
Unraveling the Genetic Architecture for Low Temperature Germinability-Related Traits in Rice Using Genome-Wide Association Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Population Structure Analysis
2.2. Indices for Evaluating LTG
2.3. Statistical Analysis, GWAS Mapping and Candidate Gene Analysis
2.4. Gene Expression Analysis
3. Results
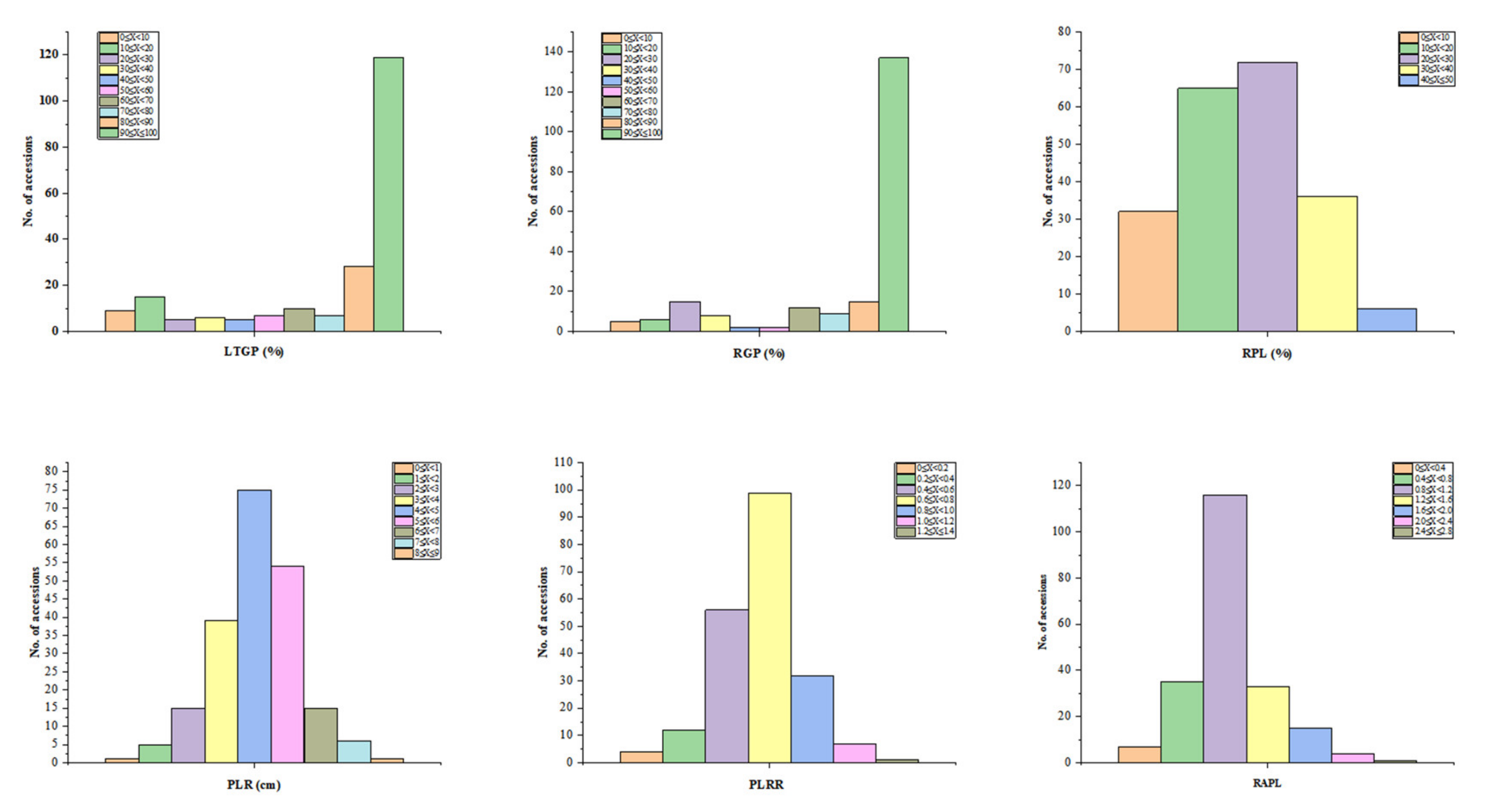
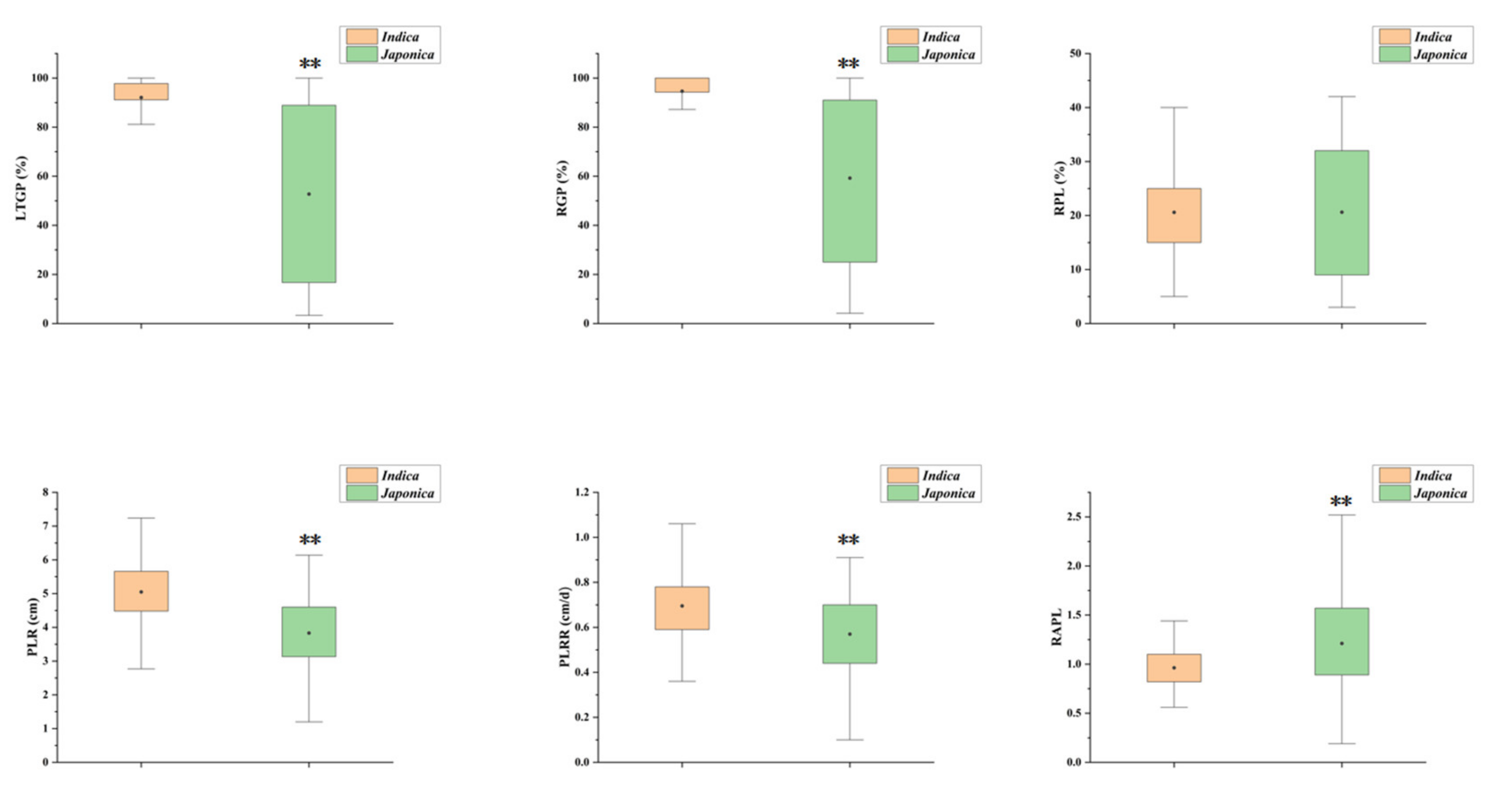
3.1. Assessment of Six LTG Indices in 211 Rice Accessions
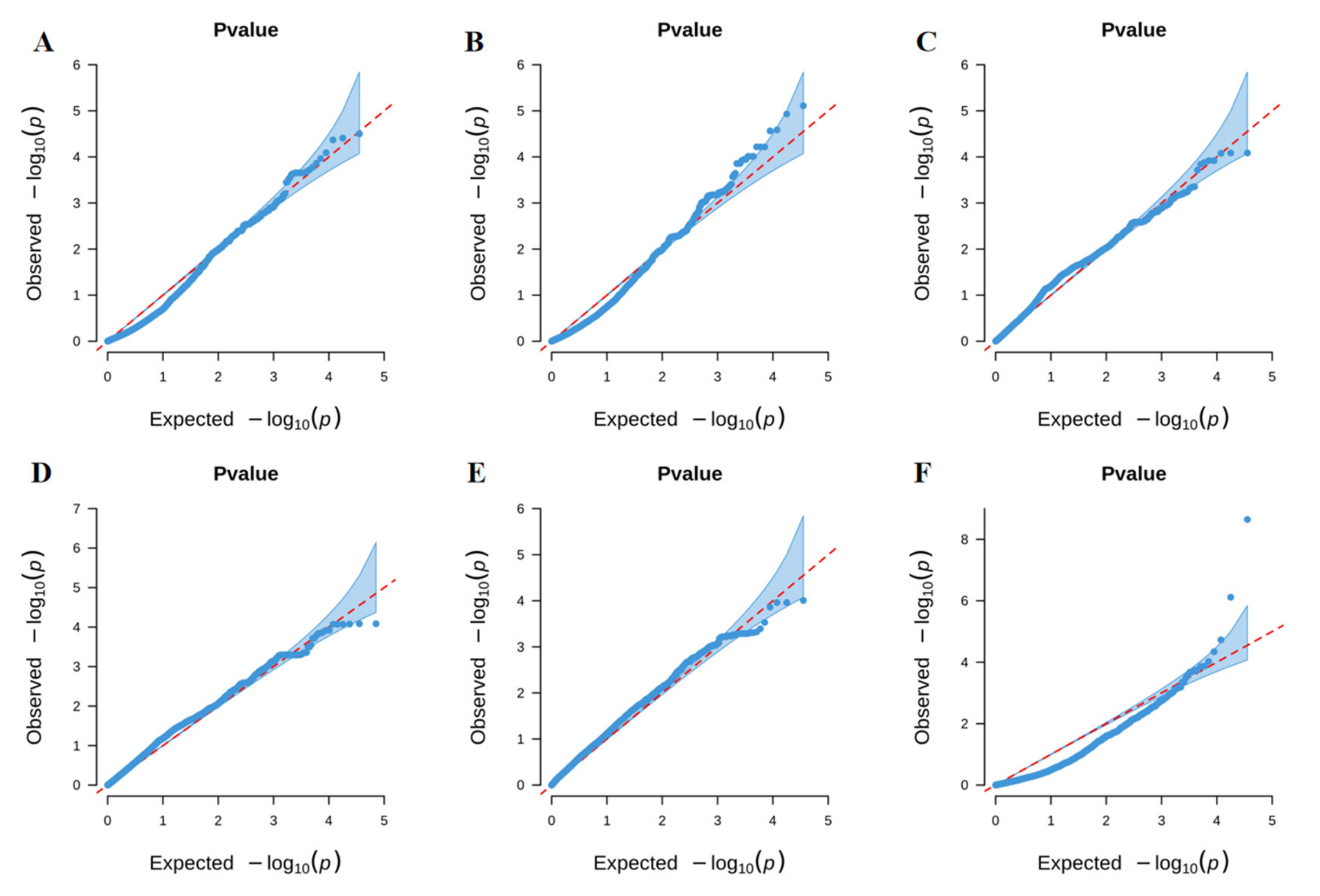
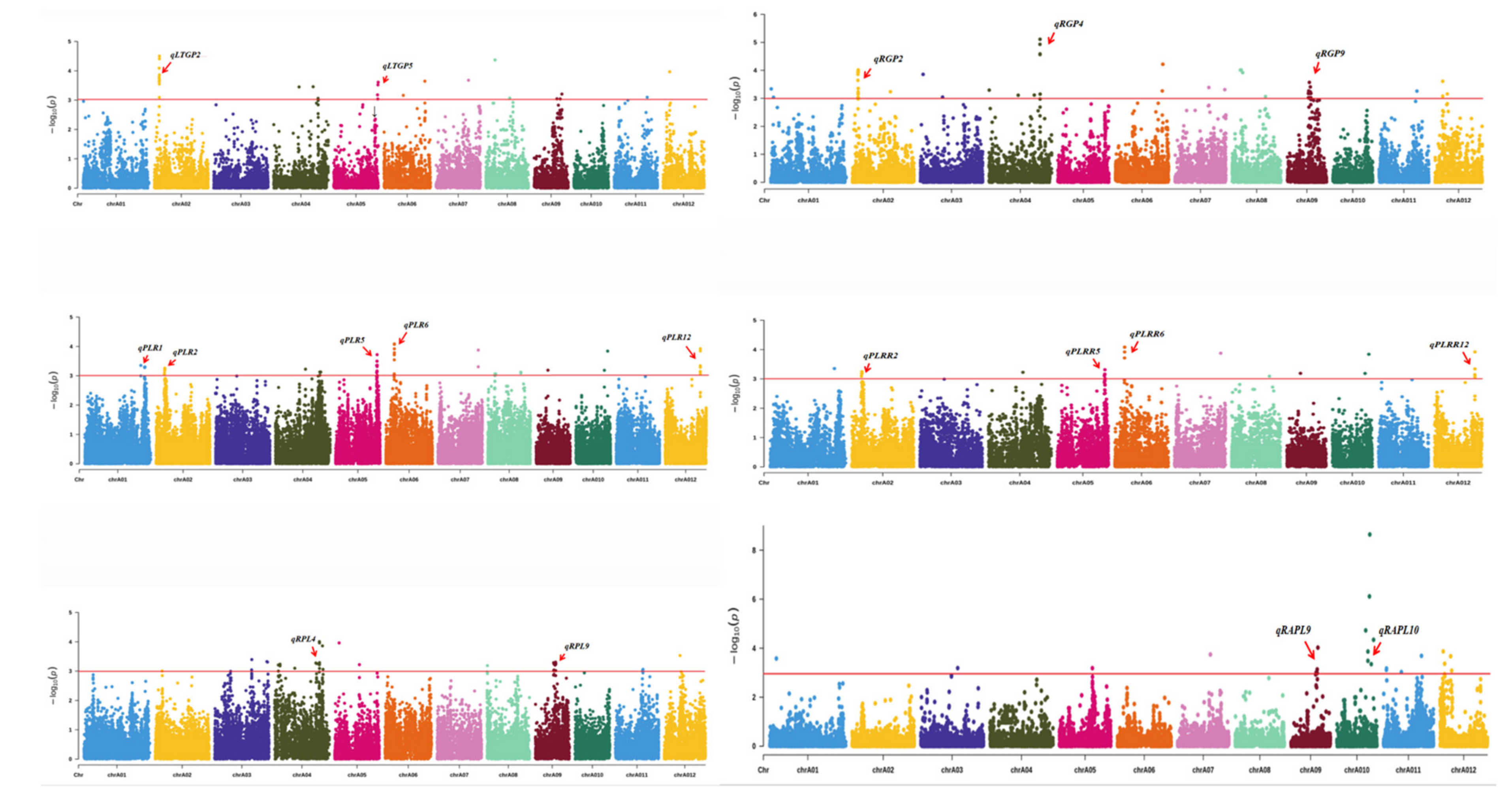
3.2. GWAS for Identification of QTLs
3.3. Candidate Gene Analysis
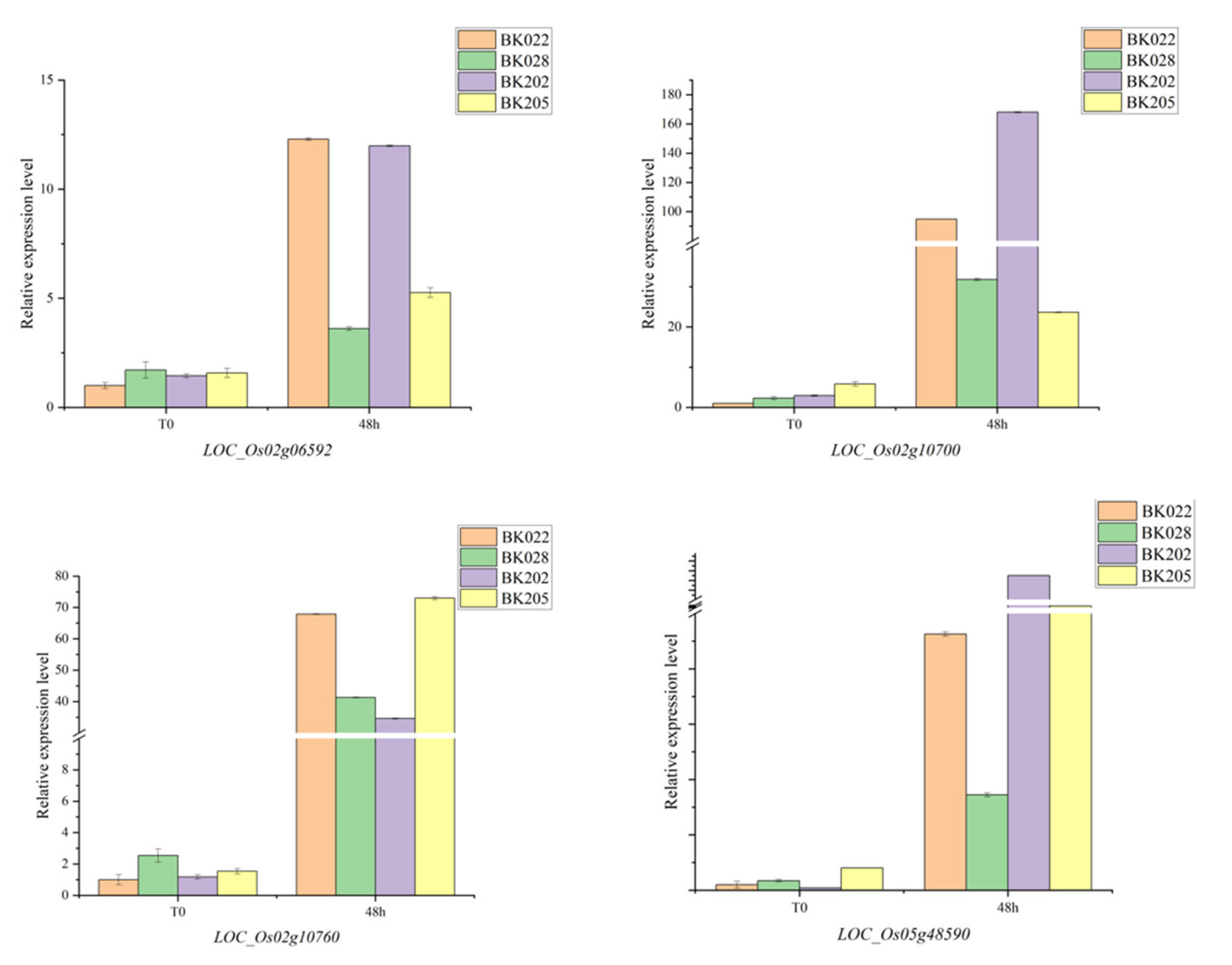
3.4. Gene Expression Analyses of Candidate Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, H.; Lee, A.R.; Park, S.Y.; Jin, S.H.; Lee, J.; Ham, T.H.; Park, Y.; Zhao, W.G.; Kwon, S.W. Genome-wide association study reveals candidate genes related to low temperature tolerance in rice (Oryza sativa) during germination. 3 Biotech 2018, 8, 235. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Zhang, H.; Zhang, D.; Li, J.; Xiong, H.; Yu, J.; Li, J.; Rashid, M.A.R.; Li, G.; Ma, X.; et al. Genetic Analysis of Cold Tolerance at the Germination and Booting Stages in Rice by Association Mapping. PLoS ONE. 2015, 10, e0120590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujino, K. A major gene for low temperature germinability in rice (Oryza sativa L.). Euphytica 2004, 136, 63–68. [Google Scholar] [CrossRef]
- Schläppi, M.R.; Jackson, A.K.; Eizenga, G.C.; Wang, A.; Chu, C.; Shi, Y.; Shimoyama, N.; Boykin, D.L. Assessment of Five Chilling Tolerance Traits and GWAS Mapping in Rice Using the USDA Mini-Core Collection. Front. Plant Sci. 2017, 8, 957. [Google Scholar] [CrossRef] [Green Version]
- Shim, K.C.; Kim, S.H.; Lee, H.S.; Adeva, C.; Jeon, Y.A.; Luong, N.H.; Kim, W.J.; Akhtamov, M.; Park, Y.J.; Ahn, S.N. Characterization of a New qLTG3–1 Allele for Low-temperature Germinability in Rice from the Wild Species Oryza rufipogon. Rice 2020, 13, 10. [Google Scholar] [CrossRef]
- Jiang, N.; Shi, S.; Shi, H.; Khanzada, H.; Wassan, G.M.; Zhu, C.; Peng, X.; Yu, Q.; Chen, X.; He, X.; et al. Mapping QTL for seed germinability under low temperature using a new high-density genetic map of rice. Front. Plant Sci. 2017, 8, 1223. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Liu, X.; Xie, K.; Wang, Y.; Liu, F.; Lin, Q.; Wang, W.; Yang, C.; Lu, B.; Liu, S.; et al. qLTG-9, a stable quantitative trait locus for low-temperature germination in rice (Oryza sativa L.). Theor. Appl. Genet. 2013, 126, 2313–2322. [Google Scholar] [CrossRef]
- Xie, L.; Tan, Z.; Zhou, Y.; Xu, R.; Feng, L. Identification and fine mapping of quantitative trait loci for seed vigor in germination and seedling establishment in rice. J. Integr. Plant Biol. 2014, 56, 749–759. [Google Scholar] [CrossRef]
- Jiang, S.; Yang, C.; Xu, Q.; Wang, L.; Yang, X.; Song, X.; Wang, J.; Zhang, X.; Li, B.; Li, H.; et al. Genetic dissection of germinability under low temperature by building a resequencing linkage map in japonica rice. Int. J. Mol. Sci. 2020, 21, 1284. [Google Scholar] [CrossRef] [Green Version]
- Fujino, K.; Sekiguchi, H.; Matsuda, Y.; Sugimoto, K.; Ono, K.; Yano, M. Molecular identification of a major quantitative trait locus, qLTG3-1, controlling low-temperature germinability in rice. Proc. Natl. Acad. Sci. USA 2008, 105, 12623–12628. [Google Scholar] [CrossRef] [Green Version]
- Fujino, K.; Matsuda, Y. Genome-wide analysis of genes targeted by qLTG3-1 controlling low-temperature germinability in rice. Plant Mol. Biol. 2010, 72, 137–152. [Google Scholar] [CrossRef]
- Mao, D.; Yu, L.; Chen, D.; Li, L.; Zhu, Y.; Xiao, Y.; Zhang, D.; Chen, C. Multiple cold resistance loci confer the high cold tolerance adaptation of Dongxiang wild rice (Oryza rufipogon) to its high-latitude habitat. Theor. Appl. Genet. 2015, 128, 1359–1371. [Google Scholar] [CrossRef]
- Fujino, K.; Obara, M.; Shimizu, T.; Koyanagi, K.O.; Ikegaya, T. Genome-wide association mapping focusing on a rice population derived from rice breeding programs in a region. Breed. Sci. 2015, 65, 403–410. [Google Scholar] [CrossRef] [Green Version]
- Shakiba, E.; Edwards, J.D.; Jodari, F. Genetic architecture of cold tolerance in rice (Oryza sativa) determined through high resolution genome-wide analysis. PLoS ONE. 2017, 12, e0172133. [Google Scholar] [CrossRef]
- Sales, E.; Viruel, J.; Domingo, C.; Marqués, L. Genome wide association analysis of cold tolerance at germination in temperate japonica rice (Oryza sativa L.) varieties. PLoS ONE. 2017, 12, e0183416. [Google Scholar] [CrossRef] [Green Version]
- Thapa, R.; Tabien, R.E.; Thomson, M.J.; Septiningsih, E.M. Genome-Wide Association Mapping to Identify Genetic Loci for Cold Tolerance and Cold Recovery During Germination in Rice. Front. Genet. 2020, 11, 22. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Zhou, L.; Zhao, J.; Dong, J.; Liu, Q.; Fu, H.; Mao, X.; Yang, W.; Ma, Y.; Chen, L.; et al. The Candidate Genes Underlying a Stably Expressed QTL for Low Temperature Germinability in Rice (Oryza sativa L.). Rice 2020, 13, 74. [Google Scholar] [CrossRef]
- Wang, X.; Zou, B.; Shao, Q.; Cui, Y.; Lu, S.; Zhang, Y.; Huang, Q.; Huang, J.; Hua, J. Natural variation reveals that OsSAP16 controls low-temperature germination in rice. J. Exp. Bot. 2018, 69, 413–421. [Google Scholar] [CrossRef]
- Li, C.; Liu, J.; Bian, J.; Jin, T.; Zou, B.; Liu, S.; Zhang, X.; Wang, P.; Tan, J.; Wu, G.; et al. Identification of cold tolerance QTLs at the bud burst stage in 211 rice accessions by GWAS. BMC Plant Biol. 2021, 21, 542. [Google Scholar] [CrossRef]
- Hu, H.; You, J.; Fang, Y.; Zhu, X.; Qi, Z.; Xiong, L. Characterization of transcription factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol. Biol. 2008, 67, 169–181. [Google Scholar] [CrossRef]
- Abe, F.; Saito, K.; Miura, K.; Toriyama, K. A single nucleotide polymorphism in the alternative oxidase gene among rice varieties differing in low temperature tolerance. FEBS Lett. 2002, 527, 181–185. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.A.; Cho, J.I.; Han, M.; Ahn, C.H.; Jeon, J.S.; An, G.; Park, P.B. The ABRE-binding bZIP transcription factor OsABF2 is a positive regulator of abiotic stress and ABA signaling in rice. J. Plant Physiol. 2010, 167, 1512–1520. [Google Scholar] [CrossRef]
- Oh, C.S.; Choi, Y.H.; Lee, S.J.; Yoon, D.B.; Moon, H.P.; Ahn, S.N. Mapping of Quantitative Trait Loci for Cold Tolerance in Weedy Rice. Breed. Sci. 2004, 54, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Ke, Y.G.; Yang, Z.J.; Yu, S.W.; Li, T.F.; Wu, J.H.; Gao, H.; Fu, Y.P.; Luo, L.J. Characterization of OsDREB6 responsive to osmotic and cold stresses in rice. Ceram. Int. 2016, 42, 9264–9269. [Google Scholar] [CrossRef]
- Yokotani, N.; Sato, Y.; Tanabe, S.; Chujo, T.; Shimizu, T.; Okada, K.; Yamane, H.; Shimono, M.; Sugano, S.; Takatsuji, H.; et al. WRKY76 is a rice transcriptional repressor playing opposite roles in blast disease resistance and cold stress tolerance. J. Exp. Bot. 2013, 64, 5085–5097. [Google Scholar] [CrossRef]
- Gao, Z.R.; Zhang, H.W.; Huang, R.F. Expressive Characteristics Analysis of OsDDMla and OsDDMlb in Response to Abiotic Stresses of Rice. J. Agric. Sci. Technol. 2011, 13, 41–46. [Google Scholar]
- Zhang, Y.; Lan, H.; Shao, Q.; Wang, R.; Chen, H.; Tang, H.; Zhang, H.; Huang, J. An A20/AN1-type zinc finger protein modulates gibberellins and abscisic acid contents and increases sensitivity to abiotic stress in rice (Oryza sativa L.). J. Exp. Bot. 2015, 67, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Wang, G.; Zhu, S. DNA Damage Inducible Protein 1 is Involved in Cold Adaption of Harvested Cucumber Fruit. Front. Plant Sci. 2020, 10, 1723. [Google Scholar] [CrossRef]
- González-Schain, N.; Roig-Villanova, I.; Kater, M.M. Early cold stress responses in post-meiotic anthers from tolerant and sensitive rice cultivars. Rice 2019, 12, 94. [Google Scholar] [CrossRef]
- Song, W.; Hao, Q.; Cai, M.; Wang, Y.; Zhu, X.; Liu, X.; Huang, Y.; Nguyen, T.; Yang, C.; Yu, J.; et al. Rice OsBT1 regulates seed dormancy through the glycometabolism pathway. Plant Physiol. Biochem. 2020, 151, 469–476. [Google Scholar] [CrossRef]
- Yang, Y.W.; Chen, H.C.; Jen, W.F.; Liu, L.Y.; Chang, M.C. Comparative Transcriptome Analysis of Shoots and Roots of TNG67 and TCN1 Rice Seedlings under Cold Stress and Following Subsequent Recovery: Insights into Metabolic Pathways, Phytohormones, and Transcription Factors. PLoS ONE. 2015, 10, e0131391. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Chen, L.; Li, D.; Lv, B.; Chen, Y.; Chen, J.; Liang, J. OsRACK1 is involved in abscisic acid- and H2O2-mediated signaling to regulate seed germination in rice (Oryza sativa L.). PLoS ONE. 2014, 9, e97120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, Y.; Sun, S.; Li, M.; Li, L.; Cao, W.; Luo, J.; Tang, H.; Huang, J.; Wang, Z.; Wang, J.; et al. Overexpression of the Qc-SNARE gene OsSYP71 enhances tolerance to oxidative stress and resistance to rice blast in rice (Oryza sativa L.). Gene 2012, 504, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wei, Q.; Zhang, K.; Wang, L.; Liu, F.; Zhao, L.; Tan, Y.; Di, C.; Yan, H.; Yu, J.; et al. Down-Regulation of OsSPX1 Causes High Sensitivity to Cold and Oxidative Stresses in Rice Seedlings. PLoS ONE 2013, 8, e81849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cen, W.; Liu, J.; Lu, S.; Jia, P.; Yu, K.; Han, Y.; Li, R.; Luo, J. Comparative proteomic analysis of QTL CTS-12 derived from wild rice (Oryza rufipogon Griff.), in the regulation of cold acclimation and de-acclimation of rice (Oryza sativa L.) in response to severe chilling stress. BMC Plant Biol. 2018, 18, 163. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Liu, H.; Qu, Z.; Wang, J.; Wang, X.; Wang, Z.; Yang, L.; Zhang, D.; Zou, D.; Zhao, H. Transcriptome Sequencing and iTRAQ of Different Rice Cultivars Provide Insight into Molecular Mechanisms of Cold-Tolerance Response in Japonica Rice. Rice 2020, 13, 43. [Google Scholar] [CrossRef]
- Zhu, N.; Cheng, S.; Liu, X.; Du, H.; Dai, M.; Zhou, D.X. The R2R3-type MYB gene OsMYB91 has a function in coordinating plant growth and salt stress tolerance in rice. Plant Sci. 2015, 236, 146–156. [Google Scholar] [CrossRef]
- Li, W.; Yang, B.; Xu, J.; Peng, L.; Sun, S.; Huang, Z.; Jiang, X.; He, Y.; Wang, Z. A genome-wide association study reveals that the 2-oxoglutarate/malate translocator mediates seed vigor in rice. Plant J. 2021, 108, 478–491. [Google Scholar] [CrossRef]
- Cui, K.; Peng, S.; Xing, Y.; Xu, C.; Yu, S.; Zhang, Q. Molecular dissection of seedling-vigor and associated physiological traits in rice. Theor. Appl. Genet. 2002, 105, 745–753. [Google Scholar] [CrossRef]
- Morsy, M.R.; Almutairi, A.M.; Gibbons, J.; Yun, S.J.; Benildo, G. The OsLti6 genes encoding low-molecular-weight membrane proteins are differentially expressed in rice cultivars with contrasting sensitivity to low temperature. Gene 2005, 344, 171–180. [Google Scholar] [CrossRef]
- Lv, Y.; Guo, Z.; Li, X.; Ye, H.; Li, X.; Xiong, L. New insights into the genetic basis of natural chilling and cold shock tolerance in rice by genome-wide association analysis. Plant Cell Environ. 2016, 39, 556–570. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
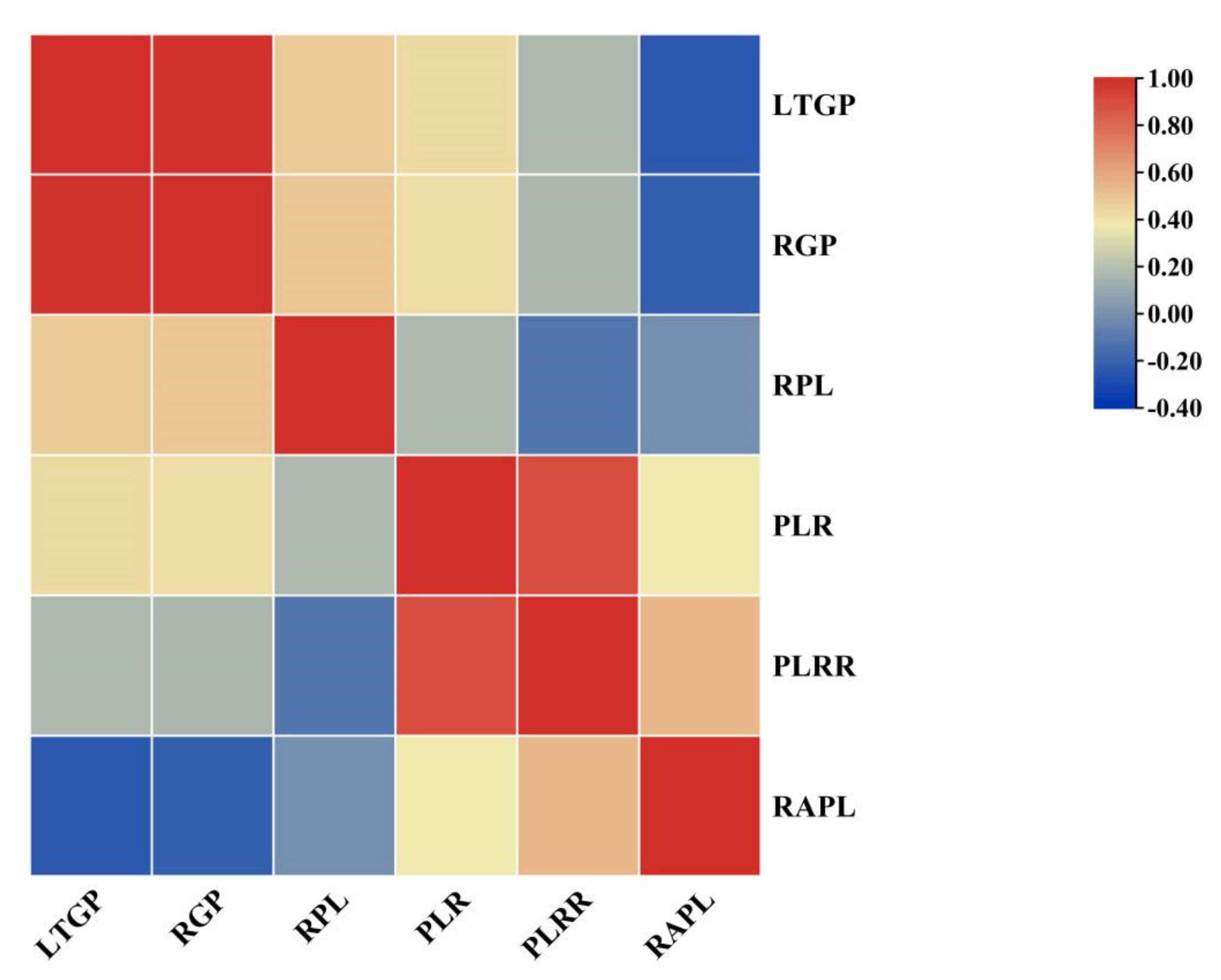
| Trait | LTGP | RGP | RPL | PLR | PLRR |
|---|---|---|---|---|---|
| RGP | 0.986 ** | ||||
| RPL | 0.483 ** | 0.497 ** | |||
| PLR | 0.429 ** | 0.417 ** | 0.167 * | ||
| PLRR | 0.168 * | 0.162 * | −0.115 | 0.895 ** | |
| RAPL | −0.243 ** | −0.217 ** | −0.008 | 0.381 ** | 0.543 ** |
| QTL ID | Chr. | Peak SNPs | p Value | R2 | Previous QTLs/Genes |
|---|---|---|---|---|---|
| qPLR1 | 1 | 39993855 | 0.000504630 | 0.060240000 | SNAC2 [20] |
| qLTGP2 | 2 | 3339289 | 0.000533207 | 0.062685322 | |
| qRGP2 | 3339289 | 0.000111934 | 0.075171265 | ||
| qPLR2 | 5505457 | 0.000816111 | 0.069550000 | ||
| qPLRR2 | 5505457 | 0.000813330 | 0.068310000 | ||
| qRGP4 | 4 | 29781197 | 0.000007752 | 0.057870000 | OsAOX1a [21] |
| qRPL4 | 29921285 | 0.000098708 | 0.085890000 | OsAOX1a | |
| qLTGP5 | 5 | 27865039 | 0.000206354 | 0.070831922 | |
| qPLR5 | 27865039 | 0.000039949 | 0.096883103 | ||
| qPLRR5 | 27865039 | 0.000991230 | 0.074340000 | ||
| qPLR6 | 6 | 5745395 | 0.000021543 | 0.103143691 | OsABF2 [22] |
| qPLRR6 | 5745395 | 0.000496299 | 0.065075903 | OsABF2 | |
| qRGP9 | 9 | 12912697 | 0.000268990 | 0.039760000 | clr9 [23]; OsDREB6 [24] |
| qRPL9 | 13641693 | 0.000494500 | 0.072930000 | clr9 | |
| qRAPL9 | 15684209 | 0.000094869 | 0.087940000 | clr9; OsWRKY76 [25] | |
| qRAPL10 | 10 | 19057293 | 0.000001319 | 0.118803366 | qLTG10-1 [4] |
| qPLR12 | 12 | 23577481 | 0.000142160 | 0.089220000 | qLTSS12-1 [4] |
| qPLRR12 | 23577481 | 0.000120220 | 0.087880000 | qLTSS12-1 [4] |
| QTLs ID | Candidate Genes | |||
|---|---|---|---|---|
| Locus | Gene | Protein | Description | |
| qLTGP2 | LOC_Os02g06592 | CHR701 | SNF2 family N-terminal domain containing protein | Snf2 family proteins can be induced by various abiotic stresses [26]. |
| qPLR2 | LOC_Os02g10200 | ZFP185 | A20/AN1-type zinc finger protein | The expression level of ZFP185 was susceptible to salt stress, osmotic stress and cold stress [27]. |
| LOC_Os02g10510 | OsDDI1 | Ubiquitin family domain-containing protein | CsDDI1 is involved in the physiological and molecular mechanisms of cold acclimation [28]. | |
| LOC_Os02g10700 | OsEBF2 | OsFBL7—F-box domain- and LRR-containing protein | OsEBF2 involved in ethylene signaling in anthers is upregulated after cold stress in rice [29]. | |
| LOC_Os02g10760 | OsWR1 | AP2 domain-containing protein | OsWR1 is also a gene associated with ethylene metabolism in anthers, which is slightly downregulated after cold stress [29]. | |
| LOC_Os02g10800 | OsBT1 | mitochondrial carrier protein | OsBT1 is involved in seed germination and regulates seed dormancy through glucose metabolism [30]. | |
| qLTGP5 | LOC_Os05g47840 | IPT7 | tRNA isopentenyltransferase family protein | It was upregulated during the recovery stage after cold stress in cold-tolerant cultivars [31]. |
| LOC_Os05g47890 | OsRACK1B | WD domain, G-beta repeat domain-containing protein | It may be involved in rice seed germination by regulating G protein to control hormone signaling response [32]. | |
| LOC_Os05g48020 | OsSYP71 | SNARE domain-containing protein | It was significantly upregulated under oxidative stress [33]. | |
| LOC_Os05g48390 | OsPHO2 | Ubiquitin-conjugating enzyme protein | After cold acclimation, arabidopsis pho2 mutants, with increased stem Pi, were more sensitive to freezing than the wild type [34]. | |
| LOC_Os05g48590 | IAA19 | OsIAA19—Auxin-responsive Aux/IAA gene family member | IAA19 is an auxin-related gene that is induced by cold stress [31]. | |
| qRAPL10 | LOC_Os10g35810 | thylakoid lumenal protein | It induced during the chilling and recovery treatment periods of 9311 and DC90 [35]. | |
| qPLR12 | LOC_Os12g38200 | OsDof29 | dof zinc finger domain-containing protein | It is upregulated after cold stress [36]. |
| LOC_Os12g38400 | OsMYB91 | MYB family transcription factor | It is a stress responsive gene that participates in the coordination of abiotic stress resistance [37]. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Zou, B.; Lu, C.; Song, G.; Gao, Q.; Wang, P.; Wu, G.; Jin, W.; Yin, H.; Cheng, Q.; et al. Unraveling the Genetic Architecture for Low Temperature Germinability-Related Traits in Rice Using Genome-Wide Association Study. Agronomy 2022, 12, 1194. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051194
Li C, Zou B, Lu C, Song G, Gao Q, Wang P, Wu G, Jin W, Yin H, Cheng Q, et al. Unraveling the Genetic Architecture for Low Temperature Germinability-Related Traits in Rice Using Genome-Wide Association Study. Agronomy. 2022; 12(5):1194. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051194
Chicago/Turabian StyleLi, Caijing, Baoli Zou, Changsheng Lu, Guiting Song, Qiang Gao, Peng Wang, Guangliang Wu, Wei Jin, Hui Yin, Qin Cheng, and et al. 2022. "Unraveling the Genetic Architecture for Low Temperature Germinability-Related Traits in Rice Using Genome-Wide Association Study" Agronomy 12, no. 5: 1194. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051194