
Defining Suitable Reference Genes for qRT-PCR in Plagiodera versicolora (Coleoptera: Chrysomelidae) under Different Biotic or Abiotic Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Experimental Treatments
2.3. Development Stage and Sex
2.3.1. Tissue
2.3.2. Thermal Exposure
2.3.3. Pathogen Treatment
2.3.4. dsRNA Treatment
2.4. RNA Extraction and cDNA Preparation
2.5. Candidate Reference Genes and Primer Design
2.6. qRT-PCR Assay
2.7. Stability Analysis of Candidate Reference Genes
2.8. Validation of Reference Genes
3. Results
3.1. Evaluation of Primer Specificity and Amplification Efficiency
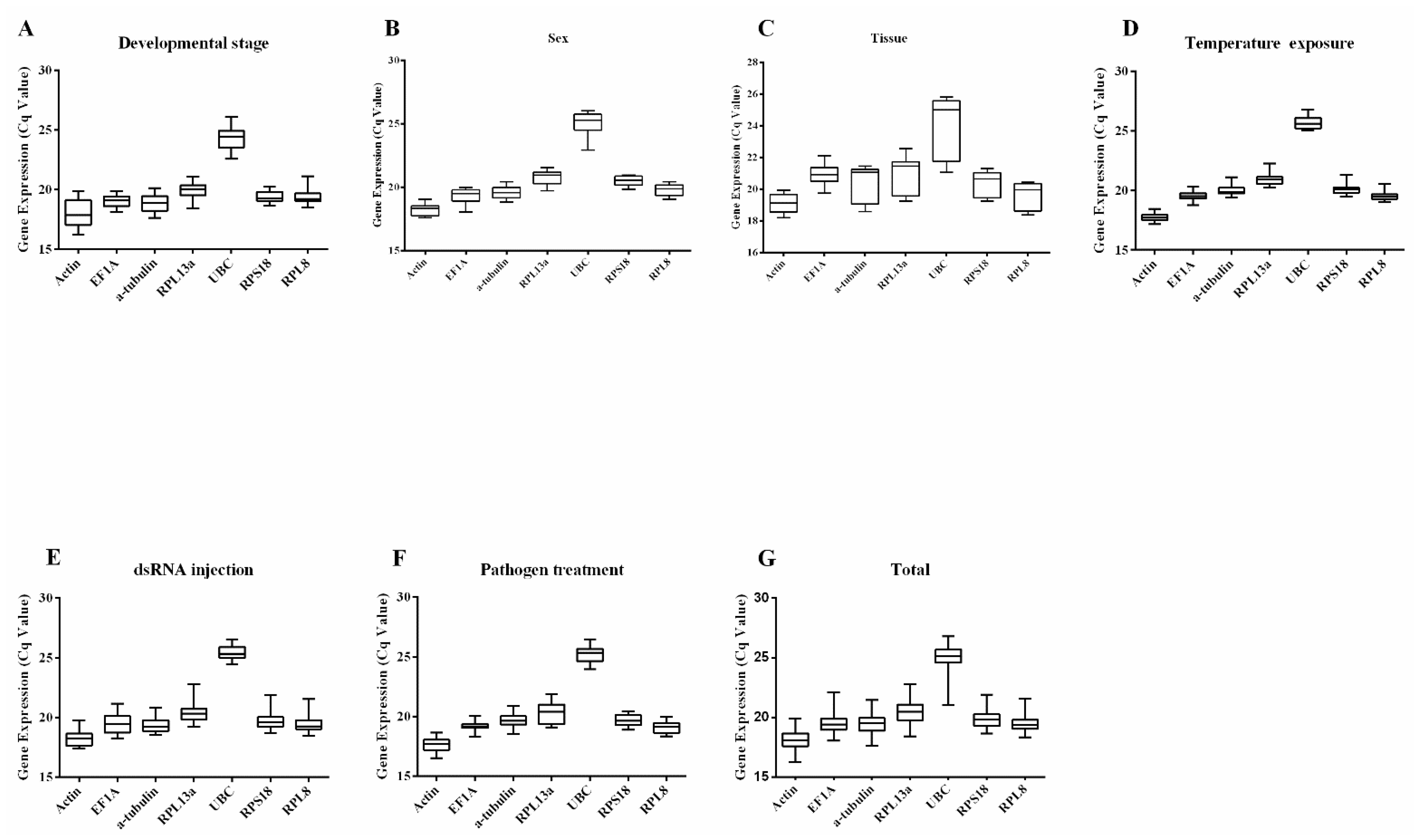
3.2. Expression Patterns of Candidate Reference Genes
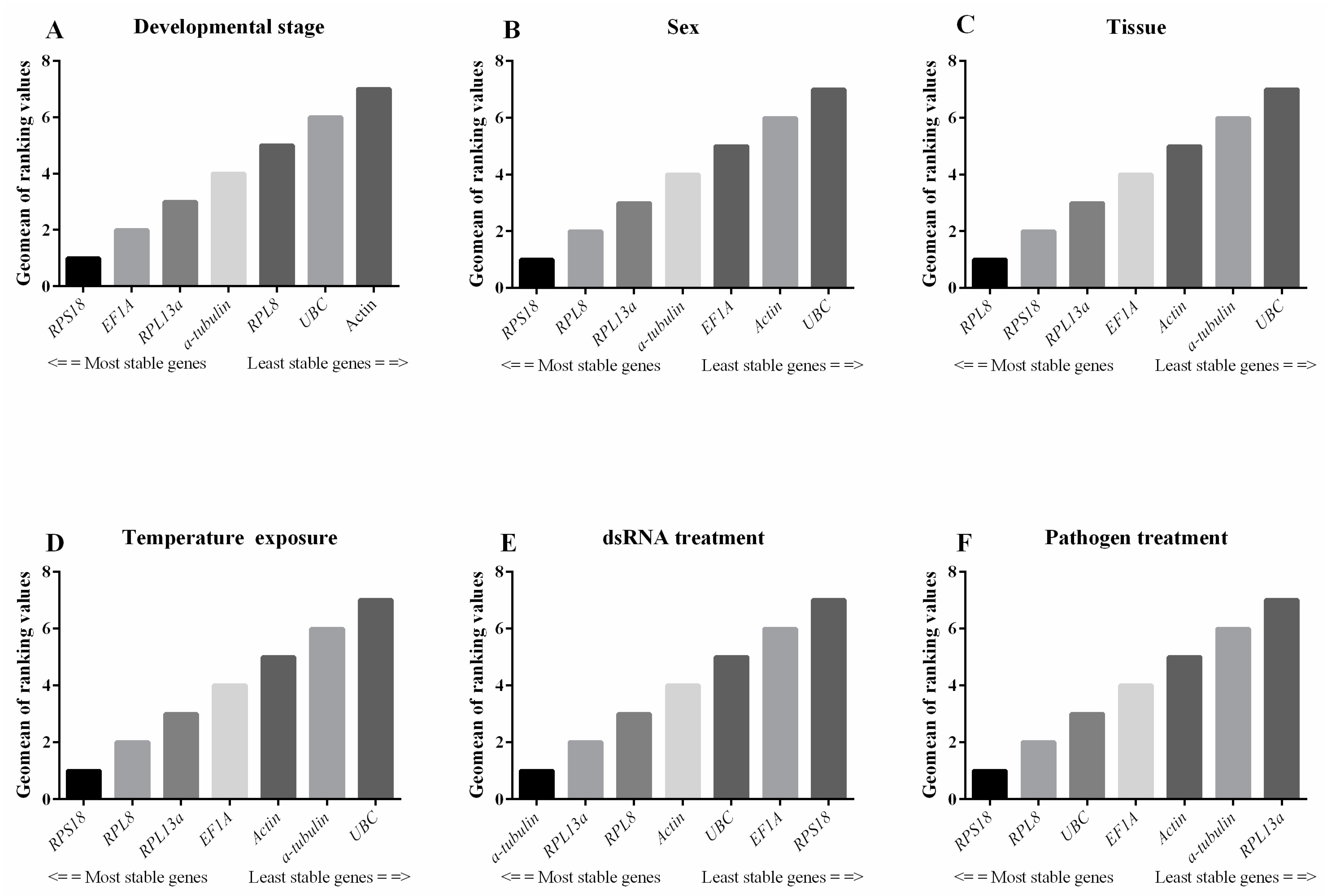
3.3. Stability of Candidate Reference Genes
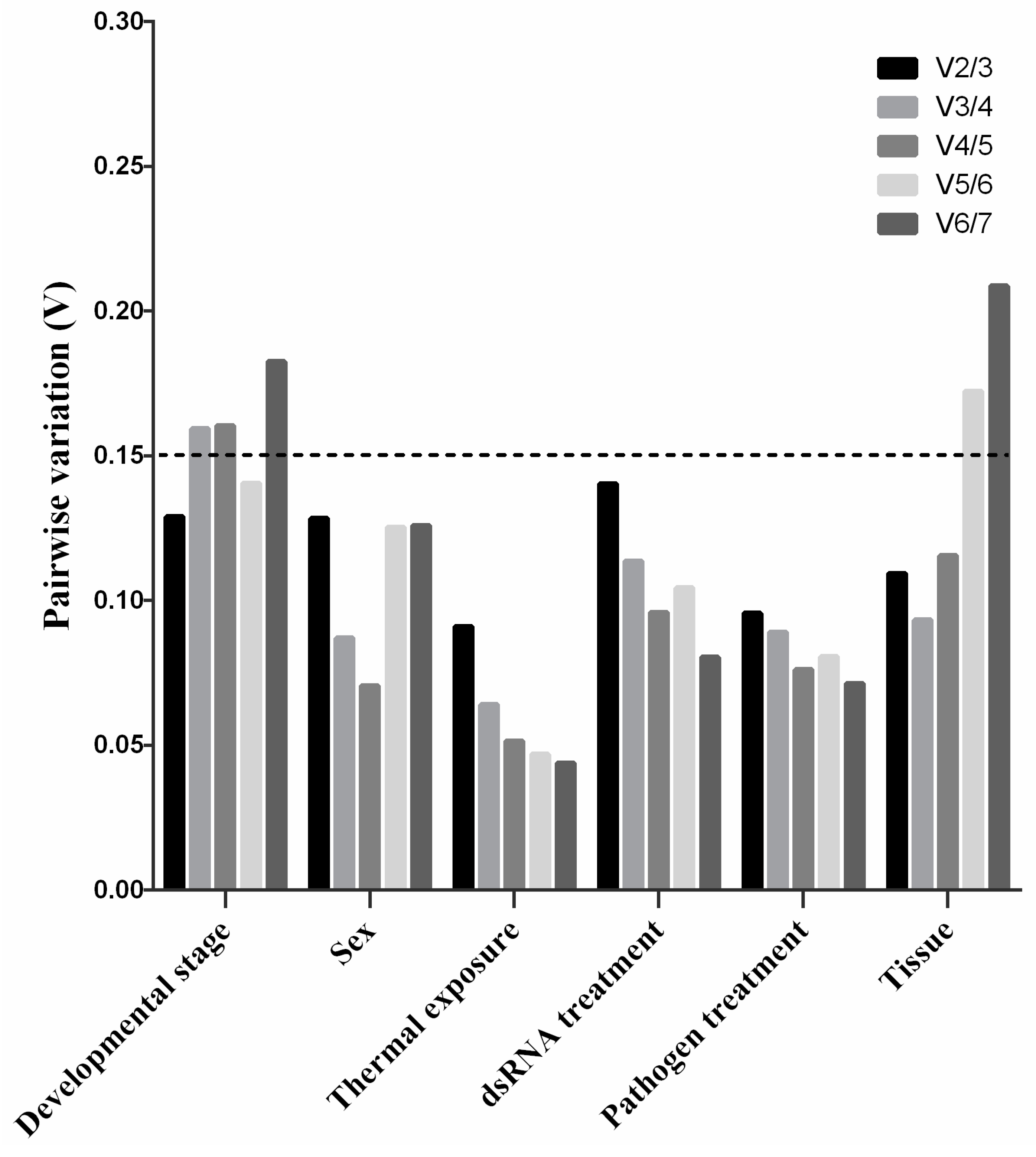
3.4. The Optimal Number of Reference Genes for Normalization in P. versicolora
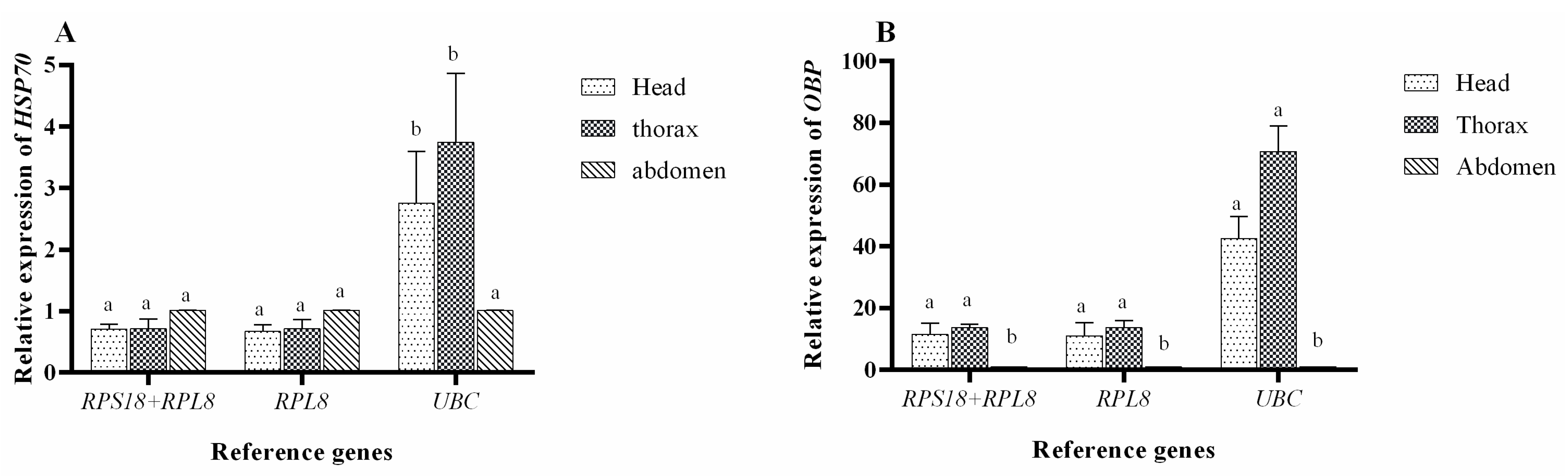
3.5. Validation of Reference Genes in P. versicolora
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shakeel, M.; Rodriguez, A.; Tahir, U.B.; Jin, F. Gene expression studies of reference genes for quantitative real-time PCR: An overview in insects. Biotechnol. Lett. 2018, 40, 227–236. [Google Scholar] [CrossRef]
- Bustin, S.A.; Mueller, R. Real-time reverse transcription PCR (qRT-PCR) and its potential use in clinical diagnosis. Clin. Sci. 2005, 109, 365–379. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.; Bustin, S.A. Reliability of real-time reverse-transcription PCR in clinical diagnostics: Gold standard or substandard? Expert Rev. Mol. Diagn. 2009, 9, 187–197. [Google Scholar] [CrossRef]
- Vogels, C.B.F.; Brito, A.F.; Wyllie, A.L.; Fauver, J.R.; Ott, I.M.; Kalinich, C.C.; Petrone, M.E.; Casanovas-Massana, A.; Catherine Muenker, M.; Moore, A.J.; et al. Analytical sensitivity and efficiency comparisons of SARS-CoV-2 RT-qPCR primer-probe sets. Nat. Microbiol. 2020, 5, 1299–1305. [Google Scholar] [CrossRef]
- He, S.; Chen, H.; Wei, Y.; An, T.; Liu, S. Development of a DNA-based real-time PCR assay for the quantification of Colletotrichum camelliae growth in tea (Camellia sinensis). Plant Methods 2020, 16, 17. [Google Scholar] [CrossRef] [Green Version]
- Milstein, S.; Nguyen, M.; Meyers, R.; de Fougerolles, A. Measuring RNAi knockdown using qPCR. Methods Enzym. 2013, 533, 57–77. [Google Scholar] [CrossRef]
- Zhang, B.; Leonard, S.P.; Li, Y.; Moran, N.A. Obligate bacterial endosymbionts limit thermal tolerance of insect host species. Proc. Natl. Acad. Sci. USA 2019, 116, 24712–24718. [Google Scholar] [CrossRef]
- Raza, M.F.; Wang, Y.; Cai, Z.; Bai, S.; Yao, Z.; Awan, U.A.; Zhang, Z.; Zheng, W.; Zhang, H. Gut microbiota promotes host resistance to low-temperature stress by stimulating its arginine and proline metabolism pathway in adult Bactrocera dorsalis. PLoS Pathog. 2020, 16, e1008441. [Google Scholar] [CrossRef] [Green Version]
- Fleige, S.; Pfaffl, M.W. RNA integrity and the effect on the real-time qRT-PCR performance. Mol. Asp. Med. 2006, 27, 126–139. [Google Scholar] [CrossRef]
- Ruijter, J.M.; Barnewall, R.J.; Marsh, I.B.; Szentirmay, A.N.; Quinn, J.C.; van Houdt, R.; Gunst, Q.D.; van den Hoff, M.J.B. Efficiency Correction Is Required for Accurate Quantitative PCR Analysis and Reporting. Clin. Chem. 2021, 67, 829–842. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Nolan, T.; Pfaffl, M.W. Quantitative real-time RT-PCR—A perspective. J. Mol. Endocrinol. 2005, 34, 597–601. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Guenin, S.; Mauriat, M.; Pelloux, J.; Van Wuytswinkel, O.; Bellini, C.; Gutierrez, L. Normalization of qRT-PCR data: The necessity of adopting a systematic, experimental conditions-specific, validation of references. J. Exp. Bot. 2009, 60, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Sang, J.; Wang, Z.; Li, M.; Cao, J.; Niu, G.; Xia, L.; Zou, D.; Wang, F.; Xu, X.; Han, X.; et al. ICG: A wiki-driven knowledgebase of internal control genes for RT-qPCR normalization. Nucleic Acids Res. 2018, 46, D121–D126. [Google Scholar] [CrossRef] [Green Version]
- Kozera, B.; Rapacz, M. Reference genes in real-time PCR. J. Appl. Genet. 2013, 54, 391–406. [Google Scholar] [CrossRef] [Green Version]
- Sang, W.; Ma, W.H.; Qiu, L.; Zhu, Z.H.; Lei, C.L. The involvement of heat shock protein and cytochrome P450 genes in response to UV-A exposure in the beetle Tribolium castaneum. J. Insect Physiol. 2012, 58, 830–836. [Google Scholar] [CrossRef]
- Zhou, L.; Meng, J.Y.; Ruan, H.Y.; Yang, C.L.; Zhang, C.Y. Expression stability of candidate RT-qPCR housekeeping genes in Spodoptera frugiperda (Lepidoptera: Noctuidae). Arch. Insect Biochem. Physiol. 2021, 108, e21831. [Google Scholar] [CrossRef]
- Li, M.; Li, X.; Wang, C.; Li, Q.; Zhu, S.; Zhang, Y.; Li, X.; Yang, F.; Zhu, X. Selection and Validation of Reference Genes For qRT-PCR Analysis of Rhopalosiphum padi (Hemiptera: Aphididae). Front. Physiol. 2021, 12, 663338. [Google Scholar] [CrossRef]
- Yin, J.; Sun, L.; Zhang, Q.; Cao, C. Screening and evaluation of the stability of expression of reference genes in Lymantria dispar (Lepidoptera: Erebidae) using qRT-PCR. Gene 2020, 749, 144712. [Google Scholar] [CrossRef]
- Xie, M.; Zhong, Y.; Lin, L.; Zhang, G.; Su, W.; Ni, W.; Qu, M.; Chen, H. Evaluation of reference genes for quantitative real-time PCR normalization in the scarab beetle Holotrichia oblita. PLoS ONE 2020, 15, e0240972. [Google Scholar] [CrossRef]
- Guo, Y.Q.; Yang, Y.; Chai, Y.; Gao, L.L.; Ma, R. Identification and Evaluation of Reference Genes for Quantitative PCR Normalization in Alligator Weed Flea Beetle (Coleoptera: Chrysomelidae). J. Insect Sci. 2021, 21, 9. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, D.H.; Siegfried, B.D. Selection of reference genes for normalization of RT-qPCR data in gene expression studies in Anthonomus eugenii Cano (Coleoptera: Curculionidae). Sci. Rep. 2020, 10, 5070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Chen, S.; Guo, M.; Ye, C.; Qiu, B.; Yang, C.; Pan, H. Selection of appropriate reference genes for RT-qPCR analysis in Propylea japonica (Coleoptera: Coccinellidae). PLoS ONE 2018, 13, e0208027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Ma, C.; Li, Z.; Zhu, B.-q.; Zhang, J.; Lei, C.-l.; Jin, S.-x.; Hull, J.J.; Chen, L.-z. Assessment of suitable reference genes for qRT-PCR analysis in Adelphocoris suturalis. J. Integr. Agric. 2018, 17, 2745–2757. [Google Scholar] [CrossRef]
- Ling, J.; Li, X.; Yang, G.; Yin, T. Volatile metabolites of willows determining host discrimination by adult Plagiodera versicolora. J. For. Res. 2021, 33, 679–687. [Google Scholar] [CrossRef]
- Demirci, M.; Sevim, E.; Demir, İ.; Sevim, A. Culturable bacterial microbiota of Plagiodera versicolora (L.) (Coleoptera: Chrysomelidae) and virulence of the isolated strains. Folia Microbiol. 2013, 58, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, L.; Li, S.; Zhang, J. Bacteria-Mediated RNA Interference for Management of Plagiodera versicolora (Coleoptera: Chrysomelidae). Insects 2019, 10, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Xu, S.; Sun, L.; Zhang, Y.; Luo, J.; Bock, R.; Zhang, J. Synergistic action of the gut microbiota in environmental RNA interference in a leaf beetle. Microbiome 2021, 9, 98. [Google Scholar] [CrossRef]
- Xu, S.; Zhang, Y.; Li, S.; Chang, L.; Wu, Y.; Zhang, J. Plastid-expressed Bacillus thuringiensis (Bt) cry3Bb confers high mortality to a leaf eating beetle in poplar. Plant Cell Rep. 2020, 39, 317–323. [Google Scholar] [CrossRef]
- Ma, M.; Tu, C.; Luo, J.; Lu, M.; Zhang, S.; Xu, L. Metabolic and immunological effects of gut microbiota in leaf beetles at the local and systemic levels. Integr. Zool. 2021, 16, 313–323. [Google Scholar] [CrossRef]
- Liu, X.; Tong, N.; Wu, Z.; Li, Y.; Ma, M.; Liu, P.; Lu, M. Identification of Chemosensory Genes Based on the Antennal Transcriptomic Analysis of Plagiodera versicolora. Insects 2021, 13, 36. [Google Scholar] [CrossRef] [PubMed]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, Research0034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper--Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Yun, J.S. Studies on Entomopathogenic Fungi Isolated from Dead Pine Caterpillars, Dendrolimus spectabilis. Entomol. Res. 2003, 33, 247–252. [Google Scholar] [CrossRef]
- Jaber, S.; Mercier, A.; Knio, K.; Brun, S.; Kambris, Z. Isolation of fungi from dead arthropods and identification of a new mosquito natural pathogen. Parasit Vectors 2016, 9, 491. [Google Scholar] [CrossRef] [Green Version]
- Radonic, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. Commun. 2004, 313, 856–862. [Google Scholar] [CrossRef]
- De Spiegelaere, W.; Dern-Wieloch, J.; Weigel, R.; Schumacher, V.; Schorle, H.; Nettersheim, D.; Bergmann, M.; Brehm, R.; Kliesch, S.; Vandekerckhove, L.; et al. Reference gene validation for RT-qPCR, a note on different available software packages. PLoS ONE 2015, 10, e0122515. [Google Scholar] [CrossRef] [Green Version]
- Behnke, J.; Mann, M.J.; Scruggs, F.L.; Feige, M.J.; Hendershot, L.M. Members of the Hsp70 Family Recognize Distinct Types of Sequences to Execute ER Quality Control. Mol. Cell 2016, 63, 739–752. [Google Scholar] [CrossRef] [Green Version]
- Shiota, M.; Kusakabe, H.; Izumi, Y.; Hikita, Y.; Nakao, T.; Funae, Y.; Miura, K.; Iwao, H. Heat shock cognate protein 70 is essential for Akt signaling in endothelial function. Arter. Thromb. Vasc. Biol. 2010, 30, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Ren, D.; Zhao, L.; Jiang, F.; Song, J.; Wang, X.; Kang, L. Identification of Odorant-Binding Proteins (OBPs) and Functional Analysis of Phase-Related OBPs in the Migratory Locust. Front. Physiol. 2018, 9, 984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, S.; Yin, J.; Zhong, T.; Cao, Y.; Li, K. Function and immunocytochemical localization of two novel odorant-binding proteins in olfactory sensilla of the scarab beetle Holotrichia oblita Faldermann (Coleoptera: Scarabaeidae). Chem. Senses 2012, 37, 141–150. [Google Scholar] [CrossRef]
- Wang, L.; Yin, H.; Zhu, Z.; Yang, S.; Fan, J. A Detailed Spatial Expression Analysis of Wing Phenotypes Reveals Novel Patterns of Odorant Binding Proteins in the Soybean Aphid, Aphis glycines. Front. Physiol. 2021, 12, 702973. [Google Scholar] [CrossRef]
- Urban, J. Contribution to the knowledge of development and harmfulness of imported willow leaf beetle (Plagiodera versicolora) (Coleoptera, Chrysomelidae). J. For. Sci. 2005, 51, 481–507. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.W.; Sheng, L.J.; Wan, Y.; Weng, X.Q.; Liang, G.H.; Zhang, F.P.; Chen, H. The complete mitochondrial genome of Plagiodera versicolora (Laicharting)(Coleoptera: Chrysomelidae). Mitochondrial DNA B Resour. 2020, 5, 3600–3601. [Google Scholar] [CrossRef] [PubMed]
- Qu, C.; Wang, R.; Che, W.; Zhu, X.; Li, F.; Luo, C. Selection and evaluation of reference genes for expression analysis using quantitative real-time PCR in the Asian Ladybird Harmonia axyridis (Coleoptera: Coccinellidae). PLoS ONE 2018, 13, e0192521. [Google Scholar] [CrossRef]
- Chen, H.; Qu, M.; Ali, F.; Lin, L.; Xie, M.; Zhang, G.; Su, W. Expression Analysis of Odorant-Binding Protein Genes and Chemosensory Protein Genes in Anomala corpulenta Motschulsky (Coleoptera: Scarabaeidae). J. Kans. Entomol. Soc. 2019, 92, 376. [Google Scholar] [CrossRef]
- Yang, C.; Preisser, E.L.; Zhang, H.; Liu, Y.; Dai, L.; Pan, H.; Zhou, X. Selection of Reference Genes for RT-qPCR Analysis in Coccinella septempunctata to Assess Un-intended Effects of RNAi Transgenic Plants. Front. Plant Sci. 2016, 7, 1672. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Sun, M.; Lu, M.-X.; Tang, X.-T.; Du, Y.-Z. Exploring Valid Reference Genes for Quantitative Real-Time PCR Analysis in Sesamia inferens (Lepidoptera: Noctuidae). PLoS ONE 2015, 10, e0115979. [Google Scholar] [CrossRef]
- Long, M.; Ting, J.; Xiangya, L.; Haijun, X.; Yingchuan, P.; Wanna, Z. Evaluation of candidate reference genes for gene expression analysis in the brassica leaf beetle, Phaedon brassicae (Coleoptera: Chrysomelidae). PLoS ONE 2021, 16, e0251920. [Google Scholar] [CrossRef]
- Fu, H.; Huang, T.; Yin, C.; Xu, Z.; Li, C.; Liu, C.; Wu, T.; Song, F.; Feng, F.; Yang, F. Selection and Validation of Reference Genes for RT-qPCR Normalization in Bradysia odoriphaga (Diptera: Sciaridae) Under Insecticides Stress. Front. Physiol. 2021, 12, 818210. [Google Scholar] [CrossRef] [PubMed]
- Ulferts, S.; Prajapati, B.; Grosse, R.; Vartiainen, M.K. Emerging Properties and Functions of Actin and Actin Filaments Inside the Nucleus. Cold Spring Harb. Perspect. Biol. 2021, 13, a040121. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Yang, X.; Siegfried, B.D.; Zhou, X. A Comprehensive Selection of Reference Genes for RT-qPCR Analysis in a Predatory Lady Beetle, Hippodamia convergens (Coleoptera: Coccinellidae). PLoS ONE 2015, 10, e0125868. [Google Scholar] [CrossRef] [PubMed]
- Pabinger, S.; Rodiger, S.; Kriegner, A.; Vierlinger, K.; Weinhausel, A. A survey of tools for the analysis of quantitative PCR (qPCR) data. Biomol. Detect. Quantif. 2014, 1, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Elin, V.; Frank, P.; An, M. Reference gene screening of Batrachochytrium dendrobatidis and Batrachochytrium salamandrivorans for quantitative real-time PCR studies. Sci. Rep. 2019, 9, 18534. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wen, H.; Li, Y.; Zhang, K.; Liu, Y. Evaluation of potential reference genes for quantitative RT-PCR analysis in spotted sea bass (Lateolabrax maculatus) under normal and salinity stress conditions. PeerJ 2018, 6, e5631. [Google Scholar] [CrossRef] [Green Version]
- Kai, F.; Jie-xia, L.; Guo-Ming, X.; Sheng, S.; Sen, L.; Ao-Qi, D.; Feng, W.; Meng-Yao, L.; Zhi-Sheng, X.; Ai-Sheng, X. Selection of appropriate reference genes for RT-qPCR analysis under abiotic stress and hormone treatment in celery. PeerJ 2019, 7, e7925. [Google Scholar] [CrossRef]
- Gothandapani, S.; Shan, A.; Jan, B.; Jirí, S.; Roman, M.; Madhab Kumar, S.; Amrita, C.; Amit, R. Reference Gene Selection for Normalizing Gene Expression in Ips Sexdentatus (Coleoptera: Curculionidae: Scolytinae) Under Different Experimental Conditions. Front. Physiol. 2021, 12, 752768. [Google Scholar] [CrossRef]
- Xie, J.; Liu, T.; Adel, K.; Yi, C.; Liu, X.; Zhang, Y. Identification and Evaluation of Suitable Reference Genes for RT-qPCR Analysis in Hippodamia variegata (Coleoptera: Coccinellidae) Under Different Biotic and Abiotic Conditions. Front. Physiol. 2021, 12, 669510. [Google Scholar] [CrossRef]
- Zhu, S.; Liu, Y.; Liao, M.; Yang, Y.; Bai, Y.; Li, N.; Li, S.; Luan, Y.; Chen, N. Evaluation of Reference Genes for Transcriptional Profiling in Two Cockroach Models. Genes 2021, 12, 1880. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Pan, H.; Noland, J.E.; Zhang, D.; Zhang, Z.; Liu, Y.; Zhou, X. Selection of reference genes for RT-qPCR analysis in a predatory biological control agent, Coleomegilla maculata (Coleoptera: Coccinellidae). Sci. Rep. 2015, 5, 18201. [Google Scholar] [CrossRef] [PubMed]
- Adeyinka, O.S.; Tabassum, B.; Nasir, I.A.; Yousaf, I.; Sajid, I.A.; Shehzad, K.; Batcho, A.; Husnain, T. Identification and validation of potential reference gene for effective dsRNA knockdown analysis in Chilo partellus. Sci. Rep. 2019, 9, 13629. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, F.; Zhang, W.; Zhang, X.; Qu, C.; Tetreau, G.; Sun, L.; Luo, C.; Zhou, J. Identification and expression profile analysis of odorant binding protein and chemosensory protein genes in Bemisia tabaci MED by head transcriptome. PLoS ONE 2017, 12, e0171739. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Luo, D.; Wu, P.; Li, H.; Zhang, H.; Zheng, W. Identification and expression profiles of novel odorant binding proteins and functional analysis of OBP99a in Bactrocera dorsalis. Arch. Insect Biochem. Physiol. 2018, 98, e21452. [Google Scholar] [CrossRef]
- Wei, H.S.; Qin, J.H.; Cao, Y.Z.; Li, K.B.; Yin, J. Two classic OBPs modulate the responses of female Holotrichia oblita to three major ester host plant volatiles. Insect Mol. Biol. 2021, 30, 390–399. [Google Scholar] [CrossRef]
- Chen, X.F.; Xu, L.; Zhang, Y.X.; Wei, D.; Wang, J.J.; Jiang, H.B. Genome-wide identification and expression profiling of odorant-binding proteins in the oriental fruit fly, Bactrocera dorsalis. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 31, 100605. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Zhang, L. Identification of putative odorant binding proteins in the peach fruit borer Carposina sasakii Matsumura (Lepidoptera: Carposinidae) by transcriptome analysis and their expression profile. Biochem. Biophys. Res. Commun. 2019, 508, 1024–1030. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Primer Sequence (5′→3′) | Product Length (bp) | R2 | E |
|---|---|---|---|---|---|
| Actin | OM885970 | F:CGTGACTTGACCGACTACCT | 118 | 0.999 | 103.3% |
| R:CGAGAGCGACATAGCAGAGT | |||||
| EF1α | OM885971 | F:TGACTCCAAGGGTGAAGGCG | 171 | 0.998 | 100.1% |
| R:TCATCGATGCTCCCGGACAC | |||||
| α-tubulin | OM885972 | F:TGGTGTCCCACCGGTTTCAA | 146 | 0.999 | 101.6% |
| R:TTGTGATCCAGACGTGCCCA | |||||
| RPL13a | OM885973 | F:AAGTGGAATGGTCCTCGGGC | 167 | 0.999 | 99.7% |
| R:CGTCTTGCGGCAATCGTAGC | |||||
| UBC | OM885974 | F:TGGCTACGTTCTCGTGGGTG | 150 | 0.998 | 105% |
| R:ACTTTTGGCGCTGCGAACTG | |||||
| RPL18S | OM885975 | F:CTTCCTCGTCGGAGCATTCT | 110 | 0.999 | 102.2% |
| R:GTTCGCCTTAACTGCCATCAA | |||||
| RPL8 | OM885976 | F:CGACCACCACCAGCTACGAT | 157 | 0.997 | 96.6% |
| R:ACCGTGGTCGATTGGCTAGG |
| Rank | GeNorm | NormFinder | BestKeeper | ΔCt | RefFinder | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | ||
| Developmental stage | 1 | RPL13a | 0.368 | RPS18 | 0.065 | RPS18 | 0.370 | RPS18 | 0.714 | RPS18 | 1.000 |
| RPS18 | 0.368 | ||||||||||
| 2 | - | - | EF1A | 0.131 | EF1A | 0.380 | EF1A | 0.718 | EF1A | 2.213 | |
| 3 | EF1A | 0.407 | RPL13a | 0.258 | RPL8 | 0.415 | RPL13a | 0.785 | RPL13a | 2.449 | |
| 4 | α-tubulin | 0.542 | α-tubulin | 0.403 | RPL13a | 0.498 | α-tubulin | 0.878 | α-tubulin | 4.229 | |
| 5 | RPL8 | 0.673 | RPL8 | 0.511 | α-tubulin | 0.616 | RPL8 | 0.977 | RPL8 | 4.401 | |
| 6 | UBC | 0.764 | UBC | 0.647 | UBC | 0.748 | UBC | 1.092 | UBC | 6.000 | |
| 7 | Actin | 0.930 | Actin | 0.872 | Actin | 0.954 | Actin | 1.343 | Actin | 7.000 | |
| Sex | 1 | RPS18 | 0.277 | RPS18 | 0.096 | RPS18 | 0.287 | RPS18 | 0.508 | RPS18 | 1.000 |
| RPL8 | 0.277 | ||||||||||
| 2 | - | - | RPL8 | 0.125 | RPL8 | 0.341 | RPL13a | 0.514 | RPL8 | 1.861 | |
| 3 | RPL13a | 0.365 | RPL13a | 0.148 | Actin | 0.364 | RPL8 | 0.525 | RPL13a | 3.080 | |
| 4 | EF1A | 0.381 | α-tubulin | 0.243 | α-tubulin | 0.407 | EF1A | 0.570 | α-tubulin | 4.472 | |
| 5 | α-tubulin | 0.398 | EF1A | 0.243 | RPL13a | 0.469 | α-tubulin | 0.580 | EF1A | 4.681 | |
| 6 | UBC | 0.530 | UBC | 0.553 | EF1A | 0.527 | UBC | 0.880 | Actin | 5.664 | |
| 7 | Actin | 0.644 | Actin | 0.600 | UBC | 0.836 | Actin | 0.931 | UBC | 6.236 | |
| Thermal exposure | 1 | RPL13a | 0.255 | RPS18 | 0.114 | Actin | 0.218 | RPS18 | 0.294 | RPS18 | 1.414 |
| RPS18 | 0.255 | ||||||||||
| 2 | - | - | RPL8 | 0.146 | RPL8 | 0.247 | RPL8 | 0.318 | RPL8 | 2.515 | |
| 3 | UBC | 0.286 | RPL13a | 0.153 | EF1A | 0.267 | RPL13a | 0.321 | RPL13a | 2.711 | |
| 4 | α-tubulin | 0.296 | EF1A | 0.164 | 18S | 0.278 | EF1A | 0.332 | EF1A | 4.120 | |
| 5 | RPL8 | 0.303 | α-tubulin | 0.167 | α-tubulin | 0.321 | α-tubulin | 0.334 | Actin | 4.304 | |
| 6 | EF1A | 0.315 | UBC | 0.180 | RPL13a | 0.401 | UBC | 0.343 | α-tubulin | 4.729 | |
| 7 | Actin | 0.329 | Actin | 0.203 | UBC | 0.422 | Actin | 0.364 | UBC | 5.244 | |
| dsRNA treatment | 1 | RPL13a | 0.454 | α-tubulin | 0.146 | α-tubulin | 0.457 | α-tubulin | 0.510 | α-tubulin | 1.000 |
| α-tubulin | 0.454 | ||||||||||
| 2 | - | - | RPL8 | 0.269 | UBC | 0.464 | RPL13a | 0.580 | RPL13a | 2.449 | |
| 3 | RPL8 | 0.470 | RPL13a | 0.275 | Actin | 0.477 | RPL8 | 0.586 | RPL8 | 3.080 | |
| 4 | EF1A | 0.497 | Actin | 0.299 | RPS18 | 0.501 | Actin | 0.610 | Actin | 3.936 | |
| 5 | Actin | 0.524 | EF1A | 0.352 | RPL8 | 0.557 | EF1A | 0.642 | UBC | 5.118 | |
| 6 | RPS18 | 0.584 | RPS18 | 0.360 | RPL13a | 0.603 | RPS18 | 0.655 | EF1A | 5.144 | |
| 7 | UBC | 0.607 | UBC | 0.372 | EF1A | 0.661 | UBC | 0.665 | RPS18 | 5.422 | |
| Pathogen treatment | 1 | RPS18 | 0.234 | RPS18 | 0.116 | EF1A | 0.232 | RPS18 | 0.384 | RPS18 | 1.316 |
| RPL8 | 0.234 | ||||||||||
| 2 | - | - | UBC | 0.153 | α-tubulin | 0.410 | RPL8 | 0.412 | RPL8 | 2.213 | |
| 3 | UBC | 0.284 | RPL8 | 0.161 | RPS18 | 0.420 | UBC | 0.412 | UBC | 3.224 | |
| 4 | Actin | 0.338 | Actin | 0.207 | RPL8 | 0.446 | Actin | 0.453 | EF1A | 3.344 | |
| 5 | EF1A | 0.376 | EF1A | 0.265 | Actin | 0.484 | EF1A | 0.496 | Actin | 4.229 | |
| 6 | α-tubulin | 0.431 | α-tubulin | 0.326 | UBC | 0.536 | α-tubulin | 0.563 | α-tubulin | 4.559 | |
| 7 | RPL13a | 0.469 | RPL13a | 0.335 | RPL13a | 0.752 | RPL13a | 0.566 | RPL13a | 7.000 | |
| Tissue | 1 | RPL8 | 0.099 | RPS18 | 0.034 | Actin | 0.465 | RPL8 | 0.609 | RPL8 | 1.189 |
| RPS18 | 0.099 | ||||||||||
| 2 | - | - | RPL8 | 0.034 | EF1A | 0.554 | RPS18 | 0.622 | RPS18 | 1.565 | |
| 3 | RPL13a | 0.254 | RPL13a | 0.140 | 18S | 0.676 | RPL13a | 0.687 | RPL13a | 3.409 | |
| 4 | α-tubulin | 0.330 | α-tubulin | 0.188 | RPL8 | 0.720 | α-tubulin | 0.714 | EF1A | 3.976 | |
| 5 | EF1A | 0.441 | EF1A | 0.297 | RPL13a | 0.998 | EF1A | 0.816 | Actin | 4.304 | |
| 6 | UBC | 0.646 | UBC | 0.842 | α-tubulin | 1.001 | UBC | 1.262 | α-tubulin | 4.427 | |
| 7 | Actin | 0.885 | Actin | 1.002 | UBC | 1.696 | Actin | 1.482 | UBC | 6.236 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, C.; Xu, P.; Han, R.; Luo, J.; Xu, L. Defining Suitable Reference Genes for qRT-PCR in Plagiodera versicolora (Coleoptera: Chrysomelidae) under Different Biotic or Abiotic Conditions. Agronomy 2022, 12, 1192. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051192
Tu C, Xu P, Han R, Luo J, Xu L. Defining Suitable Reference Genes for qRT-PCR in Plagiodera versicolora (Coleoptera: Chrysomelidae) under Different Biotic or Abiotic Conditions. Agronomy. 2022; 12(5):1192. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051192
Chicago/Turabian StyleTu, Chengjie, Pei Xu, Runhua Han, Jing Luo, and Letian Xu. 2022. "Defining Suitable Reference Genes for qRT-PCR in Plagiodera versicolora (Coleoptera: Chrysomelidae) under Different Biotic or Abiotic Conditions" Agronomy 12, no. 5: 1192. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051192