
Antimicrobial Potential of Essential Oils from Aromatic Plant Ocimum sp.; A Comparative Biochemical Profiling and In-Silico Analysis

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Essential Oil Used
2.2. Chemical Characterization of EOs
2.3. Antimicrobial Activity
2.4. Antioxidant Activity Assay
2.5. Molecular Docking Analysis
3. Results and Discussion
3.1. Biochemical Characterization of EOs
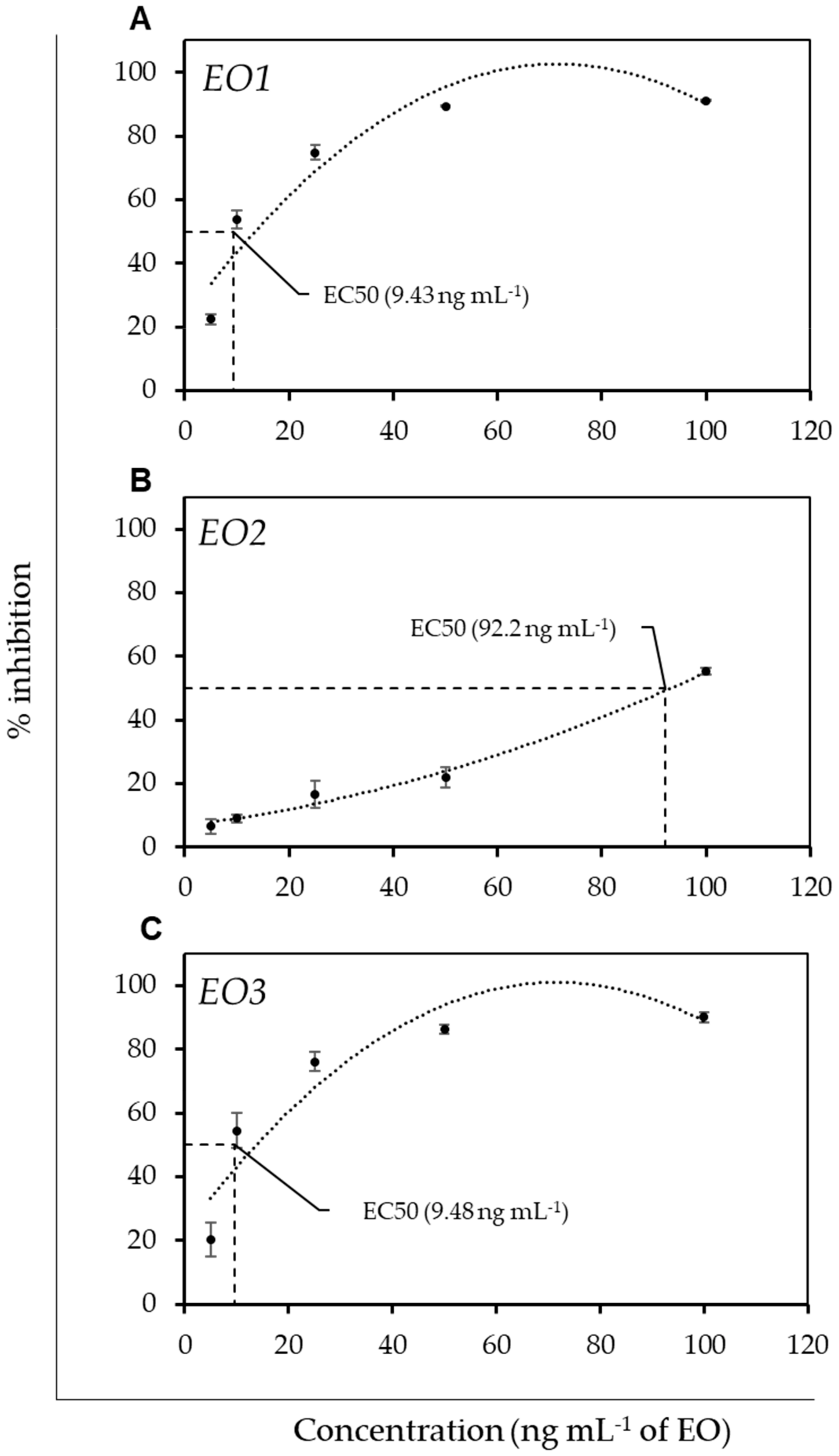
3.2. Antimicrobial Activity and Antioxidant Activity of EOs
3.3. In-Silico Evaluation of Antimicrobial Potential of EO
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tripathi, P.; Singh, A. Chapter 19—Biofertilizers: “An Ace in the Hole” in Medicinal and Aromatic Plants Cultivation; Rakshit, A., Meena, V.S., Parihar, M., Singh, H.B., Singh, A.K.B.T.-B., Eds.; Woodhead Publishing: Sawston, UK, 2021; Volume 1, pp. 253–263. ISBN 978-0-12-821667-5. [Google Scholar]
- Aftab, T. A review of medicinal and aromatic plants and their secondary metabolites status under abiotic stress. J. Med. Plants 2019, 7, 99–106. [Google Scholar]
- Wallace, R.J. Antimicrobial properties of plant secondary metabolites. Proc. Nutr. Soc. 2004, 63, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G.; Scheffer, J.J.C. Factors affecting secondary metabolite production in plants: Volatile components and essential oils. Flavour Fragr. J. 2008, 23, 213–226. [Google Scholar] [CrossRef]
- Rehman, R.; Hanif, M.A.; Mushtaq, Z.; Al-Sadi, A.M. Biosynthesis of essential oils in aromatic plants: A review. Food Rev. Int. 2016, 32, 117–160. [Google Scholar] [CrossRef]
- Gounaris, Y. Biotechnology for the production of essential oils, flavours and volatile isolates. A review. Flavour Fragr. J. 2010, 25, 367–386. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Kohli, R.K.; Kaur, S. Eucalyptus essential oil as a natural pesticide. For. Ecol. Manag. 2008, 256, 2166–2174. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Matasyoh, J.C.; Kiplimo, J.J.; Karubiu, N.M.; Hailstorks, T.P. Chemical composition and antimicrobial activity of essential oil of Tarchonanthus camphoratus. Food Chem. 2007, 101, 1183–1187. [Google Scholar] [CrossRef]
- Hazzoumi, Z.; Moustakime, Y.; Joutei, K.A. Effect of arbuscular mycorrhizal fungi (AMF) and water stress on growth, phenolic compounds, glandular hairs, and yield of essential oil in basil (Ocimum gratissimum L). Chem. Biol. Technol. Agric. 2015, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Kumar, B.; Bajpai, V.; Tiwari, S.; Pandey, R. Phytochemistry of Plants of Genus Ocimum, 1st ed.; CRC Press: Boca Raton, FL, USA; New York, NY, USA, 2020; pp. 1–77. ISBN 1003014852. [Google Scholar]
- Singh, D.; Chaudhuri, P.K. A review on phytochemical and pharmacological properties of Holy basil (Ocimum sanctum L.). Ind. Crops Prod. 2018, 118, 367–382. [Google Scholar] [CrossRef]
- Rastogi, S.; Shah, S.; Kumar, R.; Vashisth, D.; Akhtar, M.Q.; Kumar, A.; Dwivedi, U.N.; Shasany, A.K. Ocimum metabolomics in response to abiotic stresses: Cold, flood, drought and salinity. PLoS ONE 2019, 14, e0210903. [Google Scholar] [CrossRef]
- Rastogi, S.; Shah, S.; Kumar, R.; Kumar, A.; Shasany, A.K. Comparative temporal metabolomics studies to investigate interspecies variation in three Ocimum species. Sci. Rep. 2020, 10, 5234. [Google Scholar] [CrossRef]
- Reddy, D.N. Essential oils extracted from medicinal plants and their applications. In Natural Bio-Active Compounds, 1st ed.; Akhtar, M., Swamy, M., Sinniah, U., Eds.; Springer: Singapore, 2019; Volume 1, pp. 237–283. [Google Scholar]
- Dhama, K.; Sharun, K.; Gugjoo, M.B.; Tiwari, R.; Alagawany, M.; Iqbal Yatoo, M.; Thakur, P.; Iqbal, H.M.N.; Chaicumpa, W.; Michalak, I. A comprehensive review on chemical profile and pharmacological activities of Ocimum basilicum. Food Rev. Int. 2021, 1–29. [Google Scholar] [CrossRef]
- Kaur, H.; Manna, M.; Thakur, T.; Gautam, V.; Salvi, P. Imperative role of sugar signaling and transport during drought stress responses in plants. Physiol. Plant. 2021, 171, 833–848. [Google Scholar] [CrossRef]
- Salvi, P.; Manna, M.; Kaur, H.; Thakur, T.; Gandass, N.; Bhatt, D.; Muthamilarasan, M. Phytohormone signaling and crosstalk in regulating drought stress response in plants. Plant Cell Rep. 2021, 40, 1305–1329. [Google Scholar] [CrossRef]
- Manna, M.; Thakur, T.; Chirom, O.; Mandlik, R.; Deshmukh, R.; Salvi, P. Transcription factors as key molecular target to strengthen the drought stress tolerance in plants. Physiol. Plant. 2021, 172, 847–868. [Google Scholar] [CrossRef]
- Dresselhaus, T.; Hückelhoven, R. Biotic and abiotic stress responses in crop plants. Agronomy 2018, 8, 267. [Google Scholar] [CrossRef] [Green Version]
- Salvi, P.; Agarrwal, R.; Kajal; Gandass, N.; Manna, M.; Kaur, H.; Deshmukh, R. Sugar transporters and their molecular tradeoffs during abiotic stress responses in plants. Physiol. Plant. 2022, e13652. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Mohsin, S.M.; Bhuyan, M.H.M.B.; Bhuiyan, T.F.; Anee, T.I.; Masud, A.A.C.; Nahar, K. Phytotoxicity, environmental and health hazards of herbicides: Challenges and ways forward. In Agrochemicals Detection, Treatment and Remediation; Elsevier: Amsterdam, The Netherlands, 2020; pp. 55–99. [Google Scholar]
- AL-Ahmadi, M.S. Pesticides, anthropogenic activities, and the health of our environment safety. In Pesticides-Use and Misuse and Their Impact in the Environment; Larramendy, M., Soloneski, S., Eds.; IntechOpen: London, UK, 2019; pp. 1–23. [Google Scholar]
- Duran-Lara, E.F.; Valderrama, A.; Marican, A. Natural organic compounds for application in organic farming. Agriculture 2020, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Rothmann, L.A.; McLaren, N.W. Sclerotinia sclerotiorum disease prediction: A review and potential applications in South Africa. S. Afr. J. Sci. 2018, 114, 1–9. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, C.A.; Belt, K.; Thatcher, L.F. Tackling control of a cosmopolitan phytopathogen: Sclerotinia. Front. Plant Sci. 2021, 12, 1764. [Google Scholar] [CrossRef] [PubMed]
- Peltier, A.J.; Bradley, C.A.; Chilvers, M.I.; Malvick, D.K.; Mueller, D.S.; Wise, K.A.; Esker, P.D. Biology, yield loss and control of Sclerotinia stem rot of soybean. J. Integr. Pest Manag. 2012, 3, B1–B7. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.P. Identification of Essential oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation Carol Stream: Carol Stream, IL, USA, 2007; Volume 456. [Google Scholar]
- Joshi, R.K. Chemical composition, in vitro antimicrobial and antioxidant activities of the essential oils of Ocimum gratissimum, O. sanctum and their major constituents. Indian J. Pharm. Sci. 2013, 75, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddharth, S.; Vittal, R.R. Evaluation of antimicrobial, enzyme inhibitory, antioxidant and cytotoxic activities of partially purified volatile metabolites of marine Streptomyces sp. S2A. Microorganisms 2018, 6, 72. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-Pdb Viewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Stefan, M.; Zamfirache, M.M.; Padurariu, C.; Trută, E.; Gostin, I. The composition and antibacterial activity of essential oils in three Ocimum species growing in Romania. Cent. Eur. J. Biol. 2013, 8, 600–608. [Google Scholar] [CrossRef]
- Hzounda, J.B.F.; Jazet, P.M.D.; Lazar, G.; Răducanu, D.; Caraman, I.; Bassene, E.; Boyom, F.F.; Lazar, I.M. Spectral and chemometric analyses reveal antioxidant properties of essential oils from four Cameroonian Ocimum. Ind. Crops Prod. 2016, 80, 101–108. [Google Scholar] [CrossRef]
- Schulz, H.; Schrader, B.; Quilitzsch, R.; Pfeffer, S.; Krüger, H. Rapid classification of basil chemotypes by various vibrational spectroscopy methods. J. Agric. Food Chem. 2003, 51, 2475–2481. [Google Scholar] [CrossRef]
- Schulz, H.; Baranska, M. Identification and quantification of valuable plant substances by IR and Raman spectroscopy. Vib. Spectrosc. 2007, 43, 13–25. [Google Scholar] [CrossRef]
- Jadhav, S.; Shah, R.; Bhave, M.; Palombo, E.A. Inhibitory activity of yarrow essential oil on Listeria planktonic cells and biofilms. Food Control 2013, 29, 125–130. [Google Scholar] [CrossRef]
- Khan, A.; Ahmad, A.; Akhtar, F.; Yousuf, S.; Xess, I.; Khan, L.A.; Manzoor, N. Ocimum sanctum essential oil and its active principles exert their antifungal activity by disrupting ergosterol biosynthesis and membrane integrity. Res. Microbiol. 2010, 161, 816–823. [Google Scholar] [CrossRef]
- Žabka, M.; Pavela, R.; Kovaříková, K.; Tříska, J.; Vrchotová, N.; Bednář, J. Antifungal and Insecticidal Potential of the Essential Oil from Ocimum sanctum L. against Dangerous Fungal and Insect Species and Its Safety for Non-Target Useful Soil Species Eisenia fetida (Savigny, 1826). Plants 2021, 10, 2180. [Google Scholar] [CrossRef]
- Raut, J.S.; Karuppayil, S.M. A status review on the medicinal properties of essential oils. Ind. Crops Prod. 2014, 62, 250–264. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- Hara, M.; Yamauchi, N.; Sumita, Y. Monoterpenes induce the heat shock response in Arabidopsis. Z. Für Naturforsch. C 2018, 73, 177–184. [Google Scholar] [CrossRef]
- Hara, M.; Harazaki, A.; Tabata, K. Administration of isothiocyanates enhances heat tolerance in Arabidopsis thaliana. Plant Growth Regul. 2013, 69, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, Y.; Kunishima, M.; Mizutani, M.; Sugimoto, Y. Reactive short-chain leaf volatiles act as powerful inducers of abiotic stress-related gene expression. Sci. Rep. 2015, 5, 8030. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Mishra, D.; Shekhar, S.; Singh, D.; Chakraborty, S.; Chakraborty, N. Heat shock proteins and abiotic stress tolerance in plants. In Regulation of Heat Shock Protein Responses; Asea, A., Kaur, P., Eds.; Springer: Singapore, 2018; Volume 13, pp. 41–69. [Google Scholar]
- Kaur, H.; Petla, B.P.; Kamble, N.U.; Singh, A.; Rao, V.; Salvi, P.; Ghosh, S.; Majee, M. Differentially expressed seed aging responsive heat shock protein OsHSP18.2 implicates in seed vigor, longevity and improves germination and seedling establishment under abiotic stress. Front. Plant Sci. 2015, 6, 713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thirugnanasampandan, R.; Jayakumar, R. Protection of cadmium chloride induced DNA damage by Lamiaceae plants. Asian Pac. J. Trop. Biomed. 2011, 1, 391–394. [Google Scholar] [CrossRef] [Green Version]
- Güez, C.M.; de Souza, R.O.; Fischer, P.; Leão, M.F.d.M.; Duarte, J.A.; Boligon, A.A.; Athayde, M.L.; Zuravski, L.; de Oliveira, L.F.S.; Machado, M.M. Evaluation of basil extract (Ocimum basilicum L.) on oxidative, anti-genotoxic and anti-inflammatory effects in human leukocytes cell cultures exposed to challenging agents. Braz. J. Pharm. Sci. 2017, 53, e15098. [Google Scholar] [CrossRef] [Green Version]
- Kabuto, H.; Tada, M.; Kohno, M. Eugenol [2-methoxy-4-(2-propenyl) phenol] prevents 6-hydroxydopamine-induced dopamine depression and lipid peroxidation inductivity in mouse striatum. Biol. Pharm. Bull. 2007, 30, 423–427. [Google Scholar] [CrossRef] [Green Version]
- Perigo, C.V.; Torres, R.B.; Bernacci, L.C.; Guimaraes, E.F.; Haber, L.L.; Facanali, R.; Vieira, M.A.R.; Quecini, V.; Marques, M.O.M. The chemical composition and antibacterial activity of eleven Piper species from distinct rainforest areas in Southeastern Brazil. Ind. Crops Prod. 2016, 94, 528–539. [Google Scholar] [CrossRef]
- Salehi, B.; Upadhyay, S.; Erdogan Orhan, I.; Kumar Jugran, A.; Jayaweera, S.L.D.; Dias, D.A.; Sharopov, F.; Taheri, Y.; Martins, N.; Baghalpour, N. Therapeutic potential of α-and β-pinene: A miracle gift of nature. Biomolecules 2019, 9, 738. [Google Scholar] [CrossRef] [Green Version]
- Singh, G.; Marimuthu, P.; De Heluani, C.S.; Catalan, C.A.N. Antioxidant and biocidal activities of Carum nigrum (seed) essential oil, oleoresin, and their selected components. J. Agric. Food Chem. 2006, 54, 174–181. [Google Scholar] [CrossRef]
- Sabulal, B.; Dan, M.; Kurup, R.; Pradeep, N.S.; Valsamma, R.K.; George, V. Caryophyllene-rich rhizome oil of Zingiber nimmonii from South India: Chemical characterization and antimicrobial activity. Phytochemistry 2006, 67, 2469–2473. [Google Scholar] [CrossRef]
- Reece, R.J.; Maxwell, A. DNA gyrase: Structure and function. Crit. Rev. Biochem. Mol. Biol. 1991, 26, 335–375. [Google Scholar] [CrossRef]
- Collin, F.; Karkare, S.; Maxwell, A. Exploiting bacterial DNA gyrase as a drug target: Current state and perspectives. Appl. Microbiol. Biotechnol. 2011, 92, 479–497. [Google Scholar] [CrossRef] [Green Version]
- Drlica, K.; Zhao, X. DNA gyrase, topoisomerase IV, and the 4-quinolones. Microbiol. Mol. Biol. Rev. 1997, 61, 377–392. [Google Scholar]
- Basu, J.; Chattopadhyay, R.; Kundu, M.; Chakrabarti, P. Purification and partial characterization of a penicillin-binding protein from Mycobacterium smegmatis. J. Bacteriol. 1992, 174, 4829–4832. [Google Scholar] [CrossRef] [Green Version]
- Schweitzer, B.I.; Dicker, A.P.; Bertino, J.R. Dihydrofolate reductase as a therapeutic target. FASEB J. 1990, 4, 2441–2452. [Google Scholar] [CrossRef]
- Aliye, M.; Dekebo, A.; Tesso, H.; Abdo, T.; Eswaramoorthy, R.; Melaku, Y. Molecular docking analysis and evaluation of the antibacterial and antioxidant activities of the constituents of Ocimum cufodontii. Sci. Rep. 2021, 11, 10101. [Google Scholar] [CrossRef]
- da Silva, J.K.R.; Figueiredo, P.L.B.; Byler, K.G.; Setzer, W.N. Essential oils as antiviral agents, potential of essential oils to treat SARS-CoV-2 infection: An in-silico investigation. Int. J. Mol. Sci. 2020, 21, 3426. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | % Area of the Peak | ||
|---|---|---|---|
| EO1 | EO2 | EO3 | |
| 6-Isopropenyl-4,8a-dimethyl-1,2,3,5,6,7,8,8a-octahydro-naphthalen-2-ol | 16.22 | - | - |
| Phenol, 2-methoxy-3-(2-propenyl)- | 14.71 | 0.34 | 8.52 |
| γ-Muurolene | 6.57 | 0.77 | 7.43 |
| 9-Methoxycalamenene | 6.51 | - | - |
| β-Pinene | 4.86 | 0.99 | 1.09 |
| Aromadendrene oxide-(1) | 4.21 | - | - |
| Ylangene | 4.09 | - | - |
| 3-Cyclohexen-1-ol, 4-methyl-1-(1-methylethyl)-, (R)- | 3.01 | - | 0.61 |
| α-Muurolene | 2.90 | 1.48 | - |
| 2,6-Octadienal, 3,7-dimethyl-, (Z)- | 2.78 | - | - |
| Eremophilene | 2.74 | - | - |
| Caryophylene oxide | 2.67 | 0.43 | 0.66 |
| β-trans-Ocimene | 2.50 | - | 1.48 |
| β-Bourbonene | 2.48 | - | - |
| Cyclohexane, 2-ethenyl-1,1-dimethyl-3-methylene- | 2.27 | - | - |
| β-Ocimene | 1.60 | - | - |
| Bicyclo[3.1.0]hexan-2-ol, 2-methyl-5-(1-methylethyl)-, (1α,2α,5α)- | 1.60 | 0.24 | - |
| δ-Cadinene | 1.45 | - | - |
| Benzeneethanamine, α-methyl- | 1.34 | - | - |
| α-Caryophyllene | 1.25 | - | 0.50 |
| αthujene | 1.15 | - | - |
| 2-Methylbicyclo [4.3.0] non-1(6)-ene | 1.09 | - | - |
| 1H-Benzocyclohepten-7-ol, 2,3,4,4a,5,6,7,8-octahydro-1,1,4a,7-tetramethyl-, cis- | 1.09 | - | - |
| Selina-3,7(11)-diene | 1.06 | 4.11 | - |
| β-Caryophyllene epoxide | 0.99 | - | - |
| 2,6-Dimethyl-1,3,5,7-octatetraene, E,E- | 0.91 | 0.72 | - |
| γ-Terpinen | 0.72 | - | 0.82 |
| 3-Cyclohexene-1-methanol, α,α4-trimethyl- | 0.67 | - | - |
| α-Phellandrene | 0.65 | - | 0.36 |
| Naphthalene, 1,2,3,4,6,8a-hexahydro-1-isopropyl-4,7-dimethyl- | 0.61 | 0.24 | - |
| Bicyclo[3.1.0]hex-2-ene, 4,4,6,6-tetramethyl- | 0.58 | - | - |
| Alloaromadendrene oxide-(1) | 0.48 | - | - |
| 3,7-Octadiene-2,6-diol, 2,6-dimethyl- | 0.33 | - | - |
| cis-p-Mentha-2,8-dien-1-ol | 0.28 | - | - |
| Methyl 4,7,10,13-hexadecatetraenoate | 0.26 | - | - |
| β-Cubebene | 0.25 | 4.49 | - |
| Linalool | 0.25 | - | - |
| Tricyclo[5.2.2.0(1,6)]undecan-3-ol, 2-methylene-6,8,8-trimethyl- | 0.24 | - | - |
| Tau-Cadinol | 0.23 | - | - |
| Aromadendrene oxide-(2) | 0.23 | 0.37 | - |
| Aristolene epoxide | 0.22 | - | - |
| Eugenol | 0.22 | - | - |
| β-Phellandrene | 0.18 | 1.39 | 0.22 |
| 1S-α-Pinene | 0.18 | - | - |
| α-Cadinol | 0.18 | 0.38 | - |
| β-Caryophyllene | 0.17 | 1.76 | - |
| α-Guaiene | - | 20.78 | - |
| 2H-1-Benzopyran, 3,4,4a,5,6,8a-hexahydro-2,5,5,8a-tetramethyl-(2α,4aα,8aα)- | - | 12.15 | 4.52 |
| α-Selinene | - | 10.84 | - |
| β-Guaiene | - | 5.03 | - |
| Germacrene D | - | 4.82 | 0.18 |
| 1,2,3,4,6,7,8,8a-Octahydronaphthalene-6,7-diol, 5,8a-dimethyl-3-isopropenyl-, cyclic carbonate, trans- | - | 4.35 | - |
| α-Citral | - | 3.37 | 14.75 |
| Globulol | - | 2.47 | - |
| β-Citral | - | 2.28 | 2.53 |
| β-Selinene | - | 1.65 | 0.24 |
| Camphene | - | 1.34 | 1.53 |
| 1R,4S,7S,11R-2,2,4,8-Tetramethyltricyclo[5.3.1.0(4,11)]undec-8-ene | - | 1.13 | - |
| o-Menth-8-ene, 4-isopropylidene-1-vinyl- | - | 1.00 | 0.70 |
| (−)-β-Elemene | - | 0.95 | - |
| Borneol | - | 0.92 | 0.21 |
| Cyclohexene, 1-methyl-4-(1-methylethylidene)- | - | 0.80 | - |
| Cadina-1(10),4-diene | - | 0.71 | - |
| Isocaryophillene | - | 0.70 | - |
| α-Humulene | - | 0.62 | 2.74 |
| γ-Cadinene | - | 0.61 | - |
| Androstan-17-one, 3-ethyl-3-hydroxy-, (5α)- | - | 0.60 | - |
| Isoaromadendrene epoxide | - | 0.60 | - |
| 1R-α-Pinene | - | 0.52 | 3.11 |
| n-Amyl isovalerate | - | 0.50 | - |
| (+)-Cyclosativene | - | 0.46 | - |
| Terpinolene | - | 0.45 | 0.88 |
| (+)-Sativene | - | 0.45 | - |
| Bicyclo[2.2.1]heptan-2-one, 1,7,7-trimethyl-, (1S)- | - | 0.44 | - |
| Benzene, 4-allyl-1,2-dimethoxy- | - | 0.40 | - |
| D-Limonene | - | 0.31 | 1.15 |
| Geranyl vinyl ether | - | 0.27 | - |
| 7(11)-Selinen-4α-ol | - | 0.27 | 0.74 |
| 1,5-Dimethyl-1-vinyl-4-hexenyl butyrate | - | 0.27 | - |
| Cubenol | - | 0.26 | - |
| Cyclohexane, 1-ethenyl-1-methyl-2,4-bis(1-methylethenyl)-, [1S-(1α,2β,4β)]- | - | - | 11.81 |
| Caryophyllene | - | - | 8.91 |
| α-Amorphene | - | - | 7.78 |
| 2-Isopropenyl-4a,8-dimethyl-1,2,3,4,4a,5,6,7-octahydronaphthalene | - | - | 3.44 |
| Juniper camphor | - | - | 2.95 |
| β-Elemene | - | - | 2.26 |
| Isoborneol | - | - | 1.54 |
| γ-Gurjunene | - | - | 1.14 |
| α-Longipinene | - | - | 1.04 |
| Guaia-1(10),11-diene | - | - | 0.94 |
| Epiglobulol | - | - | 0.90 |
| α-Terpinen | - | - | 0.68 |
| α-Panasinsen | - | - | 0.51 |
| Furan, 2,5-diethyltetrahydro- | - | - | 0.30 |
| Selina-6-en-4-ol | - | - | 0.29 |
| 4(10)-Thujene | - | - | 0.20 |
| Octanal | - | - | 0.17 |
| Eucalyptol | - | - | 0.16 |
| Minimum Inhibitory Concentration (v/v) | |||
|---|---|---|---|
| Staphylococcus aureus | Escherichia coli | Enterobacter cloacae | |
| EO1 | 12.5 | 6.25 | 25 |
| EO2 | 25 | 25 | 25 |
| EO3 | 12.5 | 12.5 | 25 |
| Compounds | DNA-Gyrase | Topoisomerase-4 | Dihydrofolate Reductase | Penicillin Binding Protein | Dihydropteroate Synthase |
|---|---|---|---|---|---|
| Binding Energy (kcal mol−1) | |||||
| Eucalyptol | −4.6 | −5.3 | −5.9 | −5.8 | −4.8 |
| Eugenol | −5.1 | −5.4 | −6.0 | −4.6 | −4.8 |
| Linalool | −4.9 | −4.7 | −5.4 | −4.3 | −4.4 |
| γ-terpinene | −5.8 | −5.3 | −5.6 | −5.6 | −4.7 |
| β-Phellandrene | −5.7 | −5.2 | −5.7 | −5.7 | −5.2 |
| β-Pinene | −4.7 | −5.1 | −5.7 | −5.7 | −5.3 |
| β-ocimene | −5.3 | −4.7 | −5.4 | −4.7 | −4.3 |
| Borneol | −4.6 | −5.4 | −5.4 | −4.6 | −5.0 |
| 1R-α-pinene | −4.6 | −5.9 | −5.7 | −5.0 | −4.5 |
| β-cubebene | −6.7 | −6.7 | −7.6 | −6.5 | −6.1 |
| α-citral | −4.7 | −4.0 | −5.1 | −4.7 | −4.4 |
| caryophyllene oxide | −5.6 | −6.7 | −7.8 | −6.5 | −5.8 |
| Ylangene | −6.7 | −6.7 | −7.2 | −6.5 | −6.0 |
| Ciprofloxacin | −7.1 | −7.4 | - | - | - |
| Trimethoprim | - | - | −7.5 | - | - |
| Sulfamethoxazole | - | - | - | - | −6.2 |
| Cephalosporin | - | - | - | −6.1 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salvi, P.; Kumar, G.; Gandass, N.; Kajal; Verma, A.; Rajarammohan, S.; Rai, N.; Gautam, V. Antimicrobial Potential of Essential Oils from Aromatic Plant Ocimum sp.; A Comparative Biochemical Profiling and In-Silico Analysis. Agronomy 2022, 12, 627. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030627
Salvi P, Kumar G, Gandass N, Kajal, Verma A, Rajarammohan S, Rai N, Gautam V. Antimicrobial Potential of Essential Oils from Aromatic Plant Ocimum sp.; A Comparative Biochemical Profiling and In-Silico Analysis. Agronomy. 2022; 12(3):627. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030627
Chicago/Turabian StyleSalvi, Prafull, Gulshan Kumar, Nishu Gandass, Kajal, Ashish Verma, Sivasubramanian Rajarammohan, Nilesh Rai, and Vibhav Gautam. 2022. "Antimicrobial Potential of Essential Oils from Aromatic Plant Ocimum sp.; A Comparative Biochemical Profiling and In-Silico Analysis" Agronomy 12, no. 3: 627. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030627