |
| Control | BRANDT® MANNI-PLEX® Ca | p-Value | HSD0.05 | ||
|---|---|---|---|---|---|
| Basal leaves | [Ca] tot | 33.92 ± 2.49 | 41.30 ± 2.52 | * | 5.67 |
| [Ca] pec | 23.74 ± 1.58 | 30.97 ± 2.09 | * | 3.98 | |
| Intermediate leaves | [Ca] tot | 30.35 ± 2.09 | 37.63 ± 2.20 | * | 4.32 |
| [Ca] pec | 23.06 ± 1.77 | 30.85 ± 2.98 | * | 3.05 | |
| Young leaves | [Ca] tot | 16.57 ± 0.73 | 20.12 ± 0.83 | ** | 1.47 |
| [Ca] pec | 11.23 ± 0.60 | 15.43 ± 0.62 | ** | 1.23 | |
| Fruit | [Ca] tot | 1.33 ± 0.11 | 2.02 ± 0.16 | ** | 0.27 |
| [Ca] pec | 0.94 ± 0.07 | 1.57 ± 0.13 | ** | 0.18 |
| Control | BRANDT® MANNI-PLEX® Ca | p-Value | HSD0.05 | |
|---|---|---|---|---|
| N° of marketable fruits plant−1 | 3.52 ± 0.66 | 5.05 ± 0.61 | ** | 1.22 |
| Weight of marketable fruits plant−1 (g) | 138.17 ± 22.43 | 202.07 ± 39.61 | * | 60.53 |
| Marketable production (g m−2) | 276.34 ± 36.55 | 404.14 ± 57.32 | * | 94.21 |
| Length (cm) | 11.42 ± 1.35 | 15.29 ± 1.87 | * | 3.56 |
| Firmness (Kg cm−2) | 0.47 ± 0.10 | 0.77 ± 0.11 | * | 0.20 |
| °BRIX (%) | 4.40 ± 0.41 | 4.67 ± 0.35 | NS | 0.82 |
| Titratable acidity (%) | 0.38 ± 0.06 | 0.34 ± 0.04 | NS | 0.13 |
| Temperature | Treatments | Initial Fresh Weight (g fruit−1) | Final Fresh Weight (g fruit−1) | Water Loss (%) |
|---|---|---|---|---|
| Room temperature | Control | 29.04 ± 4.72 | 11.83 ± 1.59 | 59.26 ± 6.03 |
| BRANDT® MANNI-PLEX® Ca | 35.41 ± 4.78 | 30.53 ± 3.16 | 13.78 ± 2.48 | |
| p-value | * | ** | *** | |
| HSD0.05 | 5.35 | 4.31 | 9.33 | |
| 4 °C | Control | 30.27 ± 3.98 | 20.75 ± 2.31 | 31.45 ± 3.17 |
| BRANDT® MANNI-PLEX® Ca | 37.18 ± 4.83 | 34.98 ± 3.07 | 5.92 ± 0.46 | |
| p-value | * | ** | *** | |
| HSD0.05 | 5.73 | 4.32 | 4.96 |
| Temperature | Treatments | °BRIX (%) | Titratable Acidity (%) | Ascorbate (mg g−1 FW) | FRAP (mg g−1 FW) | TEAC (mmol g−1 FW) |
|---|---|---|---|---|---|---|
| Room temperature | Control | 4.70 ± 0.45 | 0.062 ± 0.014 | 1.12 ± 0.06 | 3.03 ± 0.16 | 4.06 ± 0.20 |
| BRANDT® MANNI-PLEX® Ca | 5.33 ± 0.30 | 0.044 ± 0.014 | 1.43 ± 0.05 | 3.77 ± 0.29 | 5.16 ± 0.26 | |
| p-value | NS | NS | ** | ** | *** | |
| HSD0.05 | 0.85 | 0.010 | 0.13 | 0.41 | 0.45 | |
| 4 °C | Control | 4.57 ± 0.36 | 0.034 ± 0.012 | 1.18 ± 0.04 | 3.15 ± 0.29 | 4.60 ± 0.41 |
| BRANDT® MANNI-PLEX® Ca | 4.90 ± 0.28 | 0.034 ± 0.011 | 1.48 ± 0.04 | 3.85 ± 0.29 | 5.48 ± 0.17 | |
| p-value | NS | NS | *** | * | ** | |
| HSD0.05 | 0.73 | 0.011 | 0.09 | 0.62 | 0.63 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro-León, E.; López-Moreno, F.J.; Fernández, M.A.; Maldonado, J.J.; Yánez, J.; Blasco, B.; Ruiz, J.M. A New Calcium Vectoring Technology: Concentration and Distribution of Ca and Agronomic Efficiency in Pepper Plants. Agronomy 2022, 12, 410. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020410
Navarro-León E, López-Moreno FJ, Fernández MA, Maldonado JJ, Yánez J, Blasco B, Ruiz JM. A New Calcium Vectoring Technology: Concentration and Distribution of Ca and Agronomic Efficiency in Pepper Plants. Agronomy. 2022; 12(2):410. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020410
Chicago/Turabian StyleNavarro-León, Eloy, Francisco Javier López-Moreno, Miguel Angel Fernández, Juan Jesús Maldonado, Jose Yánez, Begoña Blasco, and Juan Manuel Ruiz. 2022. "A New Calcium Vectoring Technology: Concentration and Distribution of Ca and Agronomic Efficiency in Pepper Plants" Agronomy 12, no. 2: 410. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020410