
Characterization of Flowering Time in Genebank Accessions of Grain Amaranths and Their Wild Relatives Reveals Signatures of Domestication and Local Adaptation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
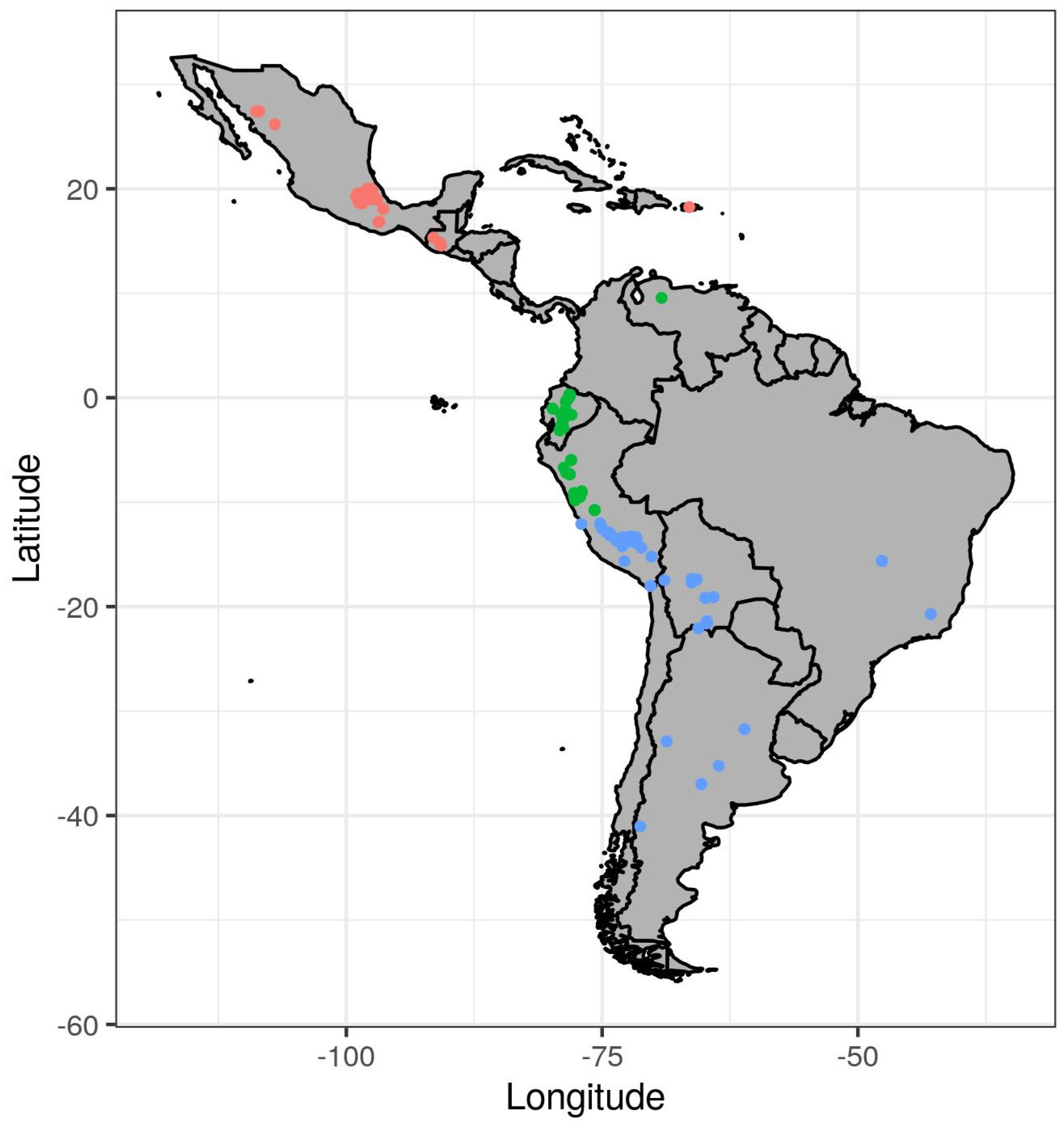
2.1. Plant Material
2.2. Field Experiment
2.3. Phenotypic Traits
2.4. Environmental Variables
2.5. Statistical Analyses
3. Results
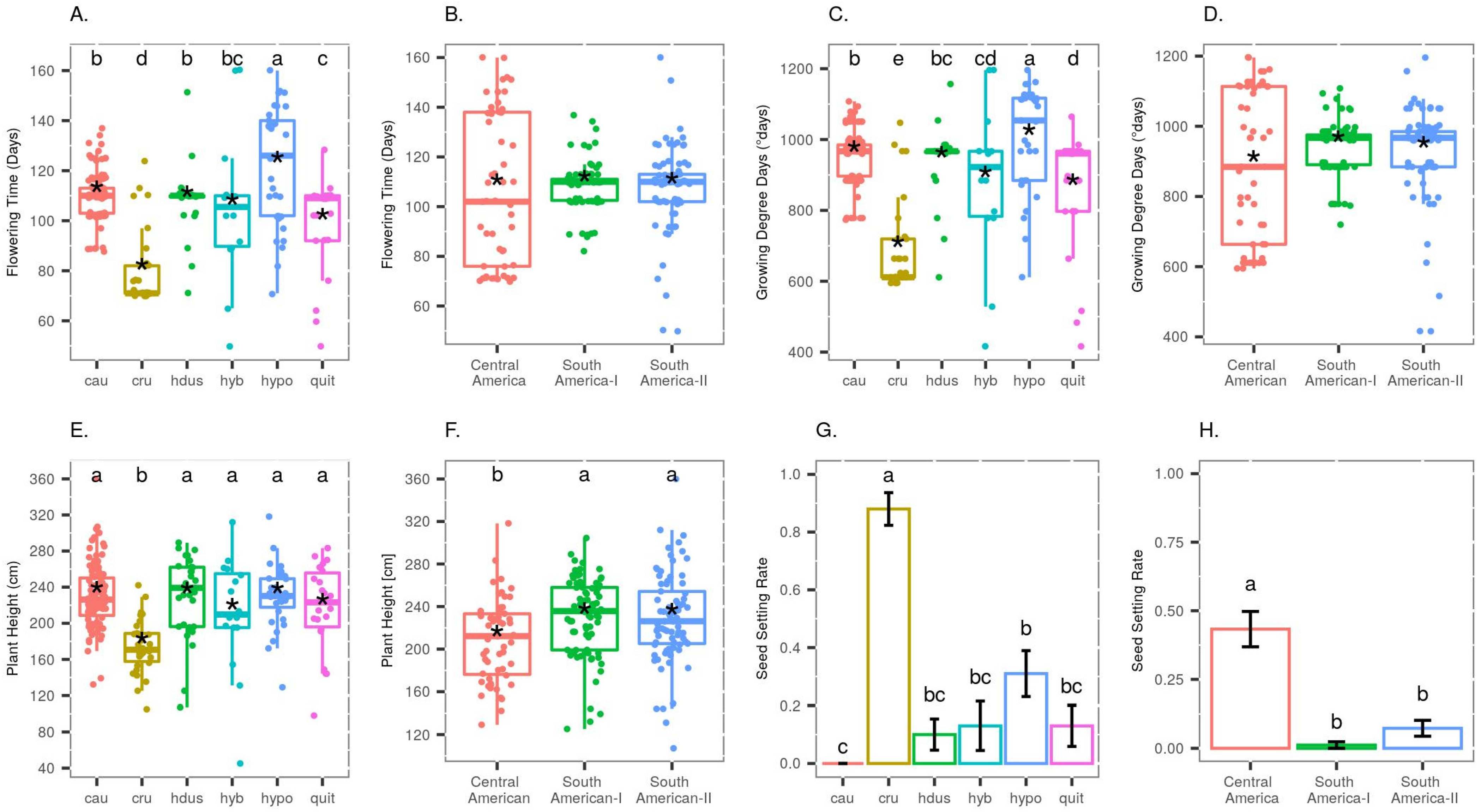
3.1. Variation in Phenotypic Traits and Environmental Variables
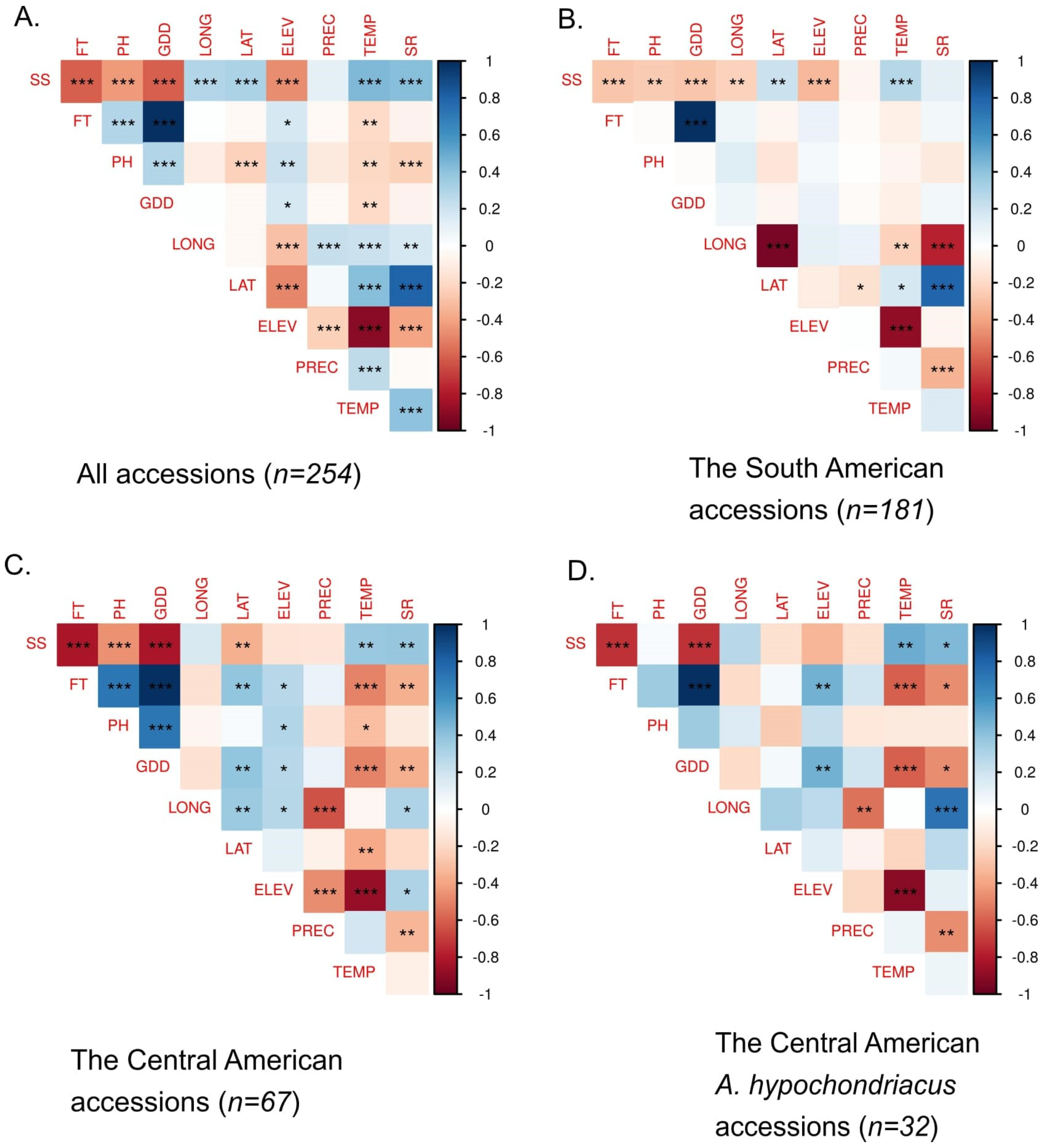
3.2. Relationship between Phenotypic Traits and Environmental Variables
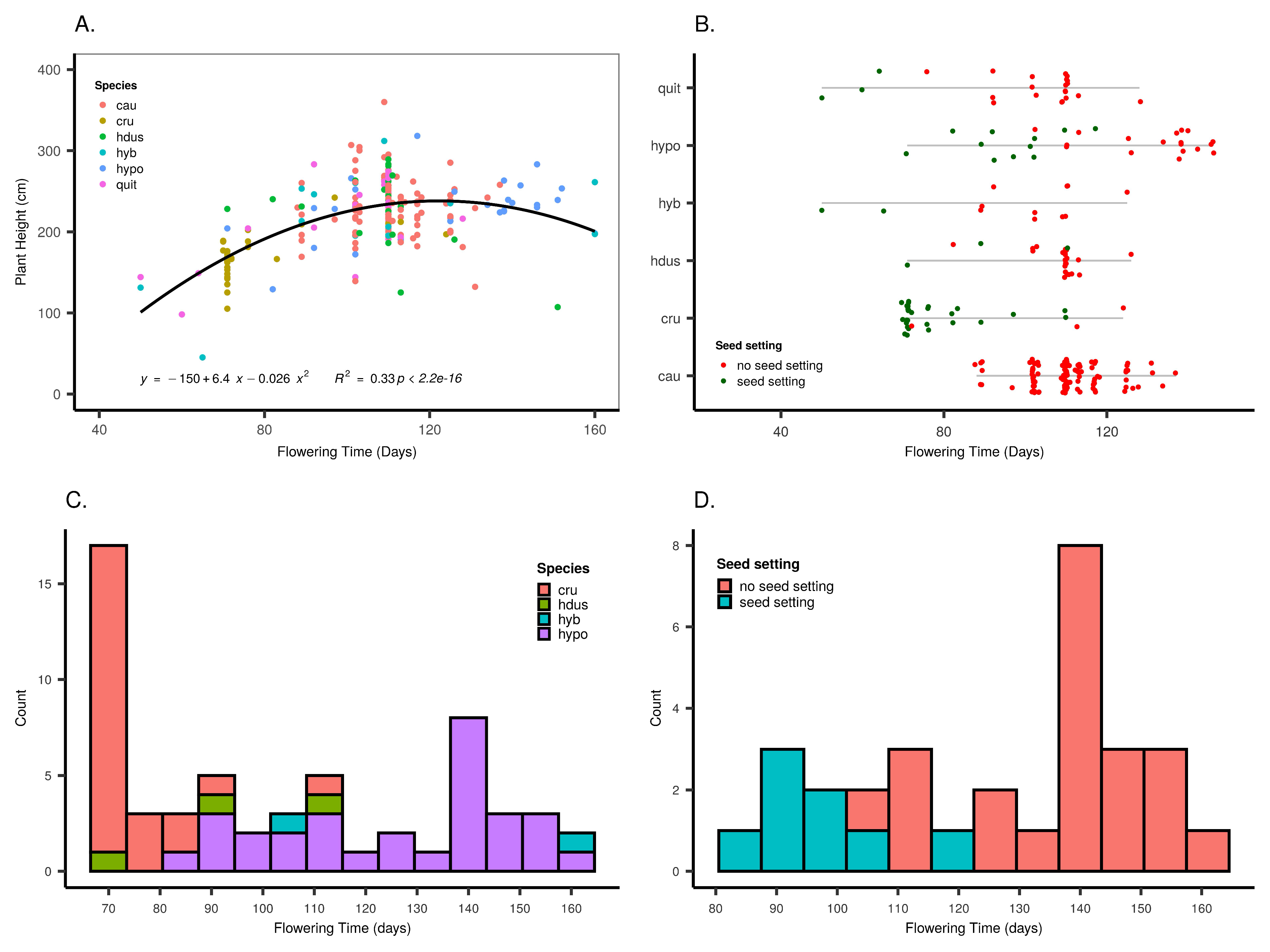
3.3. Factors Influencing Seed Setting
4. Discussion
4.1. Categorization of Photoperiodic Response
4.2. Relationship between Phenotypic Traits and Environmental Variables
4.3. Variation in Seed Setting
4.4. Selection of Early Flowering for Local Adaptation
4.5. Domestication Hypotheses in the Light of Photoperiodic Variation
5. Future Prospects
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Izawa, T. Adaptation of flowering-time by natural and artificial selection in Arabidopsis and rice. J. Exp. Bot. 2007, 58, 3091–3097. [Google Scholar] [CrossRef] [Green Version]
- Lagercrantz, U. At the end of the day: A common molecular mechanism for photoperiod responses in plants? J. Exp. Bot. 2009, 60, 2501–2515. [Google Scholar] [CrossRef] [Green Version]
- Craufurd, P.Q.; Wheeler, T.R. Climate change and the flowering time of annual crops. J. Exp. Bot. 2009, 60, 2529–2539. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liu, W.; Tsegaw, M.; Xu, X.; Qi, Y.; Sapey, E.; Liu, L.; Wu, T.; Sun, S.; Han, T. Principles and practices of the photo-thermal adaptability improvement in soybean. J. Integr. Agric. 2020, 19, 295–310. [Google Scholar] [CrossRef]
- Jung, C.; Müller, A.E. Flowering time control and applications in plant breeding. Trends Plant Sci. 2009, 14, 563–573. [Google Scholar] [CrossRef]
- Ceccarelli, S. Specific adaptation and breeding for marginal conditions. Euphytica 1994, 77, 205–219. [Google Scholar] [CrossRef]
- Herrmann, A.; Rath, J. Biogas Production from Maize: Current State, Challenges, and Prospects. 1. Methane Yield Potential. Bioenergy Res. 2012, 5, 1027–1042. [Google Scholar] [CrossRef]
- Sauer, J.D. The grain amaranths and their relatives: A revised taxonomic and geographic survey. Ann. Mo. Bot. Gard. 1967, 54, 102–137. [Google Scholar] [CrossRef]
- National Research Council. Amaranth Modern Prospects for an Ancient Crop; National Academy Press: Washington, DC, USA, 1984; Volume 1, ISBN 9788578110796. [Google Scholar]
- Costea, M.; Tardif, F.J. The biology of Canadian weeds. 130. Amaranthus retroflexus L., A. powellii S. Watson and A. hybridus L. (update). Can. J. Plant Sci. 2004, 84, 631–668. [Google Scholar]
- Chaudhari, S.; Jordan, D.L.; York, A.C.; Jennings, K.M.; Cahoon, C.W.; Chandi, A.; Inman, M.D. Biology and Management of Glyphosate-Resistant and Glyphosate-Susceptible Palmer Amaranth (Amaranthus palmeri) Phenotypes from a Segregating Population. Weed Sci. 2017, 65, 755–768. [Google Scholar] [CrossRef]
- Becker, R.; Wheeler, E.L.; Lorenz, K.; Stafford, A.E.; Grosjean, O.K.; Betschart, A.A.; Saunders, R.M. A Compositional Study of Amaranth Grain. J. Food Sci. 1981, 46, 1175–1180. [Google Scholar] [CrossRef]
- Stallknecht, G.F.; Schulz-Schaeffer, J.R. Amaranth Rediscovered. In New Crops; Janick, J., Simon, J.E., Eds.; Wiley: New York, NY, USA, 1993; pp. 211–218. [Google Scholar]
- Brenner, D.M.; Baltensperger, D.D.; Kulakow, P.A.; Lehmann, J.W.; Myers, R.L.; Slabbert, M.M.; Sleugh, B.B. Genetic resources and breeding of Amaranthus. Plant Breed. Rev. 2000, 19, 227–285. [Google Scholar] [CrossRef]
- Liu, F.; Stützel, H. Biomass partitioning, specific leaf area, and water use efficiency of vegetable amaranth (Amaranthus spp.) in response to drought stress. Sci. Hortic. 2004, 102, 15–27. [Google Scholar] [CrossRef]
- von Cossel, M.; Möhring, J.; Kiesel, A.; Lewandowski, I. Methane yield performance of amaranth (Amaranthus hypochondriacus L.) and its suitability for legume intercropping in comparison to maize (Zea mays L.). Ind. Crops Prod. 2017, 103, 107–121. [Google Scholar] [CrossRef]
- Jain, S.K.; Hauptli, H.; Valdya, K.R. Outcrossing rate in grain amaranths. J. Hered. 1982, 73, 71–72. [Google Scholar] [CrossRef]
- Hauptli, H.; Jain, S. Genetic variation in outcrossing rate and correlated floral traits in a population of grain amaranth (Amaranthus cruentus L.). Genetica 1985, 66, 21–27. [Google Scholar] [CrossRef]
- Wu, H.; Sun, M.; Yue, S.; Sun, H.; Cai, Y.; Huang, R.; Brenner, D.; Corke, H. Field evaluation of an Amaranthus genetic resource collection in China. Genet. Resour. Crop Evol. 2000, 47, 43–53. [Google Scholar] [CrossRef]
- Mursec, B.; Vindis, P.; Janzekovic, M.; Brus, M.; Cus, F. Analysis of different substrates for processing into biogas Manufacturing and processing. J. Achieve Mater. Manuf. Eng. 2009, 37, 652–659. [Google Scholar]
- Pospišil, A. Yield and Quality of Forage Sorghum and Different Amaranth Species (Amaranthus spp.) Biomass. Agric. Conspec. Sci. 2009, 74, 85–89. [Google Scholar]
- Svirskis, A. Prospects for non-traditional plant species cultivated for forage in Lithuania. Not. Bot. Horti Agrobot. Cluj-Napoca 2009, 37, 215–218. [Google Scholar]
- Balodis, O.; Bartuševics, J.; Gaile, Z. Biomass yield of different plants for biogas production. In Environment. Technologies. Resources. Proceedings of the 8th International Scientific and Practical Conference, Rezekne, Latvia, 20–22 June 2011; Rezekne Academy of Technologies: Rēzekne, Latvia, 2011; Volume 1, pp. 238–245. ISBN 978-9984-44-070-5. [Google Scholar]
- Seppälä, M.; Laine, A.; Rintala, J. Screening of novel plants for biogas production in northern conditions. Bioresour. Technol. 2013, 139, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Sitkey, V.; Gaduš, J.; Kliský, Ľ.; Dudák, A. Biogas Production From Amaranth Biomass. Acta Reg. Environ. 2013, 10, 59–62. [Google Scholar] [CrossRef] [Green Version]
- Baturaygil, A.; Stetter, M.G.; Schmid, K. Breeding Amaranth for Biomass: Evaluating Dry Matter Content and Biomass Potential in Early and Late Maturing Genotypes. Agronomy 2021, 11, 970. [Google Scholar] [CrossRef]
- Kulakow, P.A.; Jain, S.K. The inheritance of flowering time in Amaranthus species. J. Genet. 1985, 64, 85–100. [Google Scholar] [CrossRef]
- Andini, R.; Sulaiman, M.I.; Moulana, R.; Hmon, K.P.W.; Ohsawa, R. Application of principle component analysis in differentiating the three types of Amaranthus based on their photoperiodic flowering response. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Banda Aceh, Indonesia, 2020; Volume 425, p. 012005. [Google Scholar] [CrossRef]
- Rivelli, A.R.; Gherbin, P.; de Maria, S.; Pizza, S. Field evaluation of Amaranthus species for seed and biomass yields in southern Italy. Ital. J. Agron. 2008, 3, 225–229. [Google Scholar] [CrossRef] [Green Version]
- Hoidal, N.; Díaz Gallardo, M.; Jacobsen, S.E.; Alandia, G. Amaranth as a dual-use crop for leafy greens and seeds: Stable responses to leaf harvest across genotypes and environments. Front. Plant Sci. 2019, 10, 817. [Google Scholar] [CrossRef]
- Stetter, M.G.; Vidal-Villarejo, M.; Schmid, K.J. Parallel Seed Color Adaptation during Multiple Domestication Attempts of an Ancient New World Grain. Mol. Biol. Evol. 2020, 37, 1407–1419. [Google Scholar] [CrossRef]
- Stetter, M.G.; Schmid, K.J. Analysis of phylogenetic relationships and genome size evolution of the Amaranthus genus using GBS indicates the ancestors of an ancient crop. Mol. Phylogenet. Evol. 2017, 109, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Kietlinski, K.D.; Jimenez, F.; Jellen, E.N.; Maughan, P.J.; Smith, S.M.; Pratt, D.B. Relationships between the weedy Amaranthus hybridus (Amaranthaceae) and the grain amaranths. Crop Sci. 2014, 54, 220–228. [Google Scholar] [CrossRef]
- Escobedo-López, D.; Núñez-Colín, C.A.; Espitia-Rangel, E. Adaptation of Cultivated Amaranth (Amaranthus spp.) and Their Wild Relatives in Mexico. J. Crop Improv. 2014, 28, 203–213. [Google Scholar] [CrossRef]
- Lightfoot, D.J.; Jarvis, D.E.; Ramaraj, T.; Lee, R.; Jellen, E.N.; Maughan, P.J. Single-molecule sequencing and Hi-C-based proximity-guided assembly of amaranth (Amaranthus hypochondriacus) chromosomes provide insights into genome evolution. BMC Biol. 2017, 15, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahriman, F.; Aktaş, F.; Songur, U.; Şerment, M.; Egesel, C.Ö. Screening Turkish maize landraces for kernel oil content and oil quality traits. Plant Genet. Resour. Characterisation Util. 2020, 18, 278–286. [Google Scholar] [CrossRef]
- Grolemund, G.; Wickham, H. Dates and Times Made Easy with {lubridate}. J. Stat. Softw. 2011, 40, 1–25. [Google Scholar] [CrossRef]
- Horak, M.J.; Loughin, T.M. Growth Analysis of Four Amaranthus Species. Weed Sci. 2000, 48, 347–355. [Google Scholar] [CrossRef]
- Nowosad, J. Pollen: Analysis of Aerobiological Data. R Package Version 0.7.1. 2019. Available online: https://cran.r-project.org/web/packages/pollen/index.html (accessed on 3 December 2021).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 3.4-13. 2020. Available online: https://CRAN.R-project.org/package=raster (accessed on 22 January 2022).
- Kauffman, C.S. Realizing the potential of grain amaranth. Food Rev. Int. 1992, 8, 5–21. [Google Scholar] [CrossRef]
- Zeileis, A.; Hothorn, T. Diagnostic Checking in Regression Relationships. R News 2002, 2, 7–10. [Google Scholar]
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.3-3. 2020. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 18 November 2016).
- Harell, F.E. Hmisc: Harrell Miscellaneous. R Package Version 4.1-1. 2018. Available online: http://cran.nexr.com/web/packages/Hmisc/index.html (accessed on 3 January 2018).
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix. R Package Version 0.90. 2017. Available online: https://cran.r-project.org/web/packages/corrplot/corrplot.pdf (accessed on 11 December 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Liu, L.; Zhang, Y.; Yu, H. Florigen trafficking integrates photoperiod and temperature signals in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 1385–1398. [Google Scholar] [CrossRef]
- Roux, F.; Touzet, P.; Cuguen, J.L.; Le Corre, V. How to be early flowering: An evolutionary perspective. Trends Plant Sci. 2006, 11, 375–381. [Google Scholar] [CrossRef]
- Matsubara, K.; Yamanouchi, U.; Nonoue, Y.; Sugimoto, K.; Wang, Z.X.; Minobe, Y.; Yano, M. Ehd3, encoding a plant homeodomain finger-containing protein, is a critical promoter of rice flowering. Plant J. 2011, 66, 603–612. [Google Scholar] [CrossRef]
- Hung, H.Y.; Shannon, L.M.; Tian, F.; Bradbury, P.J.; Chen, C.; Flint-Garcia, S.A.; McMullen, M.D.; Ware, D.; Buckler, E.S.; Doebley, J.F.; et al. ZmCCT and the genetic basis of day-length adaptation underlying the postdomestication spread of maize. Proc. Natl. Acad. Sci. USA 2012, 109, E1913–E1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, K.M.; Wendel, J.F. Crop plants as models for understanding plant adaptation and diversification. Front. Plant Sci. 2013, 4, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, R.L.; Morishige, D.T.; Brady, J.A.; Rooney, W.L.; Yang, S.; Klein, P.E.; Mullet, J.E. Ghd7 (Ma6) Represses Sorghum Flowering in Long Days: Ghd7 Alleles Enhance Biomass Accumulation and Grain Production. Plant Genome 2014, 7. [Google Scholar] [CrossRef]
- Gaudinier, A.; Blackman, B.K. Evolutionary processes from the perspective of flowering time diversity. New Phytol. 2019, 225, 1883–1898. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Josephs, E.B.; Lee, K.M.; Roberts, L.M.; Rellán-Álvarez, R.; Ross-Ibarra, J.; Hufford, M.B. Molecular Parallelism Underlies Convergent Highland Adaptation of Maize Landraces. Mol. Biol. Evol. 2021, 38, 3567–3580. [Google Scholar] [CrossRef] [PubMed]
- Lewandowska-Sabat, A.M.; Fjellheim, S.; Olsen, J.E.; Rognli, O.A. Local Populations of Arabidopsis thaliana Show Clear Relationship between Photoperiodic Sensitivity of Flowering Time and Altitude. Front. Plant Sci. 2017, 8, 1046. [Google Scholar] [CrossRef] [Green Version]
- Swarts, K.; Bauer, E.; Glaubitz, J.C.; Ho, T.; Johnson, L.; Li, Y.; Li, Y.; Miller, Z.; Romay, C.; Schön, C.C.; et al. Joint analysis of days to flowering reveals independent temperate adaptations in maize. Heredity 2021, 126, 929–941. [Google Scholar] [CrossRef]
- Espitia Rangel, E.; Mapes Sánchez, E.; Núñez-Colín, C.A.; Escobedo López, D. Geographical distribution of cultivated species of Amaranthus and their wild relatives in Mexico Eduardo. Rev. Mex. Cienc. Agrícolas 2010, 1, 427–437. [Google Scholar]
- Jähne, F.; Hahn, V.; Würschum, T.; Leiser, W.L. Speed breeding short-day crops by LED-controlled light schemes. Theor. Appl. Genet. 2020, 133, 2335–2342. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baturaygil, A.; Schmid, K. Characterization of Flowering Time in Genebank Accessions of Grain Amaranths and Their Wild Relatives Reveals Signatures of Domestication and Local Adaptation. Agronomy 2022, 12, 505. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020505
Baturaygil A, Schmid K. Characterization of Flowering Time in Genebank Accessions of Grain Amaranths and Their Wild Relatives Reveals Signatures of Domestication and Local Adaptation. Agronomy. 2022; 12(2):505. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020505
Chicago/Turabian StyleBaturaygil, Ali, and Karl Schmid. 2022. "Characterization of Flowering Time in Genebank Accessions of Grain Amaranths and Their Wild Relatives Reveals Signatures of Domestication and Local Adaptation" Agronomy 12, no. 2: 505. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12020505