Red and Blue LED Light Supplementation in the Morning Pre-activates the Photosynthetic System of Tomato (Solanum lycopersicum L.) Leaves and Promotes Plant Growth

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Experimental Design
2.3. Growth Index Determination
2.4. Dry Matter Accumulation, Specific Leaf Weight, and Root Parameters
2.5. Pigment Content and Gas Exchange Parameters
2.6. Determination of Ribulose 1, 5-Bisphosphate Carboxylase (Rubisco) Enzyme Activity
2.7. Determination of Photosynthetic Products
2.8. Statistical Analyses
3. Results
3.1. Growth and Morphology
3.2. Dry Matter Accumulation
3.3. Root Parameters and Specific Leaf Weight
3.4. Photosynthetic Pigments
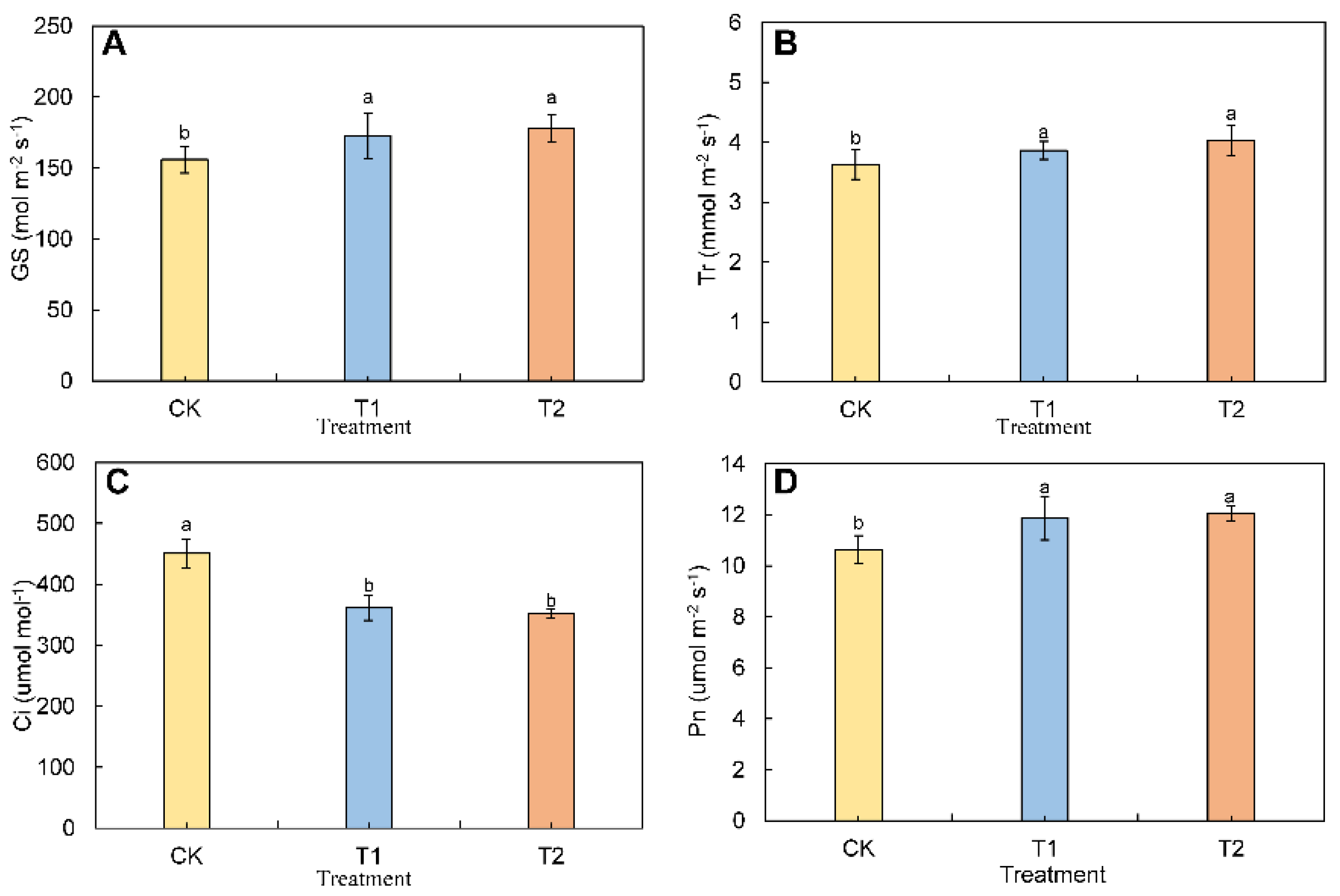
3.5. Gas Exchange Parameters
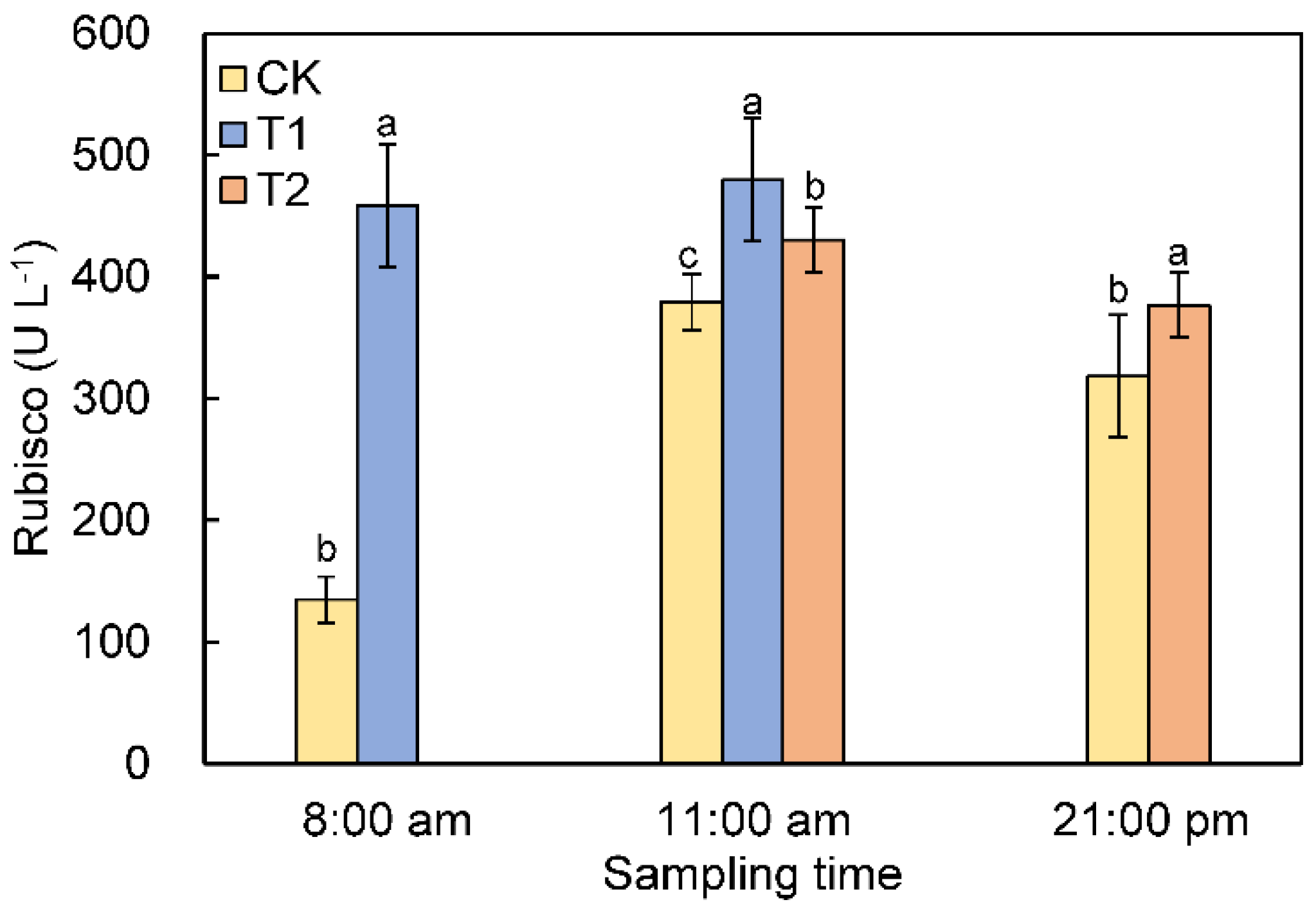
3.6. Rubisco Activity
3.7. Photosynthates
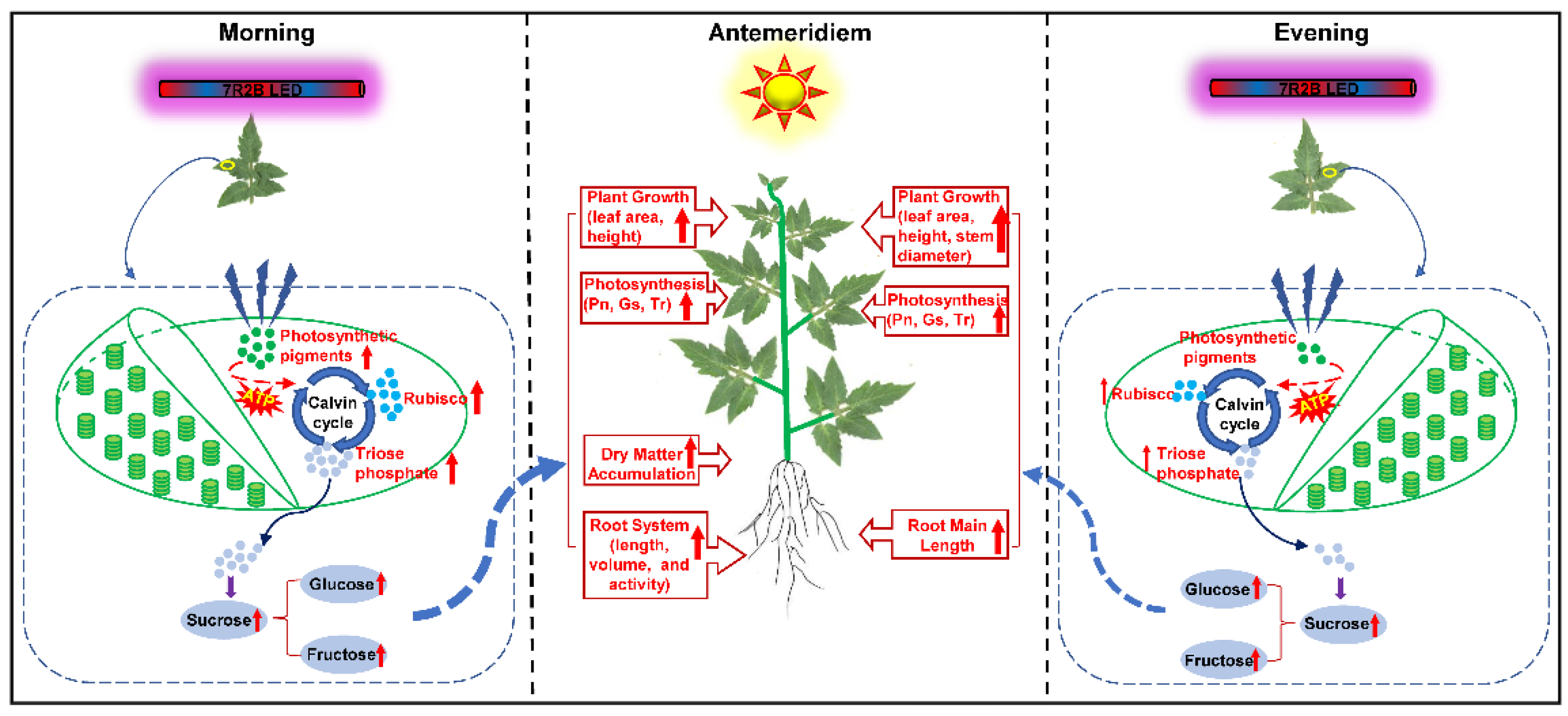
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liao, X.R.; Zhang, L.; Xu, J.Z.; Du, J.F. The role of light in plant growth and development. J. Hebei Univ. (Nat. Sci. Ed.) 2001, 21, 341–346. (In Chinese) [Google Scholar]
- Izzo, L.G.; Hay Mele, B.; Vitale, L.; Vitale, E.; Arena, C. The role of monochromatic red and blue light in tomato early photomorphogenesis and photosynthetic traits. Environ. Exp. Bot. 2020, 179, 104195. [Google Scholar] [CrossRef]
- Dorais, M.; Ehret, D.L.; Papadopoulos, A.P. Tomato (Solanum Lycopersicum) health components: From the seed to the consumer. Phytochem. Rev. 2008, 7, 231–250. [Google Scholar] [CrossRef]
- Elings, A.; Meinen, E.; Dieleman, J.A.; Visser, P.H.B.D. The modelled photosynthetic effects of different light colours on tomato crop growth and production. Acta Hortic. 2017, 1182, 177–184. [Google Scholar] [CrossRef]
- Blom, T.J.; Ingratta, F.J. The effect of high pressure sodium lighting on the production of tomatoes, cucumbers, and roses. Acta Hortic. 1984, 148, 905–914. [Google Scholar] [CrossRef]
- Frantz, J.M.; Joly, R.J.; Mitchell, C.A. Intracanopy lighting influences radiation capture, productivity, and leaf senescence in cowpea canopies. J. Am. Soc. Hortic. Sci. 2000, 125, 694–701. [Google Scholar] [CrossRef] [Green Version]
- Steinger, T.; Roy, B.A.; Stanton, M.L. Evolution in stressful environments II: Adaptive value and costs of plasticity in response to low light in Sinapis arvensis. J. Evol. Biol. 2003, 16, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, R.M. A historical background of plant lighting: An introduction to the workshop. Hortscience 2008, 43, 1942–1943. [Google Scholar] [CrossRef] [Green Version]
- Samuolienė, G.; Sirtautas, R.; Brazaitytė, A.; Duchovskis, P. LED lighting and seasonality effects antioxidant properties of baby leaf lettuce. Food Chem. 2012, 134, 1494–1499. [Google Scholar] [CrossRef]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review. J. Sci. Food Agric. 2015, 95, 869–877. [Google Scholar] [CrossRef]
- Ouzounis, T.; Fretté, X.; Ottosen, C.O.; Rosenqvist, E. Spectral effects of LEDs on chlorophyll fluorescence and pigmentation in Phalaenopsis ‘Vivien’ and ‘Purple Star’. Physiol. Plant. 2015, 154, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Dayani, S.; Heydarizadeh, P.; Sabzalian, M.R. Efficiency of Light Emitting Diodes (LEDs) for the Future Photosynthesis. In Handbook of Photosynthesis; CRC Press: Boca Raton, FL, USA, 2017; pp. 761–783. [Google Scholar]
- Nelson, J.A.; Bruce, B.; Andrew, C.D. Economic Analysis of Greenhouse Lighting: Light Emitting Diodes vs. high Intensity Discharge Fixtures. PLoS ONE 2014, 9, e99010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.D.; Liu, X.Y.; Jiao, X.L.; Xu, Z.G. Effect of blue light quantity on growth and quality of lettuce. J. Nanjing Agric. Univ. 2015, 38, 890–895. (In Chinese) [Google Scholar]
- Cosgrove, D.J. Rapid inhibition of hypocotyl growth by blue light in Sinapis alba L. Plant Sci. Lett. 1982, 25, 305–312. [Google Scholar] [CrossRef]
- Baroli, I.; Price, G.D.; Badger, M.R.; Caemmerer, S.V. The contribution of photosynthesis to the red light response of stomatal conductance. Plant Physiol. 2008, 146, 737–747. [Google Scholar] [CrossRef] [Green Version]
- Naznin, M.T.; Lefsrud, M.; Azad, M.O.K.; Park, C.H. Effect of different combinations of red and blue LED light on growth characteristics and pigment content of in vitro tomato plantlets. Agriculture 2019, 9, 196. [Google Scholar] [CrossRef] [Green Version]
- Paponov, M.; Kechasov, D.; Lacek, J.; Verheul, M.J.; Paponov, I.A. Supplemental light-emitting diode inter-lighting increases tomato fruit growth through enhanced photosynthetic light use efficiency and modulated root activity. Front. Plant Sci. 2019, 10, 1656. [Google Scholar] [CrossRef]
- Chen, X.L.; Wang, L.C.; Li, T.; Yang, Q.C.; Guo, W.Z. Sugar accumulation and growth of lettuce exposed to different lighting modes of red and blue LED light. Sci. Rep. 2019, 9, 6926. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.S. The Effect of LED Supplemental Light on the Growth and Quality of Cucumber, Tomato and Strawberry in Solar Greenhouse. Master Thesis, Hebei University of Engineering, Handan, Hebei, China, 2018. (In Chinese). [Google Scholar]
- Kaiser, E.; Ouzounis, T.; Giday, H.; Schipper, R.; Heuvelink, E.; Marcelis, L.F.M. Adding blue to red supplemental light increases biomass and yield of greenhouse-grown tomatoes, but only to an optimum. Front. Plant Sci. 2018, 9, 2002. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xin, G.F.; Wei, M.; Shi, Q.H.; Yang, F.J.; Wang, X.F. Carbohydrate accumulation and sucrose metabolism responses in tomato seedling leaves when subjected to different light qualities. Sci. Hortic. 2017, 225, 490–497. [Google Scholar] [CrossRef]
- Wang, S.Y.; Lv, J.; Yu, J.H.; Jin, N.; Jin, L.; Liu, X.Q.; Wei, S.H.; Gao, F. The effect of different light supplement time on the growth, yield and quality of tomato in solar greenhouse. Chin. Veg. 2018, 356, 41–45. (In Chinese) [Google Scholar]
- Wang, S.Y.; Fang, H.; Xie, J.M.; Wu, Y.; Tang, Z.Q.; Liu, Z.C.; Lv, J.; Yu, J.H. Physiological responses of cucumber seedlings to different supplemental light duration of red and blue LED. Front. Plant Sci. 2021, 12, 709313. [Google Scholar] [CrossRef] [PubMed]
- Müller, N.A.; Wijnen, C.L.; Srinivasan, A.; Ryngajllo, M.; Ofner, I.; Lin, T.; Ranjan, A.; West, D.; Maloof, J.N.; Sinha, N.R.; et al. Domestication selected for deceleration of the circadian clock in cultivated tomato. Nat. Genet. 2016, 48, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.X.; Li, D.X.; Li, Y.L. Study on method to measure tomato leaf area. J. Northwest A&F Univ. (Nat. Sci. Ed.) 2006, 34, 116–120. [Google Scholar] [CrossRef]
- Li, H.S. Principles and Techniques of Plant Physiological Experiment; Higher Education Press: Beijing, China, 2000; pp. 119–120. (In Chinese) [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenol oxidase in beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.N.; Xie, J.M.; Lv, J.; Li, J.; Zhang, J.; Wang, C.; Liang, G.P. Alleviating damage of photosystem and oxidative stress from chilling stress with exogenous zeaxanthin in pepper (Capsicum annuum L.) seedlings. Plant Physiol. Biochem. 2021, 162, 395–409. [Google Scholar] [CrossRef]
- Tewolde, F.T.; Lu, N.; Shiina, K.; Maruo, T.; Takagaki, M.; Kozai, T.; Yamori, W. Nighttime supplemental LED inter-lighting improves growth and yield of single-truss tomatoes by enhancing photosynthesis in both winter and summer. Front. Plant Sci. 2016, 7, 448. [Google Scholar] [CrossRef]
- Treder, J.; Borkowska, A.; Treder, W.; Klamkowski, K. The effects of LEDs on growth and morphogenesis of vegetable seedlings cultivated in growth chambers. In Proceedings of the IEEE Lighting Conference of the Visegrad Countries (Lumen V4), Karpacz, Poland, 13–16 September 2016; pp. 1–4. [Google Scholar] [CrossRef]
- Wu, M.C.; Hou, C.Y.; Jiang, C.M.; Wang, Y.T.; Wang, C.Y.; Chen, H.H.; Chang, H.M. A novel approach of LED light radiation improves the antioxidant activity of pea seedlings. Food Chem. 2007, 101, 1753–1758. [Google Scholar] [CrossRef]
- Klamkowski, K.; Treder, W.; Wojcik, K.; Puternicki, A.; Lisak, E. Influence of Supplementary Lighting on Growth and Photosynthetic Activity of Tomato Transplants. Infrastruct. Ecol. Rural Areas 2014, 3, 1377–1385. [Google Scholar] [CrossRef]
- Zou, J.; Zhou, C.B.; Xu, H.; Cheng, R.F.; Yang, Q.C.; Li, T. The effect of artificial solar spectrum on growth of cucumber and lettuce under controlled environment. J. Integr. Agric. 2020, 19, 2027–2034. [Google Scholar] [CrossRef]
- Yan, Z.N.; He, D.X.; Niu, G.H.; Zhou, Q.; Qu, Y.H. Growth, nutritional quality, and energy use efficiency in two lettuce cultivars as influenced by white plus red versus red plus blue LEDs. Int. J. Agric. Biol. Eng. 2020, 13, 33–40. [Google Scholar] [CrossRef]
- Liu, X.Y.; Jiao, X.L.; Chang, T.T.; Guo, S.R.; Xu, Z.G. Photosynthesis and leaf development of cherry tomato seedlings under different LED-based blue and red photon flux ratios. Photosynthetica 2018, 56, 1212–1217. [Google Scholar] [CrossRef]
- Ren, X.X.; Liu, Y.; Jeong, H.K.; Jeong, B.R. Supplementary light source affects the growth and development of Codonopsis lanceolata seedlings. Int. J. Mol. Sci. 2018, 19, 3074. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.; Yang, C. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Senger, H.; Briggs, W.R. The blue light receptor (s): Primary reactions and subsequent metabolic changes. In Photochemical and Photobiological Reviews; Springer: Boston, MA, USA, 1981; pp. 1–38. [Google Scholar]
- Zeiger, E. The biology of stomatal guard cells. Annu. Rev. Plant Physiol. 1983, 34, 441–474. [Google Scholar] [CrossRef]
- Kasajima, S.Y.; Inoue, N.; Mahmud, R.; Kato, M. Developmental Responses of Wheat cv. Norin. 61 to fluence rate of green light (crop physiology & ecology). Plant Prod. Sci. 2008, 11, 76–81. [Google Scholar] [CrossRef]
- Wu, Q.; Su, N.N.; Shen, W.B.; Cui, J. Analyzing photosynthetic activity and growth of Solanum Lycopersicum seedlings exposed to different light qualities. Acta Physiol. Plant. 2014, 36, 1411–1420. [Google Scholar] [CrossRef]
- Palmitessa, O.D.; Prinzenberg, A.E.; Kaiser, E.; Heuvelink, E. Led and hps supplementary light differentially affect gas exchange in tomato leaves. Plants 2021, 10, 810. [Google Scholar] [CrossRef]
- Song, Y.; Jiang, C.Y.; Gao, L.H. Polychromatic supplemental lighting from underneath canopy is more effective to enhance tomato plant development by improving leaf photosynthesis and stomatal regulation. Front. Plant Sci. 2016, 7, 1832. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.B.; Zhang, X.; Liu, B.B.; Ai, X.Z.; Bi, G.H.; Li, Q.M. Effects of supplemental light quality on the photosynthetic characteristics of leaf lettuce. Plant Physiol. J. 2015, 51, 2255–2262. [Google Scholar] [CrossRef]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130243. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Zhao, J.; Hu, J.T.; Jeong, B.R. Effect of supplementary light intensity on quality of grafted tomato seedlings and expression of two photosynthetic genes and proteins. Agronomy 2019, 9, 339. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Hu, M.J.; Guo, Y.P. Regulation of photosynthesis by light quality and its mechanism in plants. Chin. J. Appl. Ecol. 2008, 19, 1619–1624. (In Chinese) [Google Scholar]
- Fang, L.Z.; Ma, Z.Y.; Wang, Q.B.; Nian, H.; Ma, Q.B.; Huang, Q.L.; Mu, Y.H. Plant growth and photosynthetic characteristics of soybean seedlings under different LED lighting quality conditions. J. Plant Growth Regul. 2020, 40, 6451. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Photosynthetic Pigment Content (mg·g–1·FW–1) | Chlorophyll a/b | |||
|---|---|---|---|---|---|
| Chlorophyll a | Chlorophyll b | Total Chlorophyll | Carotenoid | ||
| CK | 1.54 ± 0.118b | 0.40 ± 0.019a | 1.94 ± 0.110b | 0.72 ± 0.051b | 3.68 ± 0.243b |
| T1 | 1.81 ± 0.296a | 0.43 ± 0.042a | 2.23 ± 0.362a | 0.82 ± 0.055a | 4.24 ± 0.351a |
| T2 | 1.61 ± 0.113ab | 0.44 ± 0.036a | 2.06 ± 0.156ab | 0.78 ± 0.027ab | 3.66 ± 0.205b |
| Sampling Time | Treatment | Photosynthate Content (mg·g–1) | ||||
|---|---|---|---|---|---|---|
| Glucose | Fructose | Sucrose | Starch | Total Soluble Sugar | ||
| 8:00 am | CK | 14.23 ± 1.62b | 6.63 ± 0.79b | 11.35 ± 1.67b | 7.1 ± 3.4a | 46.15 ± 8.21b |
| T1 | 17.13 ± 0.98a | 11.95 ± 1.43a | 22.71 ± 1.35a | 4.4 ± 1.2b | 98.23 ± 4.93a | |
| 11:00 am | CK | 3.05 ± 0.57c | 3.94 ± 1.07c | 6.64 ± 0.61b | 18.1 ± 2.1a | 56.12 ± 3.11b |
| T1 | 7.42 ± 0.23a | 6.43 ± 0.39b | 7.03 ± 0.94b | 8.1 ± 1.6b | 109.21 ± 11.25a | |
| T2 | 6.79 ± 0.19b | 8.18 ± 0.61a | 12.71 ± 1.38a | 2.4 ± 1.2c | 95.86 ± 10.43a | |
| 21:00 pm | CK | 3.38 ± 1.02b | 3.86 ± 0.42b | 6.61 ± 1.09b | 11.6 ± 2.1a | 40.94 ± 2.44b |
| T2 | 9.89 ± 0.96a | 5.67 ± 0.73a | 9.02 ± 0.46a | 5.1 ± 1.0b | 92.89 ± 7.76a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Meng, X.; Tang, Z.; Wu, Y.; Xiao, X.; Zhang, G.; Hu, L.; Liu, Z.; Lyu, J.; Yu, J. Red and Blue LED Light Supplementation in the Morning Pre-activates the Photosynthetic System of Tomato (Solanum lycopersicum L.) Leaves and Promotes Plant Growth. Agronomy 2022, 12, 897. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12040897
Wang S, Meng X, Tang Z, Wu Y, Xiao X, Zhang G, Hu L, Liu Z, Lyu J, Yu J. Red and Blue LED Light Supplementation in the Morning Pre-activates the Photosynthetic System of Tomato (Solanum lycopersicum L.) Leaves and Promotes Plant Growth. Agronomy. 2022; 12(4):897. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12040897
Chicago/Turabian StyleWang, Shuya, Xin Meng, Zhongqi Tang, Yue Wu, Xuemei Xiao, Guobin Zhang, Linli Hu, Zeci Liu, Jian Lyu, and Jihua Yu. 2022. "Red and Blue LED Light Supplementation in the Morning Pre-activates the Photosynthetic System of Tomato (Solanum lycopersicum L.) Leaves and Promotes Plant Growth" Agronomy 12, no. 4: 897. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12040897