
Macro- and Microscopic Characterization of Components of Resistance against Puccinia striiformis f. sp. tritici in a Collection of Spanish Bread Wheat Cultivars


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Pathogen Isolation
2.3. Screening of Bread Wheat Germplasm
2.3.1. Inoculation Assays for Evaluation of Disease Severity (DS) and Infection Type (IT)
2.3.2. Disease Assessment via DS and IT
2.4. Characterization of the Response to Yellow Rust
2.4.1. Inoculation Assays for Evaluation of Macroscopic and Microscopic Resistance Components
2.4.2. Assessment of Macroscopic Resistance Components
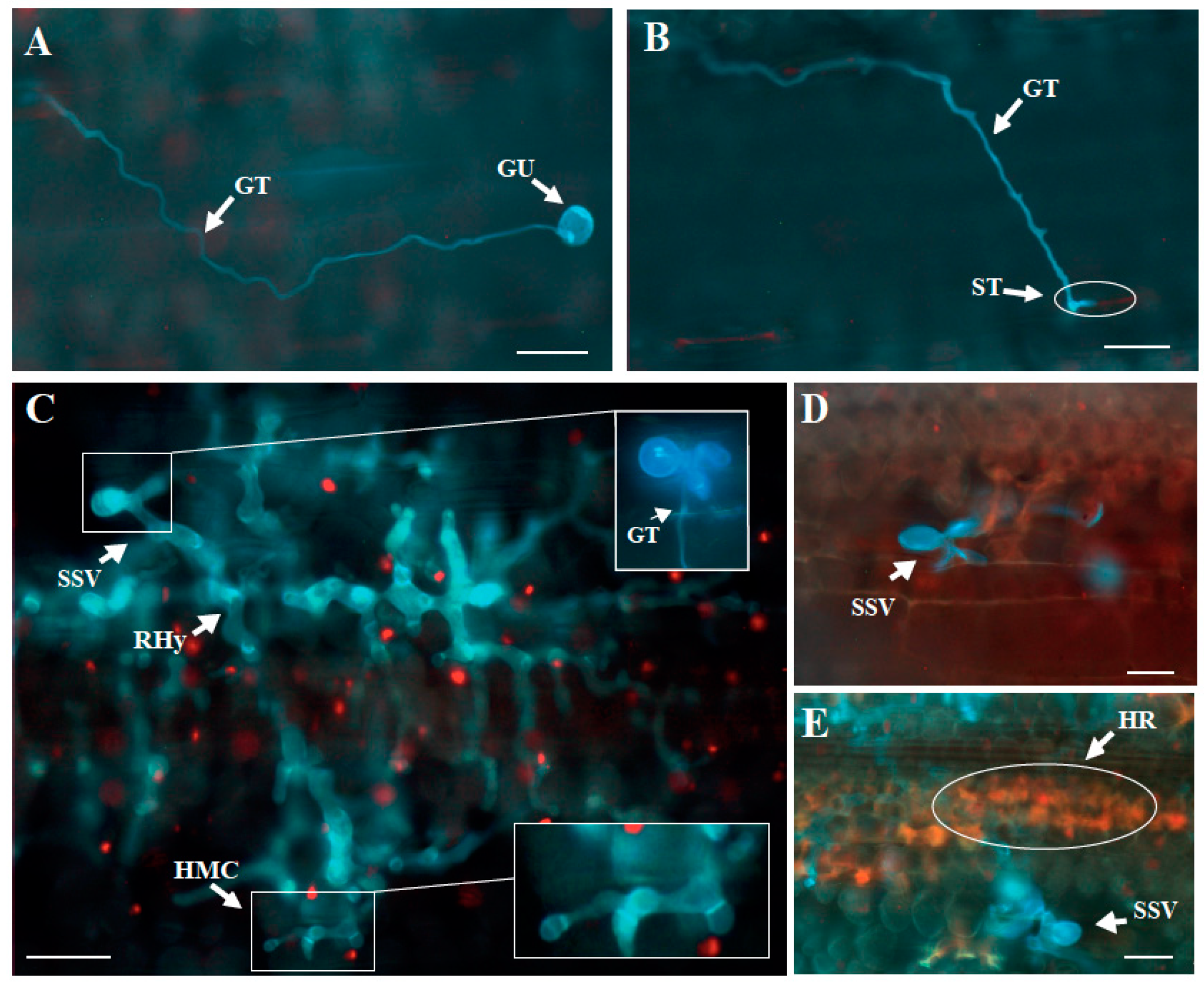
2.4.3. Assessment of Microscopic Resistance Components
2.5. Statistical Analysis
3. Results and Discussion
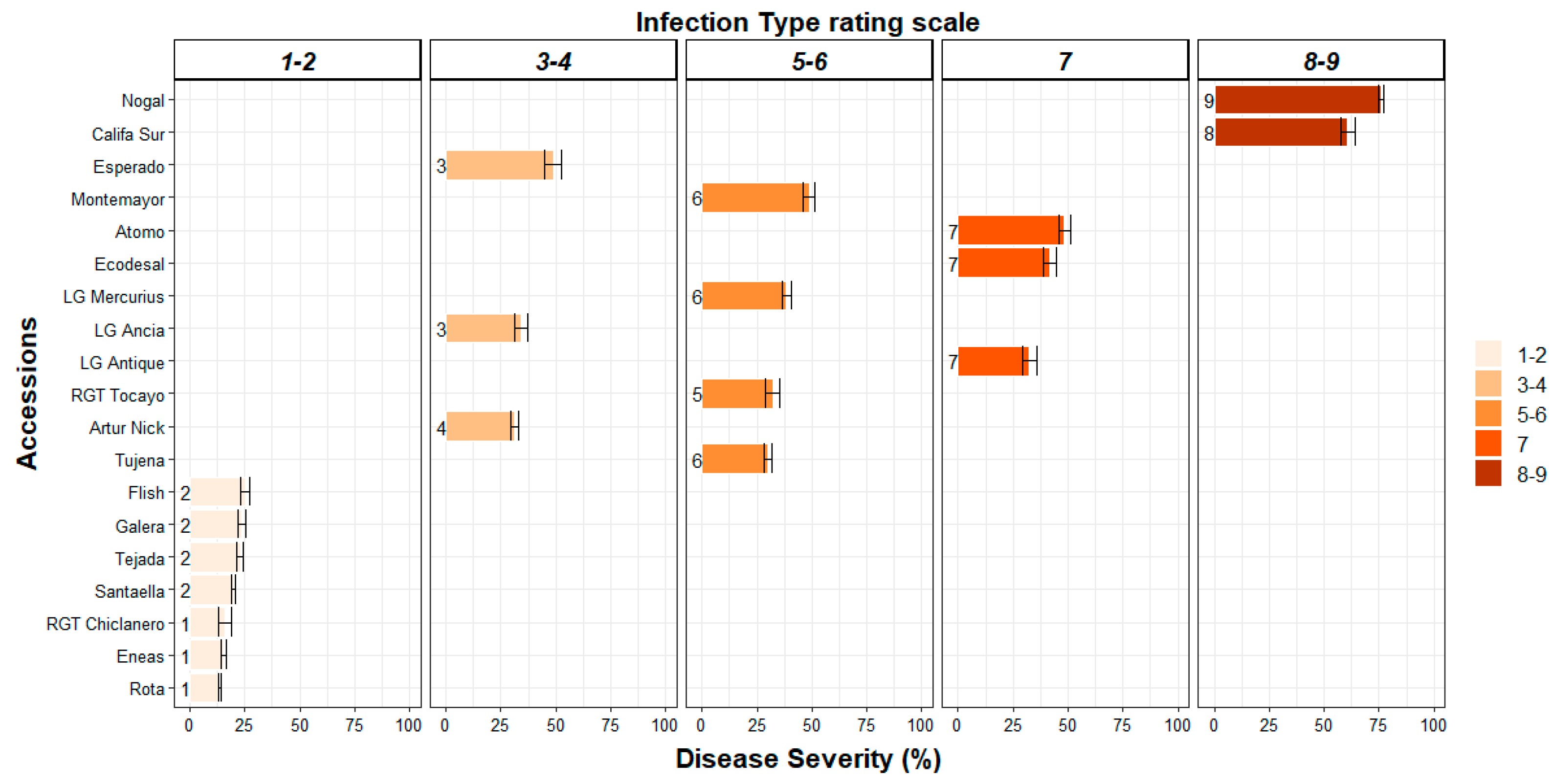
3.1. Resistance and Susceptibility Responses to P. striiformis Infection amongst Bread Wheat Accessions
3.2. Macroscopic Components of Resistance to P. striiformis Infection
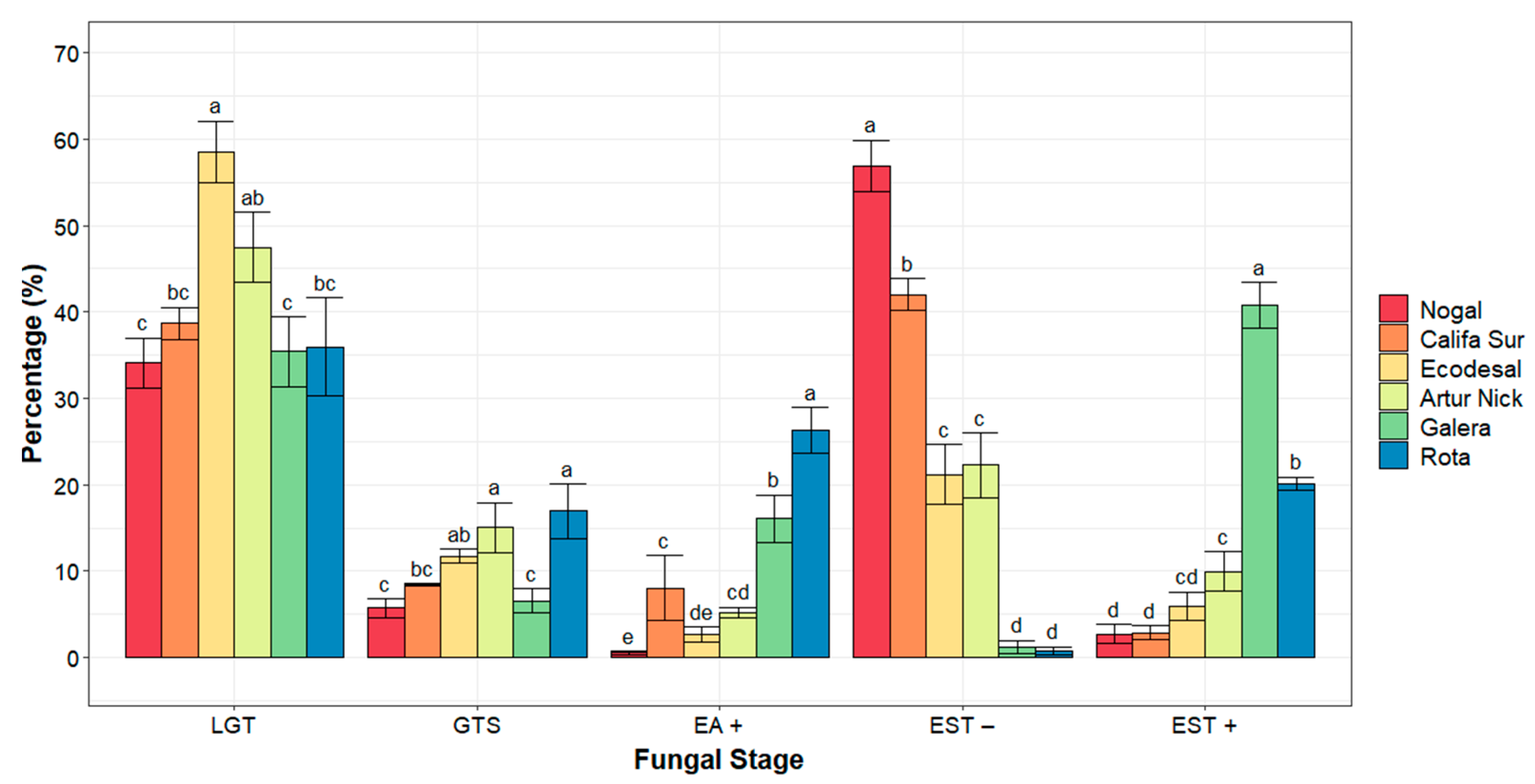
3.3. Microscopic Components of Resistance to P. striiformis Infection
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT Food and Agriculture Organization of the United Nations Statistical Database. Available online: https://www.fao.org/faostat/es/ (accessed on 1 April 2022).
- The Wheat Initiative. Available online: https://www.wheatinitiative.org/ (accessed on 1 April 2022).
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.G.; Lagudah, E.S.; Spielmeyer, W.; Dodds, P.N. The past, present and future of breeding rust resistant wheat. Front. Plant Sci. 2014, 5, 641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X. Pathogens which threaten food security: Puccinia striiformis, the wheat stripe rust pathogen. Food Secur. 2020, 12, 239–251. [Google Scholar] [CrossRef]
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Beddow, J.M.; Pardey, P.G.; Chai, Y.; Hurley, T.M.; Kriticos, D.J.; Braun, H.J.; Park, R.F.; Cuddy, W.S.; Yonow, T. Research investment implications of shifts in the global geography of wheat stripe rust. Nat. Plants 2015, 1, 15132. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wellings, C.; Chen, X.; Kang, Z.; Liu, T. Wheat stripe (yellow) rust caused by Puccinia striiformis f. sp. tritici. Mol. Plant Pathol. 2014, 15, 433–446. [Google Scholar] [CrossRef]
- Moldenhauer, J.; Moerschbacher, B.M.; Van Der Westhuizen, A.J. Histological investigation of stripe rust (Puccinia striiformis f.sp. tritici) development in resistant and susceptible wheat cultivars. Plant Pathol. 2006, 55, 469–474. [Google Scholar] [CrossRef]
- Hovmøller, M.S.; Sørensen, C.K.; Walter, S.; Justesen, A.F. Diversity of Puccinia striiformis on Cereals and Grasses. Annu. Rev. Phytopathol. 2011, 49, 197–217. [Google Scholar] [CrossRef]
- Bozkurt, T.O.; McGrann, G.R.D.; MacCormack, R.; Boyd, L.A.; Akkaya, M.S. Cellular and transcriptional responses of wheat during compatible and incompatible race-specific interactions with Puccinia striiformis f. sp. tritici. Mol. Plant Pathol. 2010, 11, 625–640. [Google Scholar] [CrossRef]
- Singh, R.P.; Huerta-Espino, J.; Roelfs, A.P. The wheat rusts. In Bread Wheat Improvement and Production; Curtis, B.C., Rajaram, S., Gómez, H., Eds.; FAO: Rome, Italy, 2002; ISBN 92-5-104809-6. [Google Scholar]
- Ali, S.; Rodriguez-Algaba, J.; Thach, T.; Sørensen, C.K.; Hansen, J.G.; Lassen, P.; Nazari, K.; Hodson, D.P.; Justesen, A.F.; Hovmøller, M.S. Yellow rust epidemics worldwide were caused by pathogen races from divergent genetic lineages. Front. Plant Sci. 2017, 8, 1057. [Google Scholar] [CrossRef] [Green Version]
- GrainGenes U.S. Department of Agriculture-Agricultural Research Service. Available online: https://wheat.pw.usda.gov/GG3/wgc (accessed on 1 April 2022).
- Jamil, S.; Shahzad, R.; Ahmad, S.; Fatima, R.; Zahid, R.; Anwar, M.; Iqbal, M.Z.; Wang, X. Role of Genetics, Genomics, and Breeding Approaches to Combat Stripe Rust of Wheat. Front. Nutr. 2020, 7, 173. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwessinger, B. Fundamental wheat stripe rust research in the 21st century. New Phytol. 2017, 213, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, X. Stripe Rust Resistance. In Stripe Rust; Chen, X., Kang, Z., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 353–558. [Google Scholar] [CrossRef]
- Saleem, K.; Arshad, H.M.I.; Shokat, S.; Atta, B.M. Appraisal of wheat germplasm for adult plant resistance against stripe rust. J. Plant Protect. Res. 2015, 55, 405–414. [Google Scholar] [CrossRef]
- Guo, Q.; Zhang, Z.J.; Xu, Y.B.; Li, G.H.; Feng, J.; Zhou, Y. Quantitative Trait Loci for High-Temperature Adult-Plant and Slow-Rusting Resistance to Puccinia striiformis f. sp. tritici in Wheat Cultivars. Phytopathology 2008, 98, 803–809. [Google Scholar] [CrossRef] [Green Version]
- Bueno-Sancho, V.; Persoons, A.; Hubbard, A.; Cabrera-Quio, L.E.; Lewis, C.M.; Corredor-Moreno, P.; Bunting, D.C.E.; Ali, S.; Chng, S.; Hodson, D.P.; et al. Pathogenomic Analysis of Wheat Yellow Rust Lineages Detects Seasonal Variation and Host Specificity. Genome Biol. Evol. 2017, 9, 3282–3296. [Google Scholar] [CrossRef] [Green Version]
- Hovmøller, M.S.; Walter, S.; Bayles, R.A.; Hubbard, A.; Flath, K.; Sommerfeldt, N.; Leconte, M.; Czembor, P.; Rodriguez-Algaba, J.; Thach, T.; et al. Replacement of the European wheat yellow rust population by new races from the centre of diversity in the near-Himalayan region. Plant Pathol. 2016, 65, 402–411. [Google Scholar] [CrossRef] [Green Version]
- Hubbard, A.; Lewis, C.M.; Yoshida, K.; Ramirez-Gonzalez, R.H.; de Vallavieille-Pope, C.; Thomas, J.; Kamoun, S.; Bayles, R.; Uauy, C.; Saunders, D.G.O. Field pathogenomics reveals the emergence of a diverse wheat yellow rust population. Genome Biol. 2015, 16, 23. [Google Scholar] [CrossRef] [Green Version]
- GRRC Global Rust Reference Center. Available online: https://agro.au.dk/forskning/internationale-platforme/wheatrust/ (accessed on 1 April 2022).
- EUROSTAT European Statistical Office. Available online: https://ec.europa.eu/eurostat/ (accessed on 1 April 2022).
- GENVCE Grupo Para la Evaluación de Nuevas Variedades de Cultivos Extensivos en España. Available online: https://genvce.org/ (accessed on 1 April 2022).
- Milus, E.A.; Kristensen, K.; Hovmøller, M.S. Evidence for increased aggressiveness in a recent widespread strain of Puccinia striiformis f. sp. tritici causing stripe rust of wheat. Phytopathology 2009, 99, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Moreno, F.; Solís, I. Wheat rust evolution in Spain: An historical review. Phytopathol. Mediterr. 2019, 58, 3–16. [Google Scholar] [CrossRef]
- Almacellas, J.; Álvaro, F. La roya amarilla del trigo: Características, situación actual y claves para el control. Gd. Cultiv. 2015, 5, 1–10. [Google Scholar]
- Singh, R.P.; Singh, P.K.; Rutkoski, J.; Hodson, D.P.; He, X.; Jørgensen, L.N.; Hovmøller, M.S.; Huerta-Espino, J. Disease Impact on Wheat Yield Potential and Prospects of Genetic Control. Annu. Rev. Phytopathol. 2016, 54, 303–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bock, C.H.; Barbedo, J.G.A.; Del Ponte, E.M.; Bohnenkamp, D.; Mahlein, A.K. From visual estimates to fully automated sensor-based measurements of plant disease severity: Status and challenges for improving accuracy. Phytopathol. Res. 2020, 2, 9. [Google Scholar] [CrossRef] [Green Version]
- Mutka, A.M.; Bart, R.S. Image-based phenotyping of plant disease symptoms. Front. Plant Sci. 2015, 5, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porras, R.; Pérez-De-Luque, A.; Sillero, J.C.; Miguel-Rojas, C. Behavior of spanish durum wheat genotypes against Zymoseptoria tritici: Resistance and susceptibility. Span. J. Agric. Res. 2021, 19, e1002. [Google Scholar] [CrossRef]
- Stewart, E.L.; McDonald, B.A. Measuring quantitative virulence in the wheat pathogen Zymoseptoria tritici using high-throughput automated image analysis. Phytopathology 2014, 104, 985–992. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Yu, K.; Pauls, K.P.; Navabi, A. Application of image analysis in studies of quantitative disease resistance, exemplified using common bacterial blight-common bean pathosystem. Phytopathology 2012, 102, 434–442. [Google Scholar] [CrossRef] [Green Version]
- Bock, C.H.; Parker, P.E.; Cook, A.Z.; Gottwald, T.R. Visual rating and the use of image analysis for assessing different symptoms of citrus canker on grapefruit leaves. Plant Dis. 2008, 92, 530–541. [Google Scholar] [CrossRef] [Green Version]
- Lamari, L. Assess 2.0 Image Analysis Software for Plant Disease Quantification; The American Phytopathological Society: St. Paul, MN, USA, 2002. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Gallego-Sánchez, L.M.; Canales, F.J.; Montilla-Bascón, G.; Prats, E. Rust: A robust, user-friendly script tool for rapid measurement of rust disease on cereal leaves. Plants 2020, 9, 1182. [Google Scholar] [CrossRef]
- Pethybridge, S.J.; Nelson, S.C. Leaf doctor: A new portable application for quantifying plant disease severity. Plant Dis. 2015, 99, 1310–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hovmøller, M.S.; Rodriguez-Algaba, J.; Thach, T.; Sørensen, C.K. Race Typing of Puccinia striiformis on Wheat. In Wheat Rust Diseases. Methods in Molecular Biology; Periyannan, S., Ed.; Humana Press: New York, NY, USA, 2017; Volume 1659, pp. 29–40. [Google Scholar] [CrossRef]
- Sørensen, C.K.; Hovmøller, M.S.; Leconte, M.; Dedryver, F.; De Vallavieille-Pope, C. New races of Puccinia striiformis found in Europe reveal race specificity of long-term effective adult plant resistance in wheat. Phytopathology 2014, 104, 1042–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sørensen, C.K.; Thach, T.; Hovmøller, M.S. Assessment of aggressiveness of Puccinia striiformis on wheat. In Wheat Rust Diseases. Methods in Molecular Biology; Periyannan, S., Ed.; Humana Press: New York, NY, USA, 2017; Volume 1659, pp. 41–47. [Google Scholar] [CrossRef]
- Zhao, Y.; Cheng, P.; Li, T.; Ma, J.; Zhang, Y.; Wang, H. Investigation of urediospore morphology, histopathology and epidemiological components on wheat plants infected with UV-B-induced mutant strains of Puccinia striiformis f. sp. tritici. Microbiologyopen 2019, 8, e870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moldenhauer, J.; Pretorius, Z.A.; Moerschbacher, B.M.; Prins, R.; Van Der Westhuizen, A.J. Histopathology and PR-protein markers provide insight into adult plant resistance to stripe rust of wheat. Mol. Plant Pathol. 2008, 9, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Soleiman, N.H.; Solis, I.; Sillero, J.C.; Herrera-Foessel, S.A.; Ammar, K.; Martinez, F. Evaluation of Macroscopic and Microscopic Components of Partial Resistance to Leaf Rust in Durum Wheat. J. Phytopathol. 2014, 162, 359–366. [Google Scholar] [CrossRef]
- Sørensen, C.K.; Labouriau, R.; Hovmøller, M.S. Temporal and spatial variability of fungal structures and host responses in an incompatible Rust-Wheat interaction. Front. Plant Sci. 2017, 8, 484. [Google Scholar] [CrossRef]
- Jagger, L.J.; Newell, C.; Berry, S.T.; Maccormack, R.; Boyd, L.A. Histopathology provides a phenotype by which to characterize stripe rust resistance genes in wheat. Plant Pathol. 2011, 60, 640–648. [Google Scholar] [CrossRef]
- McNeal, F.H.; Konzak, C.F.; Smith, E.P.; Tate, W.S.; Russell, T.S. A uniform system for recording and processing cereal research data. US Agric. Res. Serv. 1971, 42, 34–121. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.r-project.org/ (accessed on 1 April 2022).
- Martínez-Moreno, F.; Giraldo, P.; Nieto, C.; Ruiz, M. Resistance to Leaf and Yellow Rust in a Collection of Spanish Bread Wheat Landraces and Association with Ecogeographical Variables. Agronomy 2022, 12, 187. [Google Scholar] [CrossRef]
- Chen, X.M. Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Can. J. Plant Pathol. 2005, 27, 314–337. [Google Scholar] [CrossRef]
- El Jarroudi, M.; Kouadio, A.L.; Mackels, C.; Tychon, B.; Delfosse, P.; Bock, C.H. A comparison between visual estimates and image analysis measurements to determine septoria leaf blotch severity in winter wheat. Plant Pathol. 2015, 64, 355–364. [Google Scholar] [CrossRef]
- Bock, C.H.; Poole, G.H.; Parker, P.E.; Gottwald, T.R. Plant Disease Severity Estimated Visually, by Digital Photography and Image Analysis, and by Hyperspectral Imaging. Crit. Rev. Plant Sci. 2010, 29, 59–107. [Google Scholar] [CrossRef]
- Singh, R.; Huerta-Espino, J. Effect of leaf rust resistance gene Lr34 on components of slow rusting at seven growth stages in wheat. Euphytica 2003, 129, 371–376. [Google Scholar] [CrossRef]
- Wang, X.; Mccallum, B.D.; Fetch, T.; Bakkeren, G.; Marais, G.F.; Saville, B.J. Comparative microscopic and molecular analysis of Thatcher near-isogenic lines with wheat leaf rust resistance genes Lr2a, Lr3, LrB or Lr9 upon challenge with different Puccinia triticina races. Plant Pathol. 2013, 62, 698–707. [Google Scholar] [CrossRef]
- Wang, X.; McCallum, B.D.; Fetch, T.; Bakkeren, G.; Saville, B.J. Sr36- and Sr5-mediated resistance response to Puccinia graminis f. sp. tritici is associated with callose deposition in wheat guard cells. Phytopathology 2015, 105, 728–737. [Google Scholar] [CrossRef] [Green Version]
- de Vallavieille-Pope, C.; Huber, L.; Leconte, M.; Goyeau, H. Comparative effects of temperature and interrupted wet periods on germination, penetration, and infection of Puccinia recondita f. sp. tritici and P. striiformis on wheat seedlings. Phytopathology 1995, 85, 409–415. [Google Scholar] [CrossRef]
- de Vallavieille-Pope, C.; Huber, L.; Leconte, M.; Bethenod, O. Preinoculation effects of light quantity on infection efficiency of Puccinia striiformis and P. triticina on wheat seedlings. Phytopathology 2002, 92, 1308–1314. [Google Scholar] [CrossRef] [Green Version]
- Walters, D.R. Disguising the Leaf Surface: The Use of Leaf Coatings for Plant Disease Control. Eur. J. Plant Pathol. 2006, 114, 255–260. [Google Scholar] [CrossRef]
- Gupt, S.K.; Chand, R.; Mishra, V.K.; Ahirwar, R.N.; Bhatta, M.; Joshi, A.K. Spot blotch disease of wheat as influenced by foliar trichome and stomata density. J. Agric. Food Res. 2021, 6, 100227. [Google Scholar] [CrossRef]
- Imboden, L.; Afton, D.; Trail, F. Surface interactions of Fusarium graminearum on barley. Mol. Plant Pathol. 2018, 19, 1332–1342. [Google Scholar] [CrossRef] [Green Version]
- Rubiales, D.; Niks, R.E. Avoidance of rust infection by some genotypes of Hordeum chilense due to their relative inability to induce the formation of appressoria. Physiol. Mol. Plant Pathol. 1996, 49, 89–101. [Google Scholar] [CrossRef]
- Calo, L.; Garcia, I.; Gotor, C.; Romero, L.C. Leaf hairs influence phytopathogenic fungus infection and confer an increased resistance when expressing a Trichoderma-1,3-glucanase. J. Exp. Bot. 2006, 57, 3911–3920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleem, K.; Sørensen, C.K.; Labouriau, R.; Hovmøller, M.S. Spatiotemporal changes in fungal growth and host responses of six yellow rust resistant near-isogenic lines of wheat. Plant Pathol. 2019, 68, 1320–1330. [Google Scholar] [CrossRef]
- Kang, Z.S.; Wang, Y.; Huang, L.L.; Wei, G.R.; Zhao, J. Histology and ultrastructure of incompatible combination between Puccinia striiformis and wheat with low reaction type resistance. Agric. Sci. China 2003, 2, 1102–1113. [Google Scholar]
- Zhang, H.; Wang, C.; Cheng, Y.; Chen, X.; Han, Q.; Huang, L.; Wei, G.; Kang, Z. Histological and cytological characterization of adult plant resistance to wheat stripe rust. Plant Cell Rep. 2012, 31, 2121–2137. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Pustules/cm2 | Mean Pustule Size (cm2 × 10−3) | Total Pustule Area (%) | Total Affected Area (%) | Pustule Area/ Disease Area (%) |
|---|---|---|---|---|---|
| Nogal (IT 9) 2 | 230.85 ± 9.50 a | 0.49 ± 0.03 a | 11.14 ± 0.57 a | 1.64 (1.10 ± 0.25) c | 88.37 ± 4.10 a |
| Califa Sur (IT 8) | 205.27 ± 8.22 a | 0.48 ± 0.01 a | 9.91 ± 0.49 a | 5.98 (2.39 ± 0.20) b | 63.21 ± 4.58 b |
| Ecodesal (IT 7) | 153.44 ± 9.27 b | 0.46 ± 0.01 a | 7.00 ± 0.52 b | 2.92 (1.60 ± 0.23) c | 73.40 ± 5.34 b |
| Artur Nick (IT 4) | 42.27 ± 8.93 c | 0.39 ± 0.03 b | 1.70 ± 0.45 c | 24.02 (4.79 ± 0.39) a | 7.08 ± 1.71 c |
| Galera (IT 2) | - | - | - | 8.22 (2.75 ± 0.31) b | - |
| Rota (IT 1) | - | - | - | 1.51 (1.17 ± 0.14) c | - |
| Accession | LP0 (hours) | Final Lesion Length (mm) |
|---|---|---|
| Nogal | 264 ± 0.00 a | 60.58 ± 3.05 a |
| Califa Sur | 264 ± 0.00 a | 63.63 ± 2.47 a |
| Ecodesal | 303.75 ± 8.56 b | 42.89 ± 4.59 b |
| Artur Nick | 306 ± 10.88 b | 44.18 ± 6.34 b |
| Accession | Colony Length (µm) | % EST− | % EST+ |
|---|---|---|---|
| Nogal | 867.92 ± 34.31 a | 95.61 (0.20 ± 0.04) a | 4.39 (1.37 ± 0.04) c |
| Califa Sur | 822.26 ± 18.85 ab | 93.73 (0.25 ± 0.03) a | 6.27 (1.32 ± 0.03) c |
| Ecodesal | 646.61 ± 58.45 cd | 77.07 (0.49 ± 0.10) b | 22.93 (1.08 ± 0.10) b |
| Artur Nick | 537.86 ± 38.51 d | 68.42 (0.59 ± 0.09) b | 31.58 (0.98 ± 0.09) b |
| Galera | 739.33 ± 44.93 bc | 2.91 (1.43 ± 0.07) c | 97.09 (0.14 ± 0.07) a |
| Rota | 368.99 ± 19.33 e | 3.68 (1.42 ± 0.08) c | 96.32 (0.15 ± 0.08) a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porras, R.; Miguel-Rojas, C.; Pérez-de-Luque, A.; Sillero, J.C. Macro- and Microscopic Characterization of Components of Resistance against Puccinia striiformis f. sp. tritici in a Collection of Spanish Bread Wheat Cultivars. Agronomy 2022, 12, 1239. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051239
Porras R, Miguel-Rojas C, Pérez-de-Luque A, Sillero JC. Macro- and Microscopic Characterization of Components of Resistance against Puccinia striiformis f. sp. tritici in a Collection of Spanish Bread Wheat Cultivars. Agronomy. 2022; 12(5):1239. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051239
Chicago/Turabian StylePorras, Rafael, Cristina Miguel-Rojas, Alejandro Pérez-de-Luque, and Josefina C. Sillero. 2022. "Macro- and Microscopic Characterization of Components of Resistance against Puccinia striiformis f. sp. tritici in a Collection of Spanish Bread Wheat Cultivars" Agronomy 12, no. 5: 1239. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12051239