
Expression of the VvMYB60 Transcription Factor Is Restricted to Guard Cells and Correlates with the Stomatal Conductance of the Grape Leaf

 , , , , ,
, , , , ,  and
and 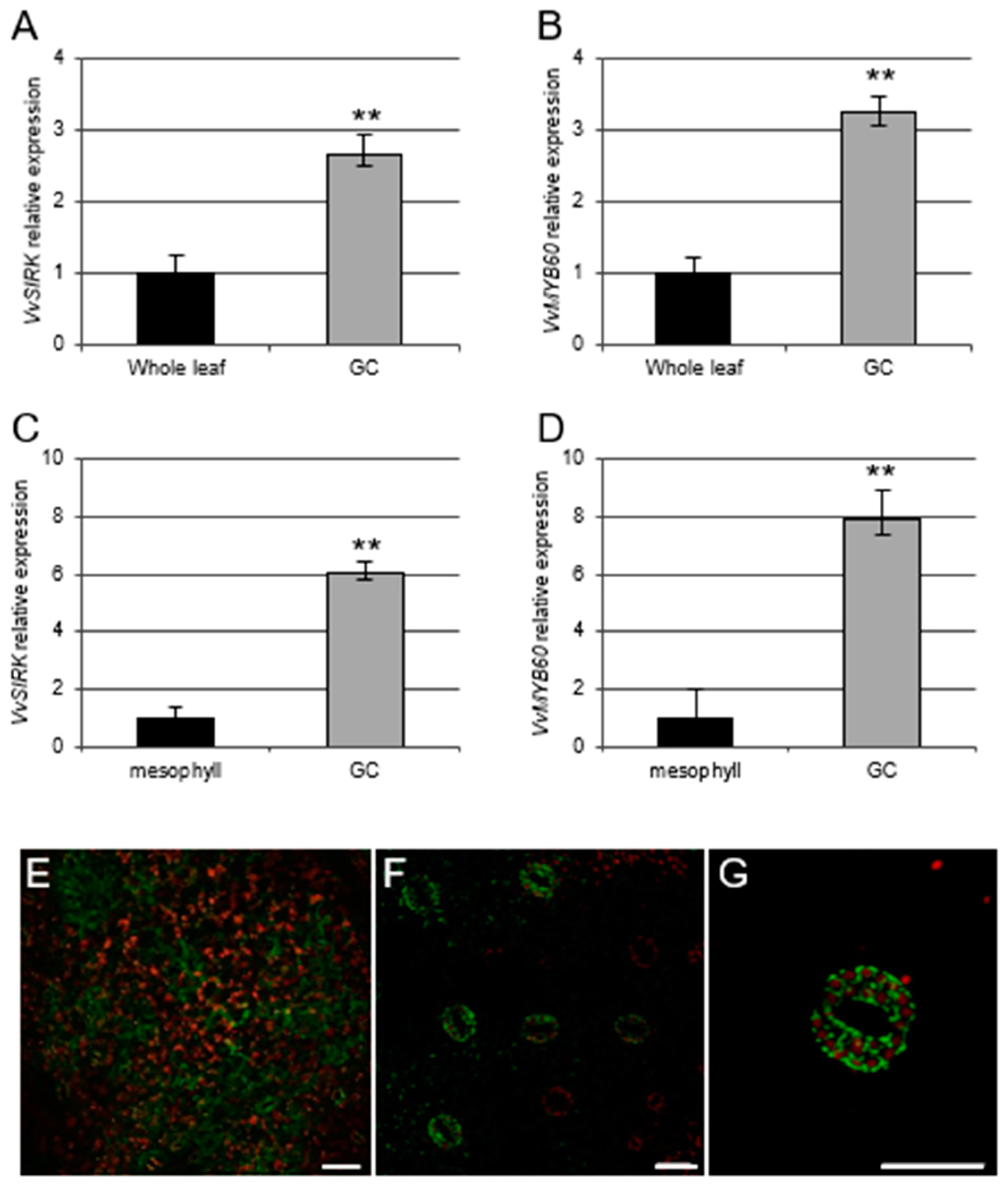{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Condition, Leaf Sampling, and Stomatal Measurements
2.2. Stomatal Conductivity (gs) and Stomatal Density Measurements
2.3. Ice Blending and Laser Microdissection of Grape Leaves
2.4. qPCR Analysis
2.5. Plasmid Constructs, Leaves Infiltration and Confocal Analysis
3. Results and Discussion
3.1. Analysis of the Cellular Specificity of VvMYB60 Expression in the Grape Leaf
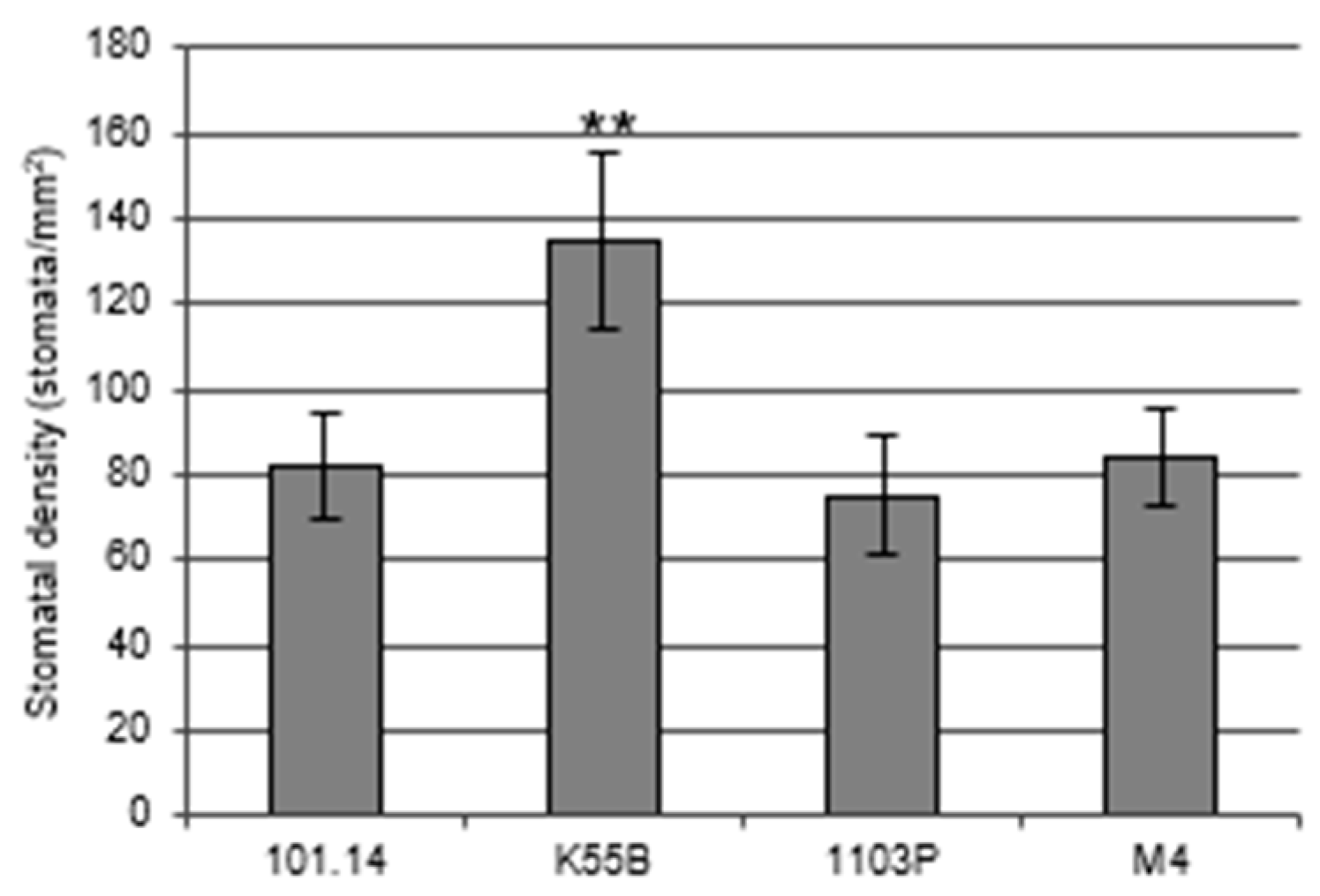
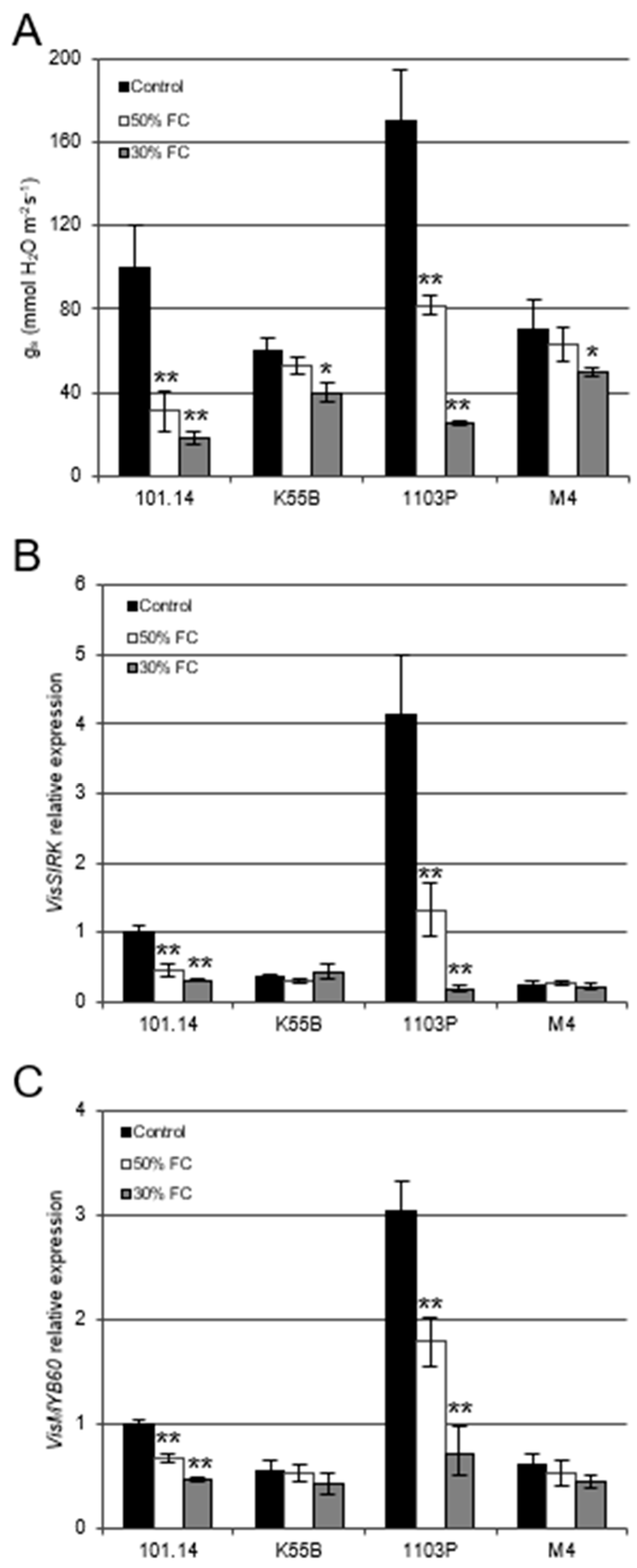
3.2. Different Rootstocks Disclose a Positive Correlation between VisMYB60 Expression and Stomatal Conductance
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Chaves, M.; Zarrouk, O.; Francisco, R.; Costa, J.; Santos, T.; Regalado, A.; Rodrigues, M.; Lopes, C. Grapevine under deficit irrigation: Hints from physiological and molecular data. Annal. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.; Chaves, M.; Wendler, R.; David, M.; Quick, P.; Leegood, R.; Stitt, M.; Pereira, J. Osmotic adjustment in water stressed grapevine leaves in relation to carbon assimilation. Austr. J. Plant Physiol. 1993, 20, 309–321. [Google Scholar] [CrossRef]
- Chaves, M.M.; Oliveira, M.M. Mechanisms underlying plant resilience to water deficits: Prospects for water-saving agriculture. J. Exp. Bot. 2004, 55, 2365–2384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tardieu, F.; Simonneau, T. Variability among species of stomatal control under fluctuating soil water status and evaporative demand: Modelling isohydric and anisohydric behaviours. J. Exp. Bot. 1998, 46, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Hochberg, U.; Windt, C.W.; Ponomarenko, A.; Zhang, Y.J.; Gersony, J.; Rockwell, F.; Holbrook, N. Stomatal closure, basal leaf embolism and shedding protect the hydraulic integrity of grape stems. Plant Physiol. 2017, 174, 764–775. [Google Scholar] [CrossRef] [Green Version]
- Guillier, C.; Gamm, M.; Lucchi, G.; Truntzer, C.; Pecqueur, D.; Ducoroy, P.; Adrian, M.; Héloir, M.C. Toward the identification of two glycoproteins involved in the stomatal deregulation of downy mildew–infected grapevine leaves. Mol. Plant Microbe Interact. 2015, 28, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- MacRobbie, E.A. Signal transduction and ion channels in guard cells. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1998, 353, 1475–1488. [Google Scholar] [CrossRef] [PubMed]
- Sirichandra, C.; Wasilewska, A.; Vlad, F.; Valon, C.; Leung, J. The guard cell as a single-cell model towards understanding drought tolerance and abscisic acid action. J. Exp. Bot. 2009, 60, 1439–1463. [Google Scholar] [CrossRef] [Green Version]
- Cominelli, E.; Galbiati, M.; Tonelli, C. Transcription factors controlling stomatal movements and drought tolerance. Transcription 2010, 1, 41–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matus, J.; Aquea, F.; Arce-Johnson, P. Analysis of the grape MYB R2R3 subfamily reveals expanded wine quality-related clades and conserved gene structure organization across Vitis and Arabidopsis genomes. BMC Plant Biol. 2008, 8, 83. [Google Scholar] [CrossRef] [Green Version]
- Czemmel, S.; Heppel, S.; Bogs, J. R2R3 MYB transcription factors: Key regulators of the flavonoid biosynthetic pathway in grapevine. Protoplasma 2012, 249, S109–S118. [Google Scholar] [CrossRef] [PubMed]
- Galbiati, M.; Matus, J.T.; Francia, P.; Rusconi, F.; Canon, P.; Medina, C.; Conti, L.; Cominelli, E.; Tonelli, C.; Arce-Johnson, P. The grapevine guard cell-related VvMYB60 transcription factor is involved in the regulation of stomatal activity and is differentially expressed in response to ABA and osmotic stress. BMC Plant Biol. 2011, 11, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cominelli, E.; Galbiati, M.; Vavasseur, A.; Conti, L.; Sala, T.; Vuylsteke, M.; Leonhardt, N.; Dellaporta, S.L.; Tonelli, C. A guard-cell-specific MYB transcription factor regulates stomatal movements and plant drought tolerance. Curr. Biol. 2005, 15, 1196–1200. [Google Scholar] [CrossRef] [PubMed]
- Simeoni, F.; Skirycz, A.; Simoni, L.; Castorina, G.; Perez de Souza, L.; Fernie, A.R.; Alseekh, S.; Giavalisco, P.; Conti, L.; Tonelli, C.; et al. The AtMYB60 transcription factor regulates stomatal opening by modulating oxylipin synthesis in guard cells. Sci. Rep. 2022, 12, 533. [Google Scholar] [CrossRef] [PubMed]
- Meggio, F.; Prinsi, B.; Negri, A.S.; Dilorenzo, G.S.; Lucchini, G.; Pitacco, A.; Failla, O.; Scienza, A.; Cocucci, M.; Espen, L. Biochemical and physiological responses of two grapevine rootstock genotypes to drought and salt treatments. Austr. J. Grape Wine Res. 2014, 20, 310–323. [Google Scholar] [CrossRef]
- Bauer, H.; Ache, P.; Lautner, S.; Fromm, J.; Hartung, W.; Al-Rasheid, K.A.; Sonnewald, S.; Sonnewald, U.; Kneitz, S.; Lachmann, N.; et al. The stomatal response to reduced relative humidity requires guard cell-autonomous ABA synthesis. Curr. Biol. 2013, 23, 53–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galbiati, M.; Simoni, L.; Pavesi, G.; Cominelli, E.; Francia, P.; Vavasseur, A.; Nelson, T.; Bevan, M.; Tonelli, C. Gene trap lines identify Arabidopsis genes expressed in stomatal guard cells. Plant J. 2008, 53, 750–762. [Google Scholar] [CrossRef]
- Matus, J.; Loyola, R.; Vega, A.; Pena-Neira, A.; Bordeu, E.; Arce-Johnson, P. Post-veraison sunlight exposure induces MYB-mediated transcriptional regulation of anthocyanin and flavonol synthesis in berry skins of Vitis vinifera. J. Exp. Bot. 2009, 60, 853–867. [Google Scholar] [CrossRef] [Green Version]
- Davis, S.; Viestra, R. Soluble, highly fluorescent variants of green fluorescent protein (GFP) for use in higher plants. Plant Mol. Biol. 1998, 36, 521–528. [Google Scholar] [CrossRef]
- Zottini, M.; Barizza, E.; Costa, A.; Formentin, E.; Ruberti, C.; Carimi, F.; Lo Schiavo, F. Agroinfiltration of grapevine leaves for fast transient assays of gene expression and for long-term production of stable transformed cells. Plant Cell Rep. 2008, 27, 845–853. [Google Scholar] [CrossRef]
- Pratelli, R.; Lacombe, B.; Torregrosa, L.; Gaymard, F.; Romieu, C.; Thibaud, J.; Sentenac, H. A grapevine gene encoding a guard cell K(+) channel displays developmental regulation in the grapevine berry. Plant Physiol. 2002, 128, 564–577. [Google Scholar] [CrossRef]
- Kerk, N.M.; Ceserani, T.; Tausta, S.; Sussex, I.; Nelson, T. Laser Capture Microdissection of cells from plant tissues. Plant Physiol. 2003, 132, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Carbonneau, A. The early selection of grapevine rootstocks for resistance to drought conditions. Am. J. Enol. Viticol. 1985, 36, 195–198. [Google Scholar]
- Pavlusek, P. Evaluation of drought tolerance of new grapevine rootstock hybrids. J. Environ. Biol. 2011, 32, 543–549. [Google Scholar]
- Dal Santo, S.; Palliotti, A.; Zenoni, S.; Tornielli, G.B.; Fasoli, M.; Paci, P.; Tombesi, S.; Frioni, T.; Silvestroni, O.; Bellincontro, A.; et al. Distinct transcriptome responses to water limitation in isohydric and anisohydric grapevine cultivars. BMC Genom. 2016, 17, 815. [Google Scholar] [CrossRef] [Green Version]
- Prinsi, B.; Simeoni, F.; Galbiati, M.; Meggio, F.; Tonelli, C.; Scienza, A.; Espen, L. Grapevine Rootstocks Differently Affect Physiological and Molecular Responses of the Scion under Water Deficit Condition. Agronomy 2021, 11, 289. [Google Scholar] [CrossRef]
- Alsina, M.; Smart, D.; Bauerle, T.; Herralde, F.D.; Biel, C.; Stockert, C.; Negron, C.; Save, R. Seasonal changes of whole root system conductance by a drought-tolerant grape root system. J. Exp. Bot. 2011, 62, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Rusconi, F.; Simeoni, F.; Francia, P.; Cominelli, E.; Conti, L.; Riboni, M.; Simoni, L.; Martin, C.R.; Tonelli, C.; Galbiati, M. The Arabidopsis thaliana MYB60 promoter provides a tool for the spatio-temporal control of gene expression in stomatal guard cells. J. Exp. Bot. 2013, 64, 3361–3371. [Google Scholar] [CrossRef]
- de la Guardia, A.R.-H.; Ugalde, M.B.; Lobos-Diaz, V.; Romero-Romero, J.L.; Meyer-Regueiro, C.; Inostroza-Blancheteau, C.; Reyes-Diaz, M.; Aquea, F.; Arce-Johnson, P. Isolation and molecular characterization of MYB60 in Solanum lycopersicum. Mol. Biol. Rep. 2021, 48, 1579–1587. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simeoni, F.; Simoni, L.; Zottini, M.; Conti, L.; Tonelli, C.; Castorina, G.; Espen, L.; Galbiati, M. Expression of the VvMYB60 Transcription Factor Is Restricted to Guard Cells and Correlates with the Stomatal Conductance of the Grape Leaf. Agronomy 2022, 12, 694. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030694
Simeoni F, Simoni L, Zottini M, Conti L, Tonelli C, Castorina G, Espen L, Galbiati M. Expression of the VvMYB60 Transcription Factor Is Restricted to Guard Cells and Correlates with the Stomatal Conductance of the Grape Leaf. Agronomy. 2022; 12(3):694. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030694
Chicago/Turabian StyleSimeoni, Fabio, Laura Simoni, Michela Zottini, Lucio Conti, Chiara Tonelli, Giulia Castorina, Luca Espen, and Massimo Galbiati. 2022. "Expression of the VvMYB60 Transcription Factor Is Restricted to Guard Cells and Correlates with the Stomatal Conductance of the Grape Leaf" Agronomy 12, no. 3: 694. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12030694