
The Production of Dual-Purpose Triticale in Arid Regions: Application of Organic and Inorganic Treatments under Water Deficit Conditions

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
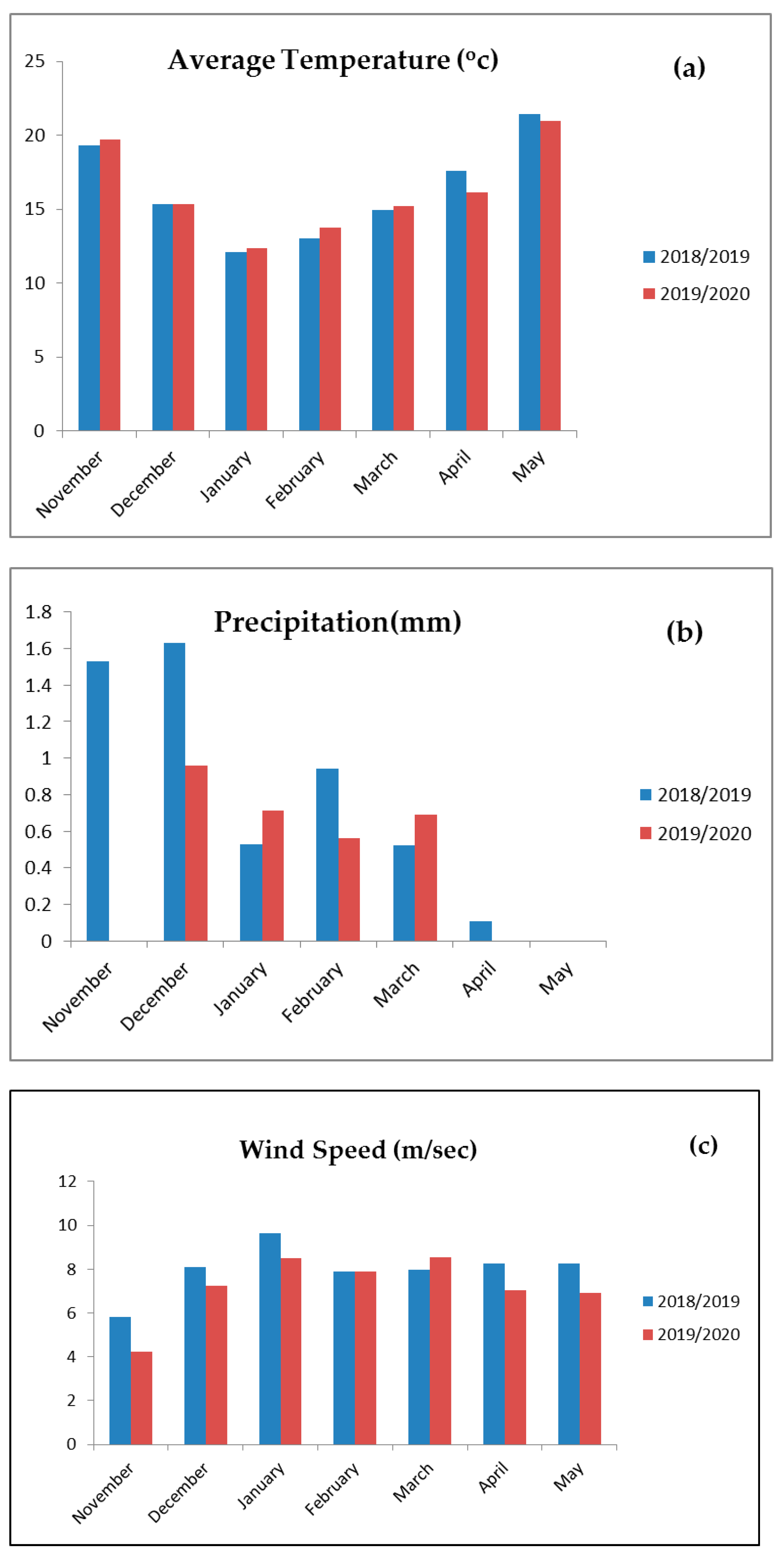
2.1. Experimental Location and Treatments
2.2. Forage Yield and Quality Characteristics
2.3. Agronomic Characteristics and Grain Yield
2.4. Experimental Design
2.5. Statistical Procedures
3. Results
3.1. Forage Yield and Quality
3.2. Grain yield and Agronomic Characteristics
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AA | Ascorbic acid |
| ADF | Acid detergent fiber |
| ADL | Acid detergent lignin |
| BY | Biological yield |
| AC | Age at cutting |
| CA | Crude ash |
| CP | Crude protein |
| DAS | Days after sowing |
| DOM | Degraded Organic matter |
| DNDF | Degraded neutral detergent fiber |
| DFY | Dry forage yield |
| 100 GW | 100-grain weight |
| GY | Grain yield |
| HA | Humic acid |
| NDF | Neutral detergent fiber |
| NSM−2 | Number of spikes m−2 |
| NFC | Non-fiber carbohydrates |
| PH | Plant height |
| PF | Potassium fertilization |
References
- Lamega, S.A.; Komainda, M.; Hoffmann, M.P.; Ayisi, K.K.; Odhiambo, J.J.O.; Isselstein, J. It depends on the rain: Smallholder farmers’ perceptions on the seasonality of feed gaps and how it affects livestock in semi-arid and arid regions in Southern Africa. Clim. Risk Manag. 2021, 34, 100362. [Google Scholar] [CrossRef]
- Shaalan, A.M.; Attia, M.A.; Salama, H.S.A. Grain yield and growth parameters of pearl millet genotypes grown in an arid environment under variable plant densities. Asian J. Plant Sci. 2021, 20, 52–60. [Google Scholar] [CrossRef]
- Beji, S. Yield and quality of dual-purpose barley and triticale in a semi-arid environment in Tunisia. Afr. J. Agric. Res. 2016, 11, 2730–2735. [Google Scholar] [CrossRef]
- Küçüközdemir, Ü.; Dumlu, B.; Yalçın, Z.; Karagöz, H. Determination of yield, quality and winter hardiness characteristics of some triticale (xTriticosecale Wittmack) genotypes in Pasinler and Erzincan locations. Ekin J. Crop Breed. Genet. 2019, 5, 74–83. [Google Scholar]
- FAOSTAT. Statistical Yearbook; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018. [Google Scholar]
- Oba, M.; Penner, G.B.; Whyte, T.D.; Wierenga, K. Effects of feeding triticale dried distillers grains plus soluble as a nitrogen source on productivity of lactating dairy cows. J Dairy Sci. 2010, 93, 2044–2052. [Google Scholar] [CrossRef]
- Bilgili, U.; Cifci, E.A.; Hanoglu, H.; Yagdi, K.; Acikgoz, E. Yield and quality of triticale forage. J. Food Agric. Environ. 2009, 7, 556–560. [Google Scholar]
- Salama, H.S.A.; Badry, H.H. Forage and grain yields of dual-purpose triticale as influenced by the integrated use of Azotobacter chroococcum and mineral nitrogen fertilizer. Ital. J. Agron. 2020, 16, 1719. [Google Scholar] [CrossRef]
- Arseniuk, E. Triticale Abiotic Stresses—An Overview; Springer International Publishing: Cham, Switzerland, 2015; pp. 69–81. [Google Scholar]
- Akram, N.A.; Shafiq, F.; Ashraf, M. Ascorbic Acid-A Potential Oxidant Scavenger and Its Role in Plant Development and Abiotic Stress Tolerance. Front. Plant Sci. 2017, 8, 613. [Google Scholar] [CrossRef]
- Godoy, F.; Olivos-Hernández, K.; Stange, C.; Handford, M. Abiotic Stress in Crop Species: Improving Tolerance by Applying Plant Metabolites. Plants 2021, 10, 186. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Nahar, K.; Hossain, M.D.; Mahmud, J.A.; Hossen, M.; Fujita, M. Potassium: A Vital Regulator of Plant Responses and Tolerance to Abiotic Stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef] [Green Version]
- Singh, G.; Manuja, S.; Parmar, D.K.; Singh, D. Response of rainfed barley (Hordeum vulgare L.) to phosphorus and potassium application. J. Pharmacogn. Phytochem. 2021, 10, 468–471. [Google Scholar] [CrossRef]
- Ahmad, M.; Waheed, A.; Niaz, A.; Hannan, A.; Ranjha, A.M. Maize fodder quality characteristics and yield as affected by potassium application on calcareous sandy clay loam soil. Soil Environ. 2009, 28, 169–173. [Google Scholar]
- Kubar, G.M.; Talpur, K.H.; Kandhro, M.N.; Khashkhali, S.; Nizamani, M.M.; Kubar, M.S.; Kubar, K.A.; Kubar, A.A. Effect of potassium (K+) on growth, yield components and macronutrient accumulation in Wheat crop. Pure Appl. Biol. 2019, 8, 248–255. [Google Scholar] [CrossRef]
- Prajapati, K.; Modi, H.A. The Importance of Potassium in Plant Growth—A Review. Indian J. Plant Sci. 2012, 1, 177–186. [Google Scholar]
- Osman, E.A.M.; Nor Eldein, G.A. Response of Three Bread Wheat to Nitrogen Fertilizer with or without Ascorbic Acid Grown on a Clay Soil. J. Soil Sci. Agric. Eng. Mansoura Univ. 2017, 8, 267–274. [Google Scholar] [CrossRef]
- Salama, K.H.A. Amelioration of NaCl-induced alterations on the plasma membrane of Allium cepa L. by ascorbic acid. Aust. J Basic Appl. Sci. 2009, 3, 990–994. [Google Scholar]
- Shah, T.; Latif, S.; Khan, H.; Munsif, F.; Nie, L. Ascorbic Acid Priming Enhances Seed Germination and Seedling Growth of Winter Wheat under Low Temperature Due to Late Sowing in Pakistan. Agronomy 2019, 9, 757. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.; Chauhan, A.; Kumar, R.; Joshi, D.; Soni, P.G.; Meena, V.K. Dual purpose barley as affected by date of sowing, varieties and stage of harvesting-A review. Agric. Rev. 2017, 38, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Ronga, D.; Dal Prà, A.; Immovilli, A.; Ruozzi, F.; Davolio, R.; Pacchioli, M.T. Effects of Harvest Time on the Yield and Quality of Winter Wheat Hay Produced in Northern Italy. Agronomy 2020, 10, 917. [Google Scholar] [CrossRef]
- Salama, H.S.A.; Safwat, A.M.; Elghalid, O.H.; Abd El-Hady, A.M. Agronomic and In Vitro Quality Evaluation of Dual-Purpose Cereals Clipped at Variable Ages and Their Utilization in Rabbit Feeding. Agronomy 2021, 11, 1147. [Google Scholar] [CrossRef]
- Salama, H.S.A. Dual Purpose Barley Production in the Mediterranean Climate: Effect of Seeding Rate and Age at Forage Cutting. Int. J. Plant Prod. 2019, 13, 285–295. [Google Scholar] [CrossRef]
- Coblentz, W.K.; Akins, M.S.; Kalscheur, K.F.; Brink, G.E.; Cavadini, J.S. Effects of growth stage and growing degree day accumulations on triticale forages: 1. Dry matter yield, nutritive value, and in vitro dry matter disappearance. J. Dairy Sci. 2018, 101, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Gerke, J. Concepts and Misconceptions of Humic Substances as the Stable Part of Soil Organic Matter: A Review. Agronomy 2018, 8, 76. [Google Scholar] [CrossRef] [Green Version]
- Fahmi, A.H.; Sallume, M.O.; Aswad, A.H.; Abdulrahman, A.L.; Hamdi, G.J.; Abood, M.A. Interaction effect of potassium fertilizer, humic acid and irrigation intervals on growth and yield of wheat. Res. Crops 2020, 21, 31–35. [Google Scholar] [CrossRef]
- Bijanzadeh, E.; Emam, Y.; Perssarakli, M. Biochemical responses of water-stressed triticale (X Triticosecale wittmack) to humic acid and jasmonic acid. J. Plant Nutr. 2020, 44, 252–269. [Google Scholar] [CrossRef]
- Daur, I.; Bakhashwain, A.A. Effect of humic acid on growth and quality of maize fodder production. Pak. J. Bot. 2013, 45, 21–25. [Google Scholar]
- AOAC. International Official Methods of Analysis, 19th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2012. [Google Scholar]
- Van Soest, P.V.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Onodera, R.; Henderson, C. Growth factors of bacterial origin for the culture of the rumen oligotrich protozoon, Entodinium caudatum. J. Appl. Bacteriol. 1980, 48, 125–134. [Google Scholar] [CrossRef]
- Blümmel, M.; Steingaβ, H.; Becker, K. The relationship between in vitro gas production, in vitro microbial biomass yield and 15N incorporation and its implications for the prediction of voluntary feed intake of roughages. Br. J. Nutr. 1997, 77, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Box, G.E.P.; Wilson, K.B. On the experimental attainment of optimum conditions. J. R. Stat. Soc. 1951, 13, 1–45. [Google Scholar] [CrossRef]
- Peterson, G.R. Design and Analysis of Experiments; Marcel Dekker, Inc.: New York, NY, USA, 1985; pp. 253–301. [Google Scholar]
- StatSoft, I. STATISTICA (Data Analysis Software System), Version 7; StatSoft Inc.: Tulsa, OK, USA, 2012; p. 150. [Google Scholar]
- Hyams, D. Curve Expert Version 1.37. A Comprehensive Curve Fitting Package for Windows. 2005. [Google Scholar]
- Kaur, G.; Singh, A.; Aulakh, C.S.; Gill, J.S. Variation in Forage Yield and Quality Traits of Dual Purpose Barley under Different Agronomic Practices. Forage Res. 2013, 39, 42–44. [Google Scholar]
- Geren, H. Dry matter yield and silage quality of some winter cereals harvested at different stages under Mediterranean climate condition. Turk. J. Field Crops 2014, 19, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Azim, A.; Naseer, Z.; Ali, A. Nutritional evaluation of maize fodder at two different vegetative stages. Asian-Australas. J. Anim. Sci. 1989, 2, 27–34. [Google Scholar] [CrossRef]
- Buxton, D.R.; Fales, S.L. Plant-environment and quality. In Forage Quality, Evaluation, and Utilization; Fahey, G., Ed.; CSSA: Madison, WI, USA, 1994; pp. 155–199. [Google Scholar]
- Midha, L.K.; Panwar, K.S.; Sharma, N.K. Effect of cutting frequency and time of Nitrogen Application on yield & Quality of Oat. Forage Res. 1994, 25, 99–102. [Google Scholar]
- Rodrigo, S.; Poblaciones, M.J.; Pinheiro, N.; Maças, B.; Olea, L.; Santamaría, O.; García-White, T.; Coutinho, J. Dry matter yield and quality evaluation at different growth stages of triticale forage in south-west of the Iberian Peninsula. In The Contributions of Grasslands to the Conservation of Mediterranean Biodiversity; Porqu Eddu, C., Ríos, S., Eds.; CIHEAM/CIBIO/FAO/SEEP: Zaragoza, Spain, 2010; pp. i–iv. [Google Scholar]
- Chattha, M.U.; Sana, M.A.; Munir, H.; Ashraf, U.; Ul-Haq, I.; Zamir, S.I. Exogenous application of plant growth promoting substances enhances the growth, yield and quality of maize (Zea mays L.). Plant Knowl. J. 2015, 4, 1–6. [Google Scholar]
- Waraich, E.A.; Ahmad, R.; Halim, A.; Aziz, T. Alleviation of temperature stress by nutrient management 615 in crop plants: A review. J. Soil Sci. Plant Nutr. 2012, 12, 221–244. [Google Scholar] [CrossRef] [Green Version]
- Arrigoni, O.; De Tullio, M.C. Ascorbic acid: Much more than just an antioxidant. Biochim. Et Biophys. Acta 2002, 1569, 1–9. [Google Scholar] [CrossRef]
- Morad, K.A.; El-shafey, A.; ElMantany, R. Effect of Humic Acid Application on Growth and Productivity of Sunflower Under Saline Soil Conditions. J. Plant Prod. 2021, 11, 1193–1200. [Google Scholar] [CrossRef]
- Du, S.; Xu, M.; Yao, J. Relationship between fiber degradation kinetics and chemical composition of forages and by-products in ruminants. J. Appl. Anim. Res. 2016, 44, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Salama, H.S.A.; Shaalan, A.M.; Nasser, M.E.A. Forage performance of pearl millet (Pennisetum glaucum [L.] R. Br.) in arid regions: Yield and quality assessment of new genotypes on different sowing dates. Chil. J. Agric. Res. 2020, 80, 572–584. [Google Scholar] [CrossRef]
- Collar, C.; Aksland, G. Harvest Stage Effects on Yield and Quality of Winter Forage. Available online: https://alfalfa.ucdavis.edu/+symposium/proceedings/2001,01-133 (accessed on 10 March 2022).
- Hadjichristodoulou, A. Dual purpose barley. Rachis 1991, 10, 13–14. [Google Scholar]
- Winter, S.R.; Thompson, E.K. Grazing winter wheat: Response of semi dwarf cultivars to grain and grazed production systems. Agron. J. 1990, 82, 33–37. [Google Scholar] [CrossRef]
- Brennan, R.F.; Jayasena, K.W. Increasing applications of potassium fertilizer to barley crops grown on deficient sandy soils increased grain yields while decreasing some foliar diseases. Aust. J. Agric. Res. 2007, 58, 680–689. [Google Scholar] [CrossRef]
- Adnan, M.; Shah, Z.; Ullah, H.; Khan, B.; Arshad, M.; Mian, I.A.; Khan, G.A.; Alam, M.; Basir, A.; Rahman, I.; et al. Yield response of wheat to nitrogen and potassium fertilization. Pure Appl. Biol. 2016, 5, 868–875. [Google Scholar] [CrossRef]
- Wulgo, U.K.; Al-Solaimani, S.G.M.; Alghabari, F.M. Grain Sorghum Yield and Yield Components Influenced by the Effect of Potassium Fertilizer and Saline Irrigation Water under Arid Land Conditions. Int. J. Eng. Res. Technol. 2019, 8, 649–654. [Google Scholar]
- Ali, I.; Khan, A.A.; Munsif, F.; He, L.; Khan, A.; Ullah, S.; Saeed, W.; Iqbal, A.; Adnan, M.; Ligeng, J. Optimizing rates and application time of potassium fertilizer for improving growth, grain nutrients content and yield of wheat crop. Open Agric. 2019, 4, 500–508. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Z.; Liang, Y.; Han, Y.; Han, Y.; Tan, J. High Potassium Application Rate Increased Grain Yield of Shading-Stressed Winter Wheat by Improving Photosynthesis and Photosynthate Translocation. Front. Plant Sci. 2020, 11, 134. [Google Scholar] [CrossRef]
- Tóth, S.Z.; Puthur, J.T.; Nagy, V.; Garab, G. Experimental evidence for ascorbate dependent electron transport in leaves with inactive oxygen-evolving complexes. Plant Physiol. 2009, 149, 1568–1578. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, Q.; Huang, B. Ascorbic acid mitigation of water stress-inhibition of root growth in association with oxidative defense in tall fescue (Festuca arundinacea Schreb.). Front. Plant Sci. 2015, 6, 807. [Google Scholar] [CrossRef] [Green Version]
- Noreen, S.; Sultan, M.; Akhter, M.S.; Shah, K.H.; Ummara, U.; Manzoor, H.; Ulfat, M.; Alyemeni, M.N.; Ahmad, P. Foliar fertigation of ascorbic acid and zinc improves growth, antioxidant enzyme activity and harvest index in barley (Hordeum vulgare L.) grown under salt stress. Plant Physiol. Biochem. 2021, 158, 244–254. [Google Scholar] [CrossRef]
- Bakry, A.B.; Abdelraouf, R.E.; Ahmad, M.A. Effect of drought stress and ascorbic acid foliar application on productivity and irrigation water use efficiency of wheat under newly reclaimed sandy soil. Elixir Agric. 2013, 57A, 14394–14403. [Google Scholar]
- Ali, A.Y.A.; Ibrahim, M.H.; Zhoua, G.; Zhua, G.; Elsiddiga, A.M.I.; Suliman, M.S.E.; Elradi, S.B.M.; Salah, E.G.I. Interactive Impacts of Soil Salinity and Jasmonic Acid and Humic Acid on Growth Parameters, Forage Yield and Photosynthesis Parameters of Sorghum Plants. S. Afr. J. Bot. 2022, 146, 293–303. [Google Scholar] [CrossRef]
- Anwar, S.; Iqbal, F.; Khattak, W.A.; Islam, M.; Iqbal, B.; Khan, S. Response of Wheat Crop to Humic Acid and Nitrogen Levels. EC Agric. 2016, 3, 558–565. [Google Scholar]
- Anwar, S.; Khan, I.; Hussain, S.; Anjum, M.M.; Iqbal, B.; Hussain, A.; Iqbal, M.O.; Ali, N. Wheat response to different levels of humic acid and brassinolide. Pure Appl. Biol. 2016, 5, 782–787. [Google Scholar] [CrossRef]
- Inamullah; Ali, N. Assessment of various humic acid and sulfur levels for higher yields in wheat (Triticum aestivum L.). Sarhad J. Agric. 2014, 30, 47–52. [Google Scholar]
- Shahryari, R.; Mollasadeghi, V. Increasing of Wheat Grain Yield by Use of a Humic Fertilizer. Adv. Environ. Biol. 2011, 5, 516–518. Available online: https://www.researchgate.net/publication/288392719 (accessed on 10 March 2022).
- Wali, A.M.; Shamseldin, A.; Radwan, F.I.; Abd El Lateef, E.M.; Zaki, N.M. Response of Barley (Hordeum vulgare) Cultivars to Humic Acid, Mineral and Biofertilization under Calcareous Soil Conditions. Middle East J. Agric. Res. 2018, 7, 71–82. [Google Scholar]
- Dulaimy, J.A.M.A.; El-Fahdawi, W.A.T. Effect of Humic Acid on Growth and Yield of Barley Humic Acid as Interacted with Row Spacing. Indian J. Ecol. 2020, 47, 62–65. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Properties | Values | ||
|---|---|---|---|
| 1st Season (2018/19 Season) | 2nd Season (2019/20 Season) | ||
| A-Physical Analysis: | |||
| Soil Type | Sand 79.4% | Clay 15.4% | Silt 5.2% |
| Sand Loamy | |||
| B-Chemical Analysis: | |||
| pH | 7.96 | 8.18 | |
| E.C. (ds/m) | 0.80 | 2.24 | |
| Organic matter (%) | 1.01 | 0.533 | |
| Total N (%) | 3.05 | 3.07 | |
| HCO3− (ppm) | 4.0 | 5.6 | |
| Cl− (ppm) | 4.0 | 14.3 | |
| SO4− (meq/L) | 3.6 | 4.7 | |
| Ca++ (meq/L) | 7.0 | 3.9 | |
| Mg++ (meq/L) | 1.0 | 3.6 | |
| Na+ (meq/L) | 5.4 | 16.6 | |
| K+ (meq/L) | 0.8 | 0.5 | |
| Coded Level Factors | Unit | −S α (−2) | −F (−1) | Central Point (0) | +F (+1) | +S +α (2) | |
|---|---|---|---|---|---|---|---|
| Potassium fertilization (PF) | (X1) | kg ha−1 | 0 | 43.2 | 86.4 | 129.6 | 172.8 |
| Ascorbic acid (AA) | (X2) | mg L−1 | 0 | 25 | 50 | 75 | 100 |
| Age at cutting (AC) | (X3) | DAS | 40 | 65 | 90 | 115 | 140 |
| Humic acid (HA) | (X4) | kg ha−1 | 0 | 2.4 | 4.8 | 7.2 | 9.6 |
| Coded Values | Actual Levels | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Exp. No. | X1 | X2 | X3 | X4 | PF | AA | AC | HA | |
| Central points = 2 × 4 = 8 | 1 | 0 | 0 | 0 | 0 | 86.4 | 50 | 90 | 4.8 |
| 2 | 0 | 0 | 0 | 0 | 86.4 | 50 | 90 | 4.8 | |
| 3 | 0 | 0 | 0 | 0 | 86.4 | 50 | 90 | 4.8 | |
| 4 | 0 | 0 | 0 | 0 | 86.4 | 50 | 90 | 4.8 | |
| 5 | 0 | 0 | 0 | 0 | 86.4 | 50 | 90 | 4.8 | |
| 6 | 0 | 0 | 0 | 0 | 86.4 | 50 | 90 | 4.8 | |
| 7 | 0 | 0 | 0 | 0 | 86.4 | 50 | 90 | 4.8 | |
| 8 | 0 | 0 | 0 | 0 | 86.4 | 50 | 90 | 4.8 | |
| Star points = 2 × 4 = 8 | 1 | −2 | 0 | 0 | 0 | 0 | 50 | 90 | 4.8 |
| 2 | +2 | 0 | 0 | 0 | 172.8 | 50 | 90 | 4.8 | |
| 3 | 0 | −2 | 0 | 0 | 86.4 | 0 | 90 | 4.8 | |
| 4 | 0 | +2 | 0 | 0 | 86.4 | 100 | 90 | 4.8 | |
| 5 | 0 | 0 | −2 | 0 | 86.4 | 50 | 40 | 4.8 | |
| 6 | 0 | 0 | +2 | 0 | 86.4 | 50 | 140 | 4.8 | |
| 7 | 0 | 0 | 0 | −2 | 86.4 | 50 | 90 | 0 | |
| 8 | 0 | 0 | 0 | +2 | 86.4 | 50 | 90 | 9.6 | |
| Factorial points = 24 (one replicate) = 16 | 1 | −1 | −1 | −1 | −1 | 34.2 | 25 | 65 | 2.4 |
| 2 | −1 | −1 | −1 | 1 | 43.2 | 25 | 65 | 7.2 | |
| 3 | −1 | −1 | 1 | −1 | 43.2 | 25 | 115 | 2.4 | |
| 4 | −1 | −1 | 1 | 1 | 43.2 | 25 | 115 | 7.2 | |
| 5 | −1 | 1 | −1 | −1 | 43.2 | 75 | 65 | 2.4 | |
| 6 | −1 | 1 | −1 | 1 | 43.2 | 75 | 65 | 7.2 | |
| 7 | −1 | 1 | 1 | −1 | 43.2 | 75 | 115 | 2.4 | |
| 8 | −1 | 1 | 1 | 1 | 43.2 | 75 | 115 | 7.2 | |
| 9 | 1 | −1 | −1 | −1 | 129.6 | 25 | 65 | 2.4 | |
| 10 | 1 | −1 | −1 | 1 | 129.6 | 25 | 65 | 7.2 | |
| 11 | 1 | −1 | 1 | −1 | 129.6 | 25 | 115 | 2.4 | |
| 12 | 1 | −1 | 1 | 1 | 129.6 | 25 | 115 | 7.2 | |
| 13 | 1 | 1 | −1 | −1 | 129.6 | 75 | 65 | 2.4 | |
| 14 | 1 | 1 | −1 | 1 | 129.6 | 75 | 65 | 7.2 | |
| 15 | 1 | 1 | 1 | −1 | 129.6 | 75 | 115 | 2.4 | |
| 16 | 1 | 1 | 1 | 1 | 129.6 | 75 | 115 | 7.2 | |
| Charact. | Seasons | Equation | R2 | C.V % |
|---|---|---|---|---|
| DFY (t ha−1) | 2018/19 | Y = 4.35 + 0.78 X3. | 0.95 | 3.48 |
| 2019/20 | Y = 3.29 + 0.51 X3. | 0.97 | 3.07 | |
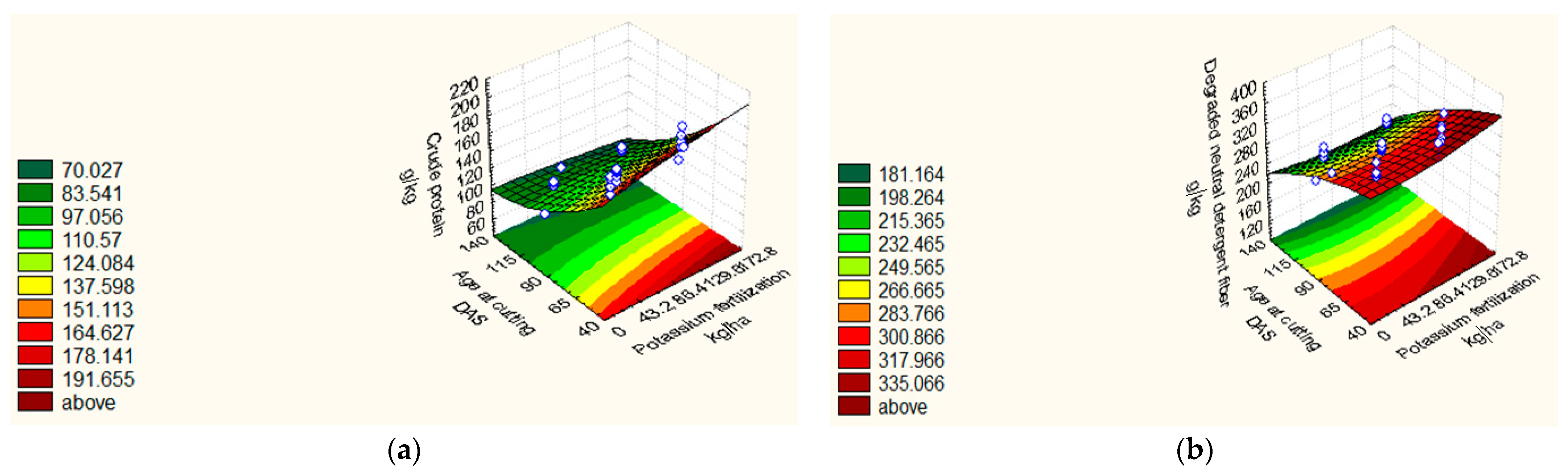
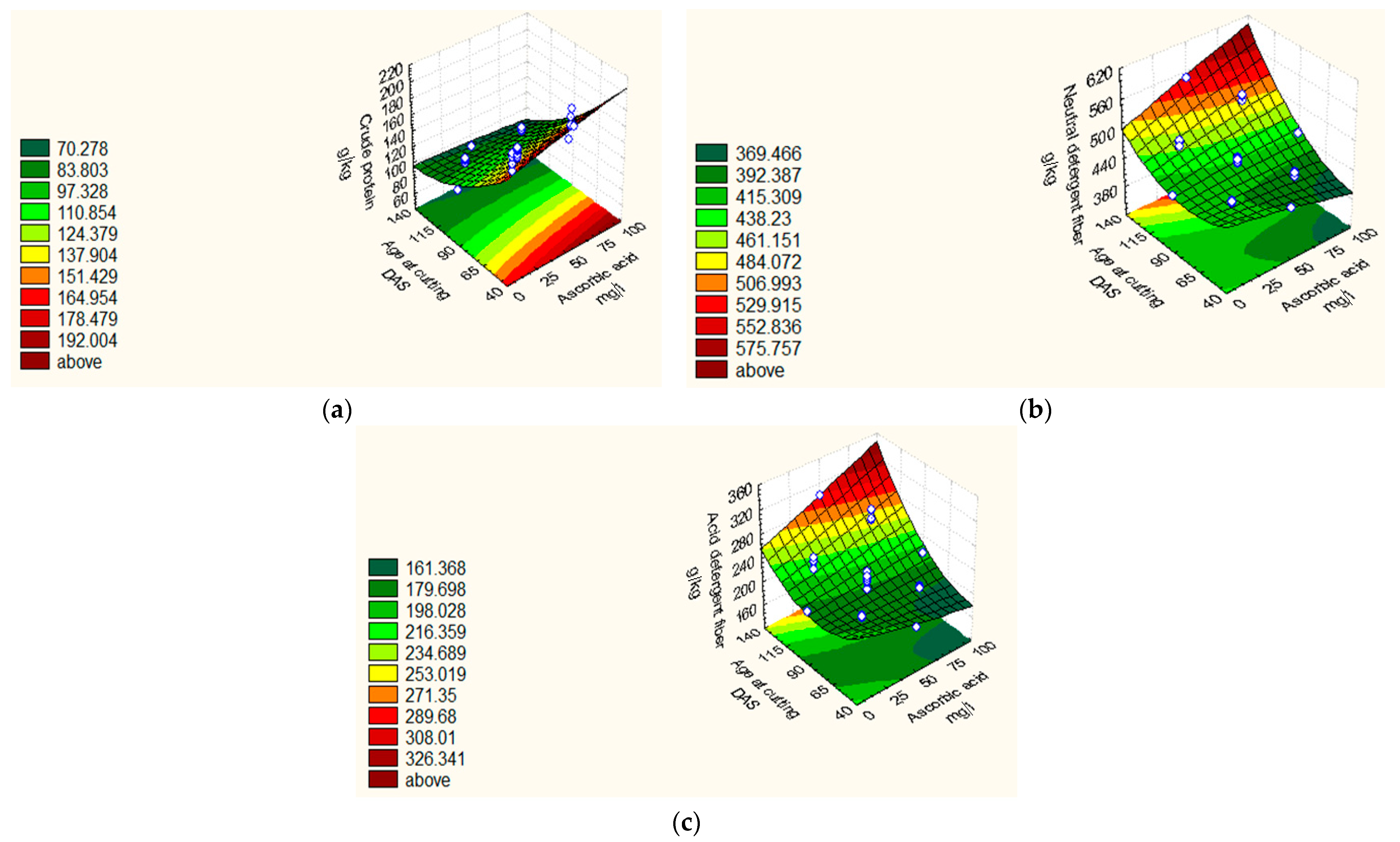
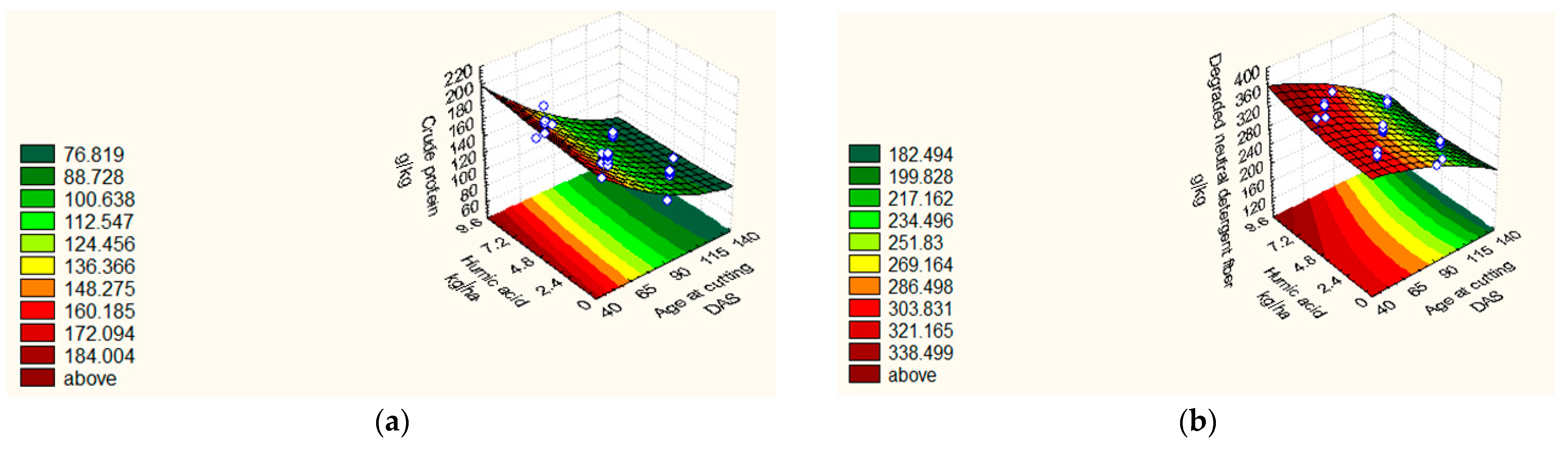
| CP (g kg−1) | 2018/19 | Y = 105.76 + 3.89 X1 +3.89 X2 − 29.84 X3 + 4.83 X32 + 1.75 X4 − 3.66 X1X3 − 3.67 X2X3 − 1.39 X3X4. | 0.93 | 1.42 |
| 2019/20 | Y = 107.01 + 3.44 X12 +2.91 X2 + 4.57 X22 − 24.91X3 + 8.36 X32 + 1.80 X4 + 5.30 X42. | 0.95 | 1.91 | |
| NDF (g kg−1) | 2018/19 | Y = 413.91 + 3.13 X12 + 6.11 X2 + 37.98 X3 + 12.65 X32 + 12.52 X2X3 + 3.22 X2X4. | 0.97 | 1.21 |
| 2019/20 | Y = 485.21 + 2.83 X1 + 5.79X12 − 5.21X22 + 40.89X3 + 4.99X4. | 0.94 | 1.05 | |
| ADF (g kg−1) | 2018/19 | Y = 183.68 + 5.43X12 + 5.94 X2 +29.17 X3 + 11.55 X32 + 10.61 X2X3. | 0.94 | 4.42 |
| 2019/20 | Y = 228.73 + 4.28X1 + 2.40 X2 − 3.08X22 + 32.02X3 − 2.23 X32. | 0.95 | 2.04 | |
| ADL (g kg−1) | 2018/19 | Y = 27.29 + 2.47 X12 + 0.80 X22 + 11.84 X3 + 5.55 X32 + 0.92 X4 + 2.22 X2X4. | 0.95 | 7.41 |
| 2019/20 | Y = 45.65 + 10.32 X3. | 0.92 | 5.33 | |
| NFC (g kg−1) | 2018/19 | Y = 303.52 − 4.58 X1 − 5.25 X2 + 4.62 X3 + 1.58 X32 − 4.28X4. | 0.64 | 1.18 |
| 2019/20 | Y = 272.52 + 3.01X22 + 5.02X3 + 6.94X32. | 0.53 | 1.97 | |
| DOM (g kg−1) | 2018/19 | Y = 553.87 − 77.50 X3 − 23.53 X32. | 0.93 | 1.83 |
| 2019/20 | Y = 455.12 + 6.82X22 − 74.08X3. | 0.89 | 3.14 | |
| DNDF (g kg−1) | 2018/19 | Y = 286.25 − 40.79 X3 − 6.43 X2X4. | 0.89 | 3.53 |
| 2019/20 | Y = 265.00 + 2.74 X1 + 2.510 X12 + 2.04X2 − 39.62 X3 − 5.11 X32 + 5.87 X4 + 2.76 X42 − 3.93 X1X2 − 3.18 X1X3 − 2.31 X3X4. | 0.80 | 1.42 |
| Treatments | DFY (t ha−1) | CP (g kg−1) | NFC (g kg−1) | ||||
|---|---|---|---|---|---|---|---|
| 2018/19 | 2019/20 | 2018/19 | 2019/20 | 2018/19 | 2019/20 | ||
| Central points | |||||||
| C1–C8 | 4.35 | 3.28 | 105.75 | 107.01 | 303.52 | 272.15 | |
| Star points | |||||||
| S1 | 4.57 | 3.01 | 92.10 | 112.00 | 305.79 | 277.45 | |
| S2 | 4.45 | 3.22 | 94.24 | 114.50 | 308.12 | 272.56 | |
| S3 | 4.57 | 3.19 | 90.50 | 115.00 | 309.37 | 269.11 | |
| S4 | 4.45 | 3.24 | 96.42 | 120.50 | 304.96 | 274.32 | |
| S5 | 2.94 | 2.30 | 173.15 | 177.20 | 293.23 | 257.68 | |
| S6 | 5.50 | 4.22 | 55.96 | 88.64 | 329.76 | 292.09 | |
| S7 | 4.13 | 3.15 | 96.50 | 117.62 | 301.45 | 273.82 | |
| S8 | 4.20 | 3.23 | 98.00 | 123.70 | 306.80 | 267.03 | |
| Factorial points | |||||||
| F1 | 1 | 3.63 | 2.54 | 132.23 | 152.24 | 301.21 | 258.20 |
| F2 | HA | 3.30 | 2.65 | 132.30 | 159.10 | 308.31 | 258.95 |
| F3 | AC | 5.00 | 3.88 | 80.71 | 94.17 | 317.58 | 281.74 |
| F4 | HA*AC | 5.05 | 3.90 | 83.59 | 102.31 | 325.28 | 275.98 |
| F5 | AA | 3.38 | 2.72 | 143.23 | 160.90 | 309.95 | 259.32 |
| F6 | AA*HA | 3.47 | 2.84 | 146.50 | 166.00 | 300.11 | 261.83 |
| F7 | AA*AC | 5.31 | 3.70 | 84.55 | 107.37 | 321.62 | 278.91 |
| F8 | AA*AC*HA | 5.18 | 3.75 | 84.81 | 107.60 | 313.06 | 280.01 |
| F9 | PF | 3.39 | 2.76 | 142.75 | 150.10 | 302.99 | 265.37 |
| F10 | PF*HA | 3.70 | 2.83 | 148.78 | 154.70 | 298.70 | 258.18 |
| F11 | PF*AC | 5.18 | 3.63 | 84.78 | 107.82 | 323.57 | 277.38 |
| F12 | PF*AC*HA | 5.43 | 3.94 | 86.50 | 109.90 | 320.82 | 280.04 |
| F13 | PF*AA | 3.44 | 2.78 | 157.60 | 161.60 | 305.40 | 266.92 |
| F14 | PF*AA*HA | 3.69 | 2.80 | 169.97 | 165.60 | 294.12 | 258.32 |
| F15 | PF*AA*AC | 5.43 | 3.78 | 87.03 | 110.10 | 314.19 | 285.10 |
| F16 | PF*AA*AC*HA | 5.18 | 3.85 | 90.60 | 110.14 | 326.61 | 285.69 |
| Treatments | NDF (g kg−1) | ADF (g kg−1) | ADL (g kg−1) | ||||
|---|---|---|---|---|---|---|---|
| 2018/19 | 2019/20 | 2018/19 | 2019/20 | 2018/19 | 2019/20 | ||
| Central points | |||||||
| C1–C8 | 413.65 | 485.21 | 183.68 | 228.73 | 27.29 | 45.65 | |
| Star points | |||||||
| S1 | 422.11 | 445.55 | 200.70 | 220.91 | 34.98 | 41.91 | |
| S2 | 422.64 | 456.27 | 201.20 | 223.66 | 33.54 | 40.34 | |
| S3 | 404.08 | 451.24 | 182.21 | 210.00 | 26.05 | 44.17 | |
| S4 | 407.28 | 455.18 | 180.29 | 209.26 | 29.16 | 40.01 | |
| S5 | 388.62 | 410.12 | 162.86 | 157.00 | 24.17 | 22.43 | |
| S6 | 532.28 | 555.25 | 288.00 | 269.00 | 69.00 | 60.97 | |
| S7 | 417.88 | 497.23 | 187.73 | 230.97 | 29.95 | 46.32 | |
| S8 | 412.70 | 490.94 | 189.16 | 226.99 | 24.45 | 41.01 | |
| Factorial points | |||||||
| F1 | 1 | 400.56 | 435.89 | 180.95 | 183.03 | 27.83 | 32.01 |
| F2 | HA | 393.39 | 443.95 | 176.85 | 185.79 | 24.47 | 31.76 |
| F3 | AC | 450.71 | 516.09 | 213.79 | 251.16 | 53.50 | 54.59 |
| F4 | HA*AC | 451.13 | 534.71 | 217.12 | 260.76 | 51.31 | 55.38 |
| F5 | AA | 391.82 | 445.45 | 177.30 | 194.39 | 26.54 | 31.89 |
| F6 | AA*HA | 393.39 | 432.84 | 178.82 | 183.87 | 27.13 | 34.41 |
| F7 | AA*AC | 481.83 | 542.72 | 243.10 | 264.54 | 49.87 | 57.77 |
| F8 | AA*AC*HA | 497.13 | 520.39 | 245.52 | 256.93 | 54.45 | 56.66 |
| F9 | PF | 397.13 | 446.53 | 179.10 | 195.66 | 26.41 | 32.43 |
| F10 | PF*HA | 396.19 | 437.12 | 179.45 | 197.14 | 25.67 | 37.50 |
| F11 | PF*AC | 456.65 | 538.8 | 224.68 | 270.94 | 57.29 | 55.86 |
| F12 | PF*AC*HA | 440.68 | 532.06 | 200.74 | 255.10 | 52.11 | 54.34 |
| F13 | PF*AA | 382.00 | 458.18 | 175.83 | 207.92 | 25.41 | 38.15 |
| F14 | PF*AA*HA | 390.00 | 449.78 | 172.72 | 209.09 | 25.50 | 39.34 |
| F15 | PF*AA*AC | 493.78 | 523.20 | 262.47 | 270.45 | 55.11 | 56.49 |
| F16 | PF*AA*AC*HA | 496.79 | 532.87 | 263.45 | 271.58 | 52.03 | 57.17 |
| Treatments | DOM (g kg−1) | DNDF (g kg−1) | |||
|---|---|---|---|---|---|
| 2018/19 | 2019/20 | 2018/19 | 2019/20 | ||
| Central points | |||||
| C1–C8 | 553.87 | 455.12 | 286.25 | 265.00 | |
| Star points | |||||
| S1 | 545.00 | 467.00 | 273.00 | 260.00 | |
| S2 | 563.00 | 455.00 | 278.00 | 271.00 | |
| S3 | 558.00 | 470.00 | 297.00 | 254.00 | |
| S4 | 549.00 | 483.00 | 288.00 | 256.00 | |
| S5 | 675.00 | 650.00 | 390.00 | 365.00 | |
| S6 | 267.00 | 228.00 | 165.00 | 105.00 | |
| S7 | 574.00 | 464.00 | 286.00 | 265.00 | |
| S8 | 542.00 | 445.00 | 283.00 | 268.00 | |
| Factorial points | |||||
| F1 | 1 | 583.00 | 536.00 | 300.00 | 280.00 |
| F2 | HA | 562.00 | 525.00 | 311.00 | 284.00 |
| F3 | AC | 430.00 | 405.00 | 211.00 | 230.00 |
| F4 | HA*AC | 450.00 | 419.00 | 254.00 | 247.00 |
| F5 | AA | 584.00 | 508.00 | 310.00 | 288.00 |
| F6 | AA*HA | 591.00 | 531.00 | 304.00 | 311.00 |
| F7 | AA*AC | 465.00 | 395.00 | 253.00 | 239.00 |
| F8 | AA*AC*HA | 474.00 | 417.00 | 230.00 | 257.00 |
| F9 | PF | 587.00 | 528.00 | 301.00 | 294.00 |
| F10 | PF*HA | 594.00 | 519.00 | 312.00 | 320.00 |
| F11 | PF*AC | 430.00 | 392.00 | 236.00 | 238.00 |
| F12 | PF*AC*HA | 451.00 | 419.00 | 257.00 | 243.00 |
| F13 | PF*AA | 577.00 | 522.00 | 311.00 | 282.00 |
| F14 | PF*AA*HA | 582.00 | 531.00 | 316.00 | 315.00 |
| F15 | PF*AA*AC | 454.00 | 416.00 | 244.00 | 240.00 |
| F16 | PF*AA*AC*HA | 462.00 | 403.00 | 251.00 | 249.00 |
| Characteristic | Seasons | Equation | R2 | C.V% |
|---|---|---|---|---|
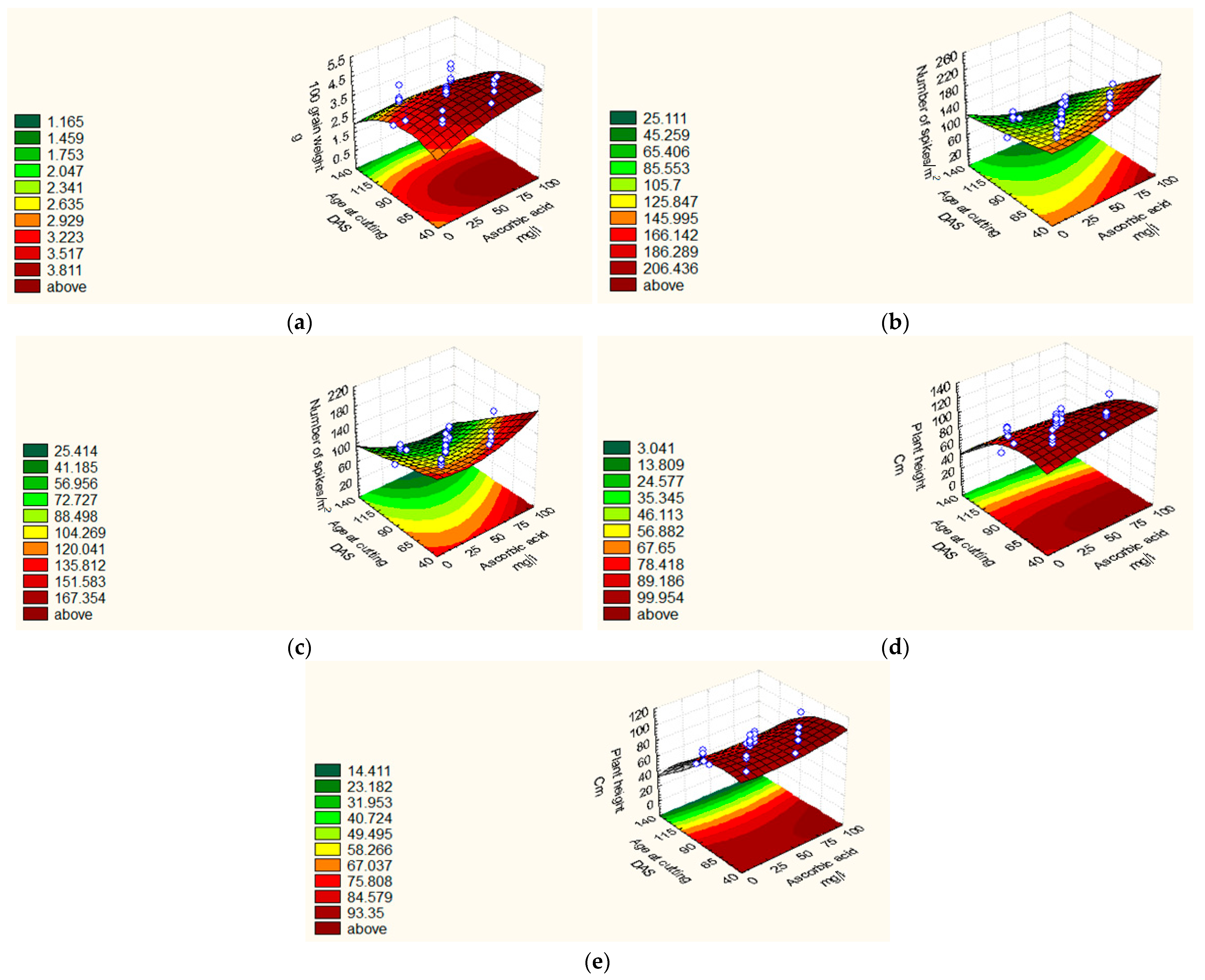
| 100 GW (g) | 18/19 | Y = 3.37 + 0.22 X21 + 0.14 X22 − 0.81 X3 − 0.18 X23 + 0.13 X24. | 0.83 | 6.42 |
| 19/20 | Y = 3.62 − 0.08 X1 + 0.09 X21 + 0.12 X2 − 0.08 X22 − 0.56 X3 − 0.27 X23 + 0.08 X4 − 0.11 X2X3. | 0.68 | 4.18 | |
| NSM−2 | 18/19 | Y = 84.75 + 6.27 X22 − 34.50 X3 + 8.27 X24 − 10.25 X2X3, | 0.89 | 11.25 |
| 19/20 | Y = 72.37 +3.50 X1+ 6.78 X22 − 28.25 X3 +3.75 X4 + 6.78 X24 − 6.75 X2X3, | 0.89 | 7.97 | |
| PH (cm) | 18/19 | Y = 86.87 − 22.86 X3 − 7.77 X23 + 2.59 X4 − 3.38 X2X3. | 0.92 | 1.41 |
| 19/20 | Y = 76.42 − 20.97 X3 − 7.05 X23 + 4.71 X4 + 1.57 X24 − 1.14 X2X3. | 0.94 | 2.27 | |
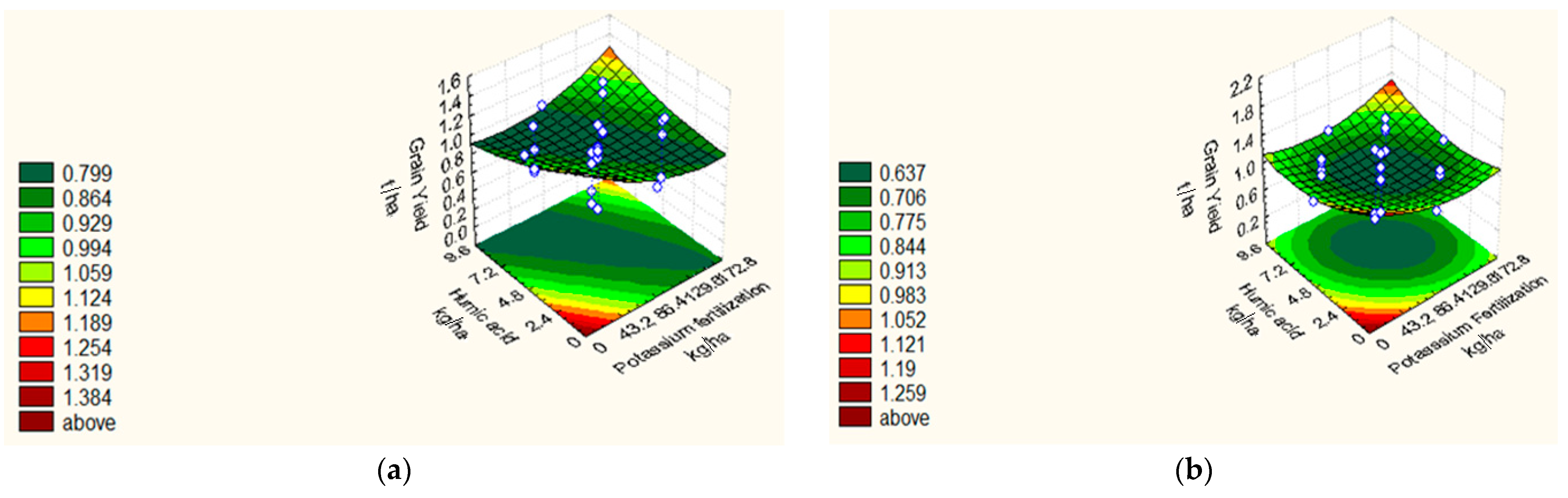
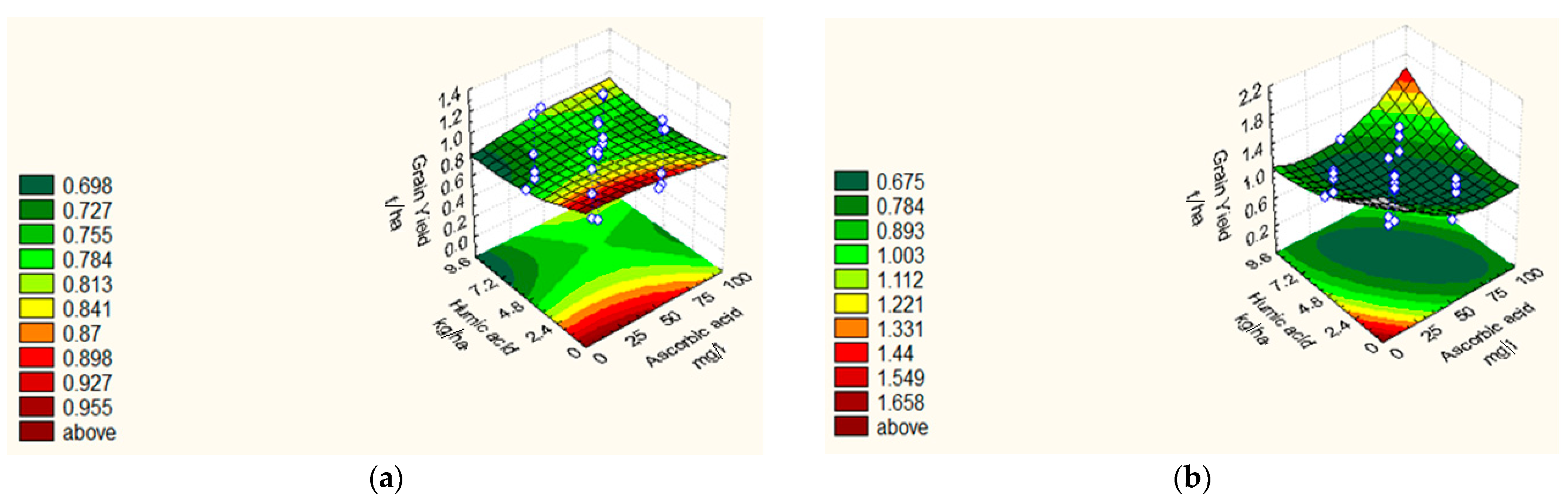
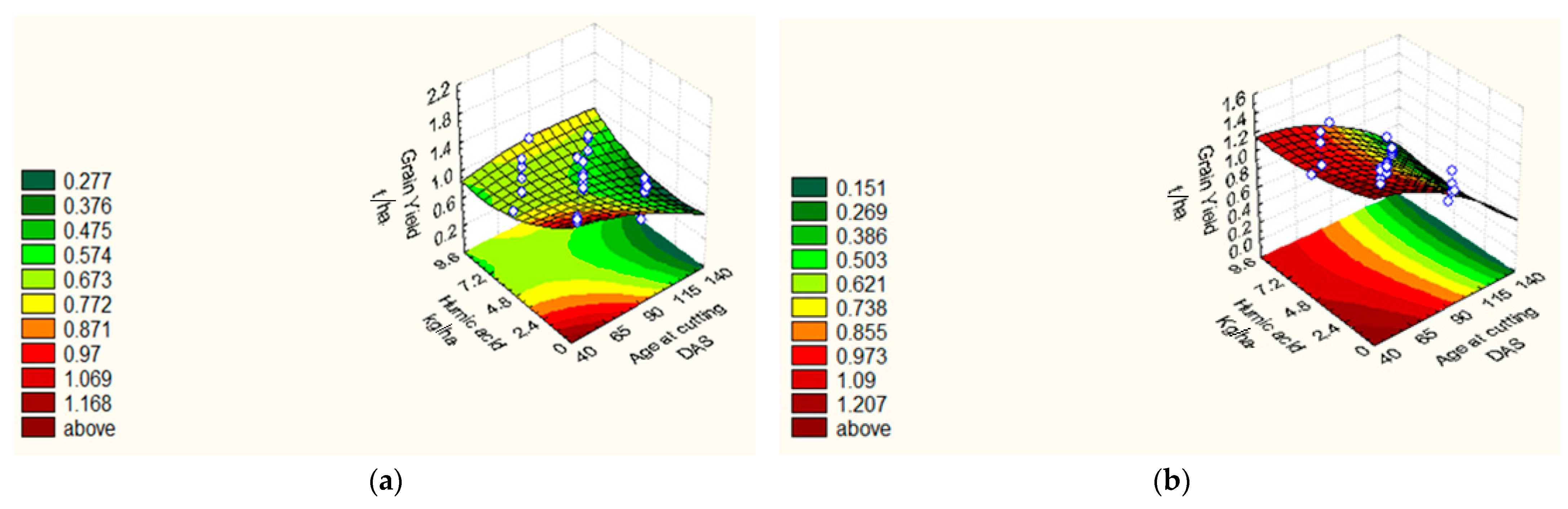
| GY (t ha−1) | 18/19 | Y = 0.76 − 0.02 X1 + 0.04 X21 − 0.01 X22 − 0.26 X3 − 0.05 X23 − 0.03 X4 + 0.02 X24 + 0.06 X1X4 + 0.02 X2X4 + 0.01 X3X4. | 0.93 | 2.59 |
| 19/20 | Y = 0.57 − 0.02 X1 + 0.05 X21 − 0.03 X2 + 0.09 X22 − 0.15 X3 − 0.03 X23 − 0.04 X4 + 0.06 X24 + 0.03X1X4 + 0.09 X2X4 + 0.02 X3X4 | 0.75 | 1.40 |
| Treatments | 100 GW (g) | NSM−2 | GY (t ha−1) | PH (cm) | |||||
|---|---|---|---|---|---|---|---|---|---|
| 2018/19 | 2019/20 | 2018/19 | 2019/20 | 2018/19 | 2019/20 | 2018/19 | 2019/20 | ||
| Central points | |||||||||
| C1–C8 | 3.37 | 3.62 | 84.75 | 72.375 | 0.76 | 0.57 | 83.49 | 76.42 | |
| Star points | |||||||||
| S1 | 4.44 | 3.94 | 72.00 | 80.00 | 1.12 | 0.81 | 76.66 | 69.30 | |
| S2 | 3.84 | 3.62 | 100.00 | 88.00 | 0.75 | 0.68 | 97.00 | 74.40 | |
| S3 | 3.65 | 2.98 | 96.00 | 92.00 | 0.74 | 0.82 | 71.00 | 76.50 | |
| S4 | 3.94 | 3.17 | 120.00 | 116.00 | 0.67 | 0.76 | 93.33 | 90.20 | |
| S5 | 5.00 | 4.59 | 172.00 | 155.00 | 1.09 | 0.78 | 109.33 | 98.70 | |
| S6 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| S7 | 3.63 | 3.54 | 104.00 | 96.00 | 0.83 | 0.75 | 81.33 | 80.50 | |
| S8 | 3.92 | 3.70 | 128.00 | 112.00 | 0.84 | 0.76 | 92.33 | 87.30 | |
| Factorial points | |||||||||
| F1 | 1 | 3.92 | 3.42 | 112.00 | 96.00 | 1.21 | 1.31 | 89.33 | 68.50 |
| F2 | HA | 4.06 | 4.00 | 124.00 | 103.00 | 1.72 | 1.73 | 113.00 | 110.00 |
| F3 | AC | 3.01 | 3.10 | 72.00 | 67.00 | 0.74 | 0.61 | 65.33 | 54.20 |
| F4 | HA*AC | 3.94 | 3.87 | 96.00 | 73.00 | 0.44 | 0.67 | 71.00 | 60.40 |
| F5 | AA | 4.50 | 4.23 | 132.00 | 112.00 | 1.44 | 1.52 | 96.33 | 80.60 |
| F6 | AA*HA | 4.77 | 4.49 | 164.00 | 124.00 | 1.02 | 0.86 | 100.30 | 81.90 |
| F7 | AA*AC | 3.03 | 2.81 | 48.00 | 36.00 | 0.57 | 0.57 | 40.30 | 40.00 |
| F8 | AA*AC*HA | 2.84 | 2.94 | 64.00 | 40.00 | 0.47 | 0.58 | 55.00 | 57.00 |
| F9 | PF | 4.20 | 4.23 | 148.00 | 110.00 | 1.50 | 1.33 | 96.00 | 88.00 |
| F10 | PF*HA | 4.68 | 3.45 | 104.00 | 112.00 | 1.16 | 0.78 | 101.00 | 91.30 |
| F11 | PF*AC | 2.98 | 2.86 | 80.00 | 71.00 | 0.46 | 0.75 | 56.00 | 43.40 |
| F12 | PF*AC*HA | 3.11 | 2.98 | 80.00 | 80.00 | 0.51 | 0.43 | 69.00 | 56.00 |
| F13 | PF*AA | 4.44 | 3.76 | 120.00 | 100.00 | 1.17 | 0.83 | 100.66 | 93.00 |
| F14 | PF*AA*HA | 4.44 | 3.79 | 152.00 | 112.00 | 1.02 | 1.01 | 102.66 | 102.60 |
| F15 | PF*AA*AC | 3.17 | 3.58 | 60.00 | 64.00 | 0.45 | 0.81 | 48.20 | 45.50 |
| F16 | PF*AA*AC*HA | 3.27 | 3.89 | 72.00 | 70.00 | 0.53 | 0.71 | 64.44 | 53.30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd-Elatty, S.A.A.; Nawar, A.I.; Salama, H.S.A.; Khattab, I.M.; Shaalan, A.M. The Production of Dual-Purpose Triticale in Arid Regions: Application of Organic and Inorganic Treatments under Water Deficit Conditions. Agronomy 2022, 12, 1251. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12061251
Abd-Elatty SAA, Nawar AI, Salama HSA, Khattab IM, Shaalan AM. The Production of Dual-Purpose Triticale in Arid Regions: Application of Organic and Inorganic Treatments under Water Deficit Conditions. Agronomy. 2022; 12(6):1251. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12061251
Chicago/Turabian StyleAbd-Elatty, Sara A. A., Ali I. Nawar, Heba S. A. Salama, Ibrahim M. Khattab, and Ahmed M. Shaalan. 2022. "The Production of Dual-Purpose Triticale in Arid Regions: Application of Organic and Inorganic Treatments under Water Deficit Conditions" Agronomy 12, no. 6: 1251. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12061251