
Comparative Genetic Diversity Analysis for Biomass Allocation and Drought Tolerance in Wheat

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Germplasm
2.2. Phenotyping Trials
2.3. Analysis of Phenotypic Data
2.4. Genotyping
2.5. Analysis of Genotypic Data
3. Results
3.1. Phenotyping
3.1.1. Genotype and Water Regime Effects on Agronomic Traits and Grain Yield
3.1.2. Principal Components of Phenotypic Data
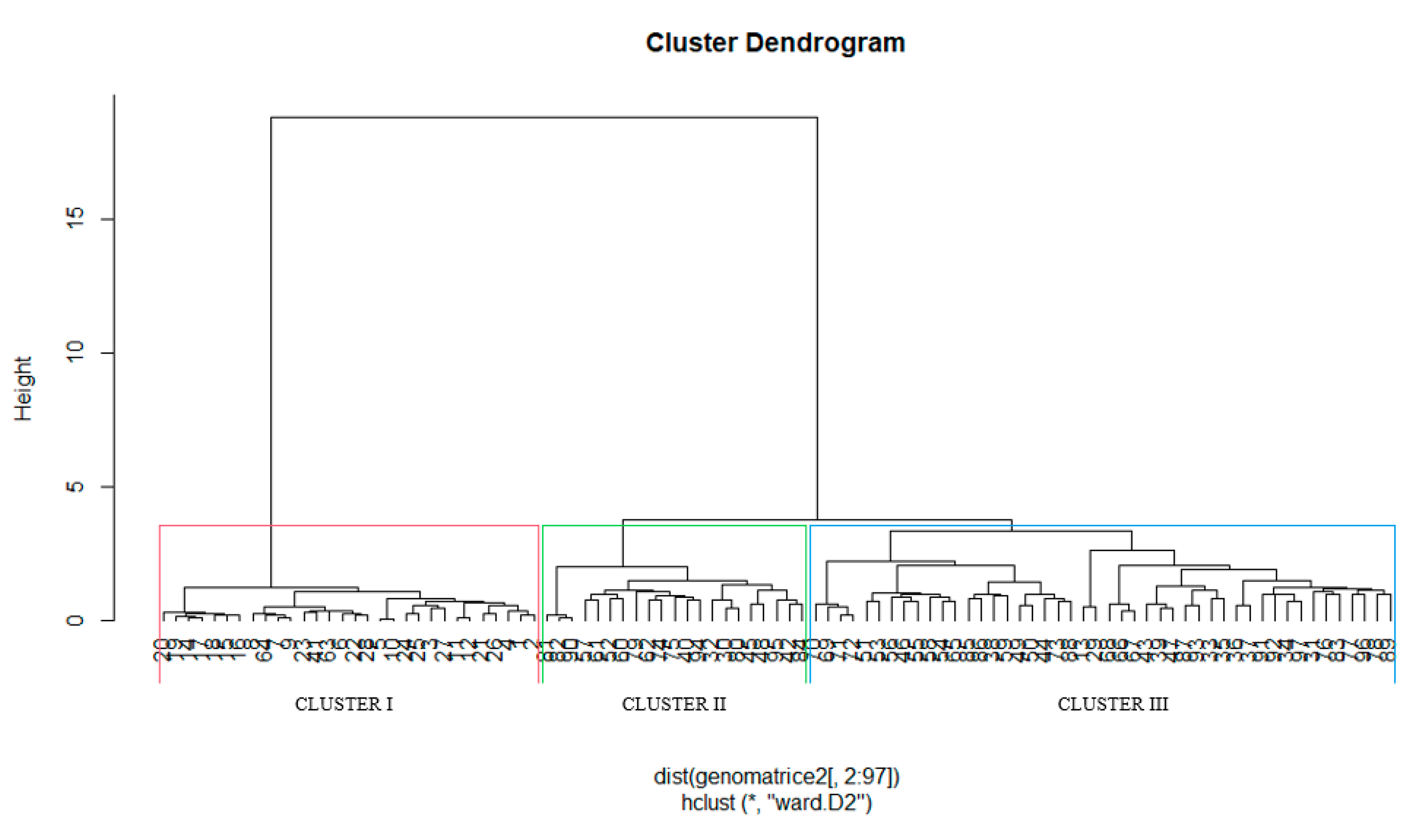
3.1.3. Phenotypic Hierarchical Clustering
3.2. Genotyping
3.2.1. Population Genetic Parameters Based on SNP Markers
3.2.2. Analysis of Molecular Variance and Genotypic Hierarchical Clustering
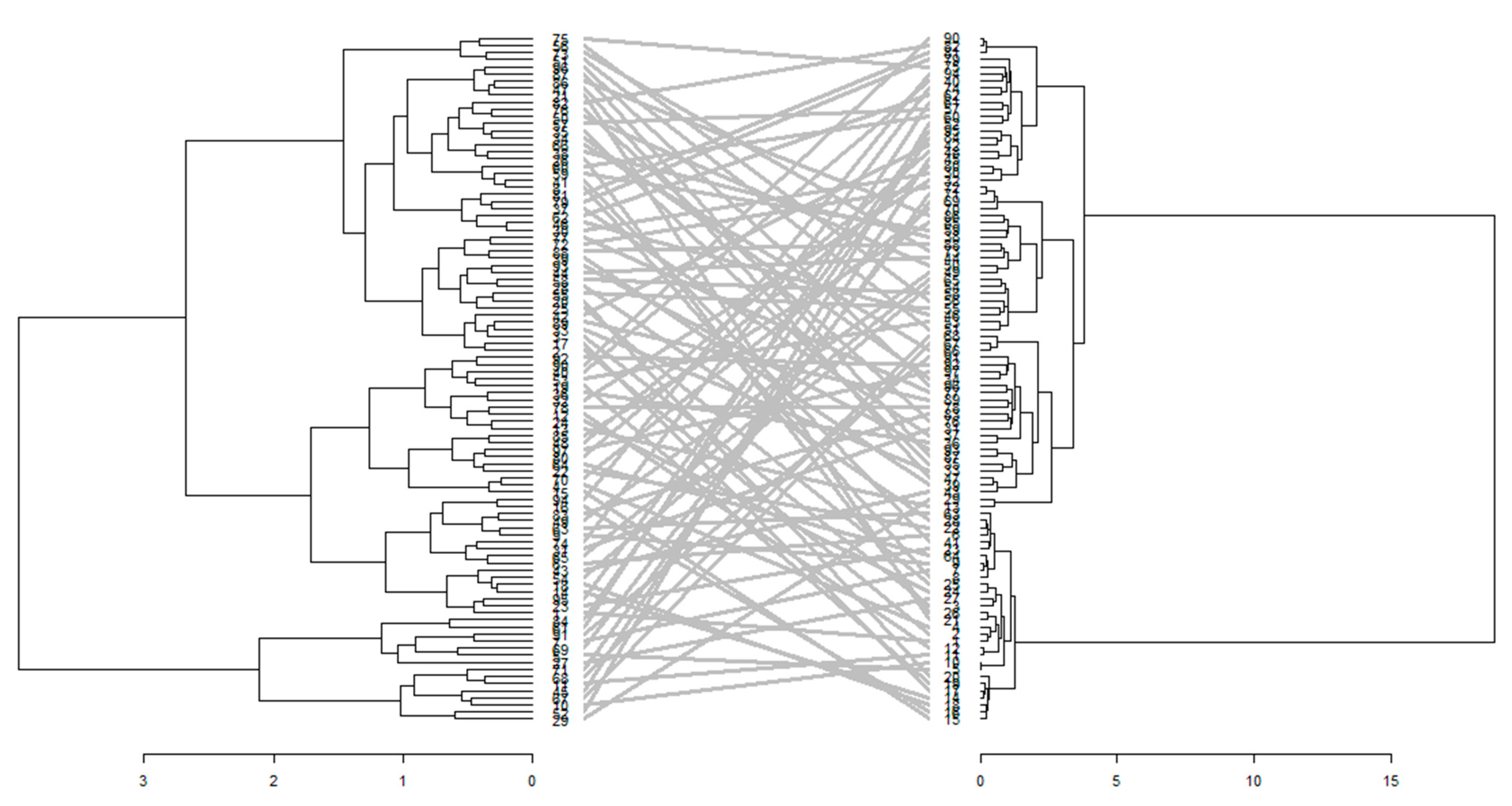
3.2.3. Comparison of Phenotypic and Genotypic Hierarchical Clusters
4. Discussion
4.1. Genotypic Variation for Agronomic Traits and Biomass Allocation
4.2. Multivariate Relationships Explained by Principal Components
4.3. Phenotypic Clustering of Genotypes and Implications for Drought Tolerance and Carbon Sequestration Breeding
4.4. Genotypic Clustering and Molecular Variance
4.5. Genotypic and Phenotypic Divergence under Different Water Regimes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT Wheat Production Statistics. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 4 March 2021).
- Tadesse, W.; Bishaw, Z.; Assefa, S. Wheat production and breeding in Sub-Saharan Africa: Challenges and opportunities in the face of climate change. Int. J. Clim. Chang. Strateg. Manag. 2019, 11, 696–715. [Google Scholar] [CrossRef] [Green Version]
- Yahaya, M.A.; Shimelis, H. Drought stress in sorghum: Mitigation strategies, breeding methods and technologies—A review. J. Agron. Crop. Sci. 2022, 208, 127–142. [Google Scholar] [CrossRef]
- Zougmoré, R.B.; Partey, S.T.; Ouédraogo, M.; Torquebiau, E.; Campbell, B.M. Facing climate variability in sub-Saharan Africa: Analysis of climate-smart agriculture opportunities to manage climate-related risks. Cah. Agric. 2018, 27, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Pironon, S.; Etherington, T.R.; Borrell, J.S.; Kühn, N.; Macias-Fauria, M.; Ondo, I.; Tovar, C.; Wilkin, P.; Willis, K.J. Potential adaptive strategies for 29 sub-Saharan crops under future climate change. Nat. Clim. 2019, 9, 758–763. [Google Scholar] [CrossRef]
- Voss-Fels, K.P.; Snowdon, R.J.; Hickey, L.T. Designer roots for future crops. Trends Plant. Sci. 2018, 23, 957–960. [Google Scholar] [CrossRef]
- Wasson, A.P.; Richards, R.; Chatrath, R.; Misra, S.; Prasad, S.S.; Rebetzke, G.; Kirkegaard, J.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OlaOlorun, B.M.; Shimelis, H.A.; Mathew, I. Variability and selection among mutant families of wheat for biomass allocation, yield and yield-related traits under drought-stressed and non-stressed conditions. J. Agron. Crop. Sci. 2020, 207, 404–421. [Google Scholar] [CrossRef]
- Voss-Fels, K.P.; Stahl, A.; Wittkop, B.; Lichthardt, C.; Nagler, S.; Rose, T.; Chen, T.W.; Zetzsche, H.; Seddig, S.; Baig, M.M. Breeding improves wheat productivity under contrasting agrochemical input levels. Nat. Plants 2019, 5, 706–714. [Google Scholar] [CrossRef]
- Gram, G.; Roobroeck, D.; Pypers, P.; Six, J.; Merckx, R.; Vanlauwe, B. Combining organic and mineral fertilizers as a climate-smart integrated soil fertility management practice in sub-Saharan Africa: A meta-analysis. PLoS ONE 2020, 15, e0239552. [Google Scholar] [CrossRef]
- Junaidi, J.; Kallenbach, C.M.; Byrne, P.F.; Fonte, S.J. Root traits and root biomass allocation impact how wheat genotypes respond to organic amendments and earthworms. PLoS ONE 2018, 13, e0200646. [Google Scholar] [CrossRef]
- Nguyen, V.L.; Stangoulis, J. Variation in root system architecture and morphology of two wheat genotypes is a predictor of their tolerance to phosphorus deficiency. Acta Physiol. Plant 2019, 41, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Svane, S.F.; Füchtbauer, W.S.; Andersen, J.R.; Jensen, J.; Thorup-Kristensen, K. Genomic prediction of yield and root development in wheat under changing water availability. Plant Methods 2020, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Maeoka, R.E.; Sadras, V.O.; Ciampitti, I.A.; Diaz, D.R.; Fritz, A.K.; Lollato, R.P. Changes in the phenotype of winter wheat varieties released between 1920 and 2016 in response to in-furrow fertilizer: Biomass allocation, yield, and grain protein concentration. Front. Plant. Sci. 2020, 10, 1786. [Google Scholar] [CrossRef] [Green Version]
- Rufo, R.; Salvi, S.; Royo, C.; Soriano, J.M. Exploring the genetic architecture of root-related traits in Mediterranean bread wheat landraces by genome-wide association analysis. Agronomy 2020, 10, 613. [Google Scholar] [CrossRef]
- Paez-Garcia, A.; Motes, C.M.; Scheible, W.R.; Chen, R.; Blancaflor, E.B.; Monteros, M.J. Root traits and phenotyping strategies for plant improvement. Plants 2015, 4, 334–355. [Google Scholar] [CrossRef] [PubMed]
- Shamuyarira, K.W.; Shimelis, H.; Mathew, I.; Zengeni, R.; Chaplot, V. A meta-analysis of combining ability effects in wheat for agronomic traits and drought adaptation: Implications for optimizing biomass allocation. Crop Sci. 2022, 62, 139–156. [Google Scholar] [CrossRef]
- Manschadi, A.M.; Christopher, J.; deVoil, P.; Hammer, G.L. The role of root architectural traits in adaptation of wheat to water-limited environments. Funct. Plant Biol. 2006, 33, 823–837. [Google Scholar] [CrossRef] [Green Version]
- Hammer, G.L.; Dong, Z.; McLean, G.; Doherty, A.; Messina, C.; Schussler, J.; Zinselmeier, C.; Paszkiewicz, S.; Cooper, M. Can changes in canopy and/or root system architecture explain historical maize yield trends in the US corn belt? Crop Sci. 2009, 49, 299–312. [Google Scholar] [CrossRef]
- Chen, Y.L.; Djalovic, I.; Rengel, Z. Phenotyping for Root Traits. In Phenomics in Crop Plants: Trends, Options and Limitations; Kumar, J., Pratap, A., Eds.; Springer: New Delhi, India, 2015; pp. 101–128. [Google Scholar] [CrossRef]
- Suneja, Y.; Gupta, A.K.; Bains, N.S. Stress adaptive plasticity: Aegilops tauschii and Triticum dicoccoides as potential donors of drought associated morpho-physiological traits in wheat. Front. Plant Sci. 2019, 10, 211. [Google Scholar] [CrossRef] [Green Version]
- Schultz, J.C.; Appel, H.M.; Ferrieri, A.; Arnold, T.M. Flexible resource allocation during plant defense responses. Front. Plant Sci. 2013, 4, 324. [Google Scholar] [CrossRef] [Green Version]
- Pieruschka, R.; Schurr, U. Plant phenotyping: Past, present, and future. Plant. Phenomics 2019, 2019, 7507131. [Google Scholar] [CrossRef] [PubMed]
- Joshi, D.C.; Singh, V.; Hunt, C.; Mace, E.; van Oosterom, E.; Sulman, R.; Jordan, D.; Hammer, G. Development of a phenotyping platform for high throughput screening of nodal root angle in sorghum. Plant Methods 2017, 13, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgante, M.; Salamini, F. From plant genomics to breeding practice. Curr. Opin. Biotechnol. 2003, 14, 214–219. [Google Scholar] [CrossRef]
- Boukar, O.; Belko, N.; Chamarthi, S.; Togola, A.; Batieno, J.; Owusu, E.; Haruna, M.; Diallo, S.; Umar, M.L.; Olufajo, O. Cowpea (Vigna unguiculata): Genetics, genomics and breeding. Plant Breed 2019, 138, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Bohra, A.; Saxena, K.; Varshney, R.K.; Saxena, R.K. Genomics-assisted breeding for pigeon pea improvement. Theor. Appl. Genet. 2020, 133, 1721–1737. [Google Scholar] [CrossRef]
- Chen, X.; Min, D.; Yasir, T.A.; Hu, Y.G. Genetic diversity, population structure and linkage disequilibrium in elite Chinese winter wheat investigated with SSR markers. PLoS ONE 2012, 7, e44510. [Google Scholar] [CrossRef] [Green Version]
- Rufo, R.; Alvaro, F.; Royo, C.; Soriano, J.M. From landraces to improved cultivars: Assessment of genetic diversity and population structure of Mediterranean wheat using SNP markers. PLoS ONE 2019, 14, e0219867. [Google Scholar] [CrossRef] [Green Version]
- Nazarzadeh, Z.; Onsori, H.; Akrami, S. Genetic diversity of bread wheat (Triticum aestivum L.) genotypes using RAPD and ISSR molecular markers. J. Genet. Res. 2020, 6, 69–76. [Google Scholar] [CrossRef]
- Mammadov, J.; Aggarwal, R.; Buyyarapu, R.; Kumpatla, S. SNP markers and their impact on plant breeding. Int. J. Plant Genom. 2012, 2012, 728398. [Google Scholar] [CrossRef]
- Chung, Y.S.; Choi, S.C.; Jun, T.H.; Kim, C. Genotyping-by-sequencing: A promising tool for plant genetics research and breeding. Hortic. Environ. Biotechnol. 2017, 58, 425–431. [Google Scholar] [CrossRef]
- Thomson, M.J. High-throughput SNP genotyping to accelerate crop improvement. Plant Breed. Biotechnol. 2014, 2, 195–212. [Google Scholar] [CrossRef]
- Agre, P.; Asibe, F.; Darkwa, K.; Edemodu, A.; Bauchet, G.; Asiedu, R.; Adebola, P.; Asfaw, A. Phenotypic and molecular assessment of genetic structure and diversity in a panel of winged yam (Dioscorea alata) clones and cultivars. Sci. Rep. 2019, 9, 18221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Department of Agriculture, Forestry and Fisheries (DAFF) 2010. Available online: https://www.dalrrd.gov.za/Portals/0/Brochures%20and%20Production%20guidelines/Wheat%20-%20Production%20Guideline.pdf (accessed on 20 January 2021).
- Payne, R.; Murray, D.; Harding, S. An Introduction to the GenStat Command Language; VSN International: Hempstead, UK, 2017. [Google Scholar]
- IBM SPSS Statistics. 25 Software, IBM Corp, Version 25.0; IBM SPSS Statistics for Windows: Armonk, NY, USA, 2017. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. 2013 Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-137. Available online: https://cran.r-project.org/web/packages/nlme/nlme.pdf (accessed on 12 November 2021).
- Gower, J.C. A general coefficient of similarity and some of its properties. Biometrics 1971, 27, 857–871. [Google Scholar] [CrossRef]
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K.; Studer, M. 2013 Package ‘Cluster’. Available online: https://cran.microsoft.com/snapshot/2014-10-10/web/packages/cluster/cluster.pdf (accessed on 12 November 2021).
- Huang, J.; Ge, X.; Sun, M. Modified CTAB protocol using a silica matrix for isolation of plant genomic DNA. Biotechniques 2000, 28, 432–434. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaccard, P. 1908 Nouvelles recherches sur la distribution florale. Bull. Soc. Vaud. Sci. Nat. 1908, 44, 223–270. [Google Scholar]
- Garnier, S.; Ross, N.; Rudis, B.; Sciaini, M.; Scherer, C. 2018 Viridis: Default Color Maps from ‘Matplotlib’. R Package Version 0.5. 1. CRAN: The Comprehensive R Archive Network. Available online: https://cran.r-project.org/web/packages/viridis/viridis.pdf (accessed on 12 November 2021).
- Nehe, A.; Akin, B.; Sanal, T.; Evlice, A.K.; Ünsal, R.; Dinçer, N.; Demir, L.; Geren, H.; Sevim, I.; Orhan, Ş. Genotype x environment interaction and genetic gain for grain yield and grain quality traits in Turkish spring wheat released between 1964 and 2010. PLoS ONE 2019, 14, e0219432. [Google Scholar] [CrossRef] [Green Version]
- Hendriks, P.W.; Kirkegaard, J.; Lilley, J.M.; Gregory, P.; Rebetzke, G. A tillering inhibition gene influences root–shoot carbon partitioning and pattern of water use to improve wheat productivity in rainfed environments. J. Exp. Bot. 2016, 67, 327–340. [Google Scholar] [CrossRef]
- Bai, C.; Liang, Y.; Hawkesford, M.J. Identification of QTLs associated with seedling root traits and their correlation with plant height in wheat. J. Exp. Bot. 2013, 64, 1745–1753. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Zhang, W.; Gao, Q.; Yang, A.; Hu, X.; Zhang, J. Enhancement of drought resistance and biomass by increasing the amount of glycine betaine in wheat seedlings. Euphytica 2011, 177, 151–167. [Google Scholar] [CrossRef]
- Schneider, H.M.; Lynch, J.P. Should root plasticity be a crop breeding target? Front. Plant Sci. 2020, 11, 546. [Google Scholar] [CrossRef]
- Waines, J.G.; Ehdaie, B. Domestication and crop physiology: Roots of green-revolution wheat. Ann. Bot. 2007, 100, 991–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passioura, J. Roots and Drought Resistance. In Developments in Agricultural and Managed Forest Ecology; Elsevier: Amsterdam, The Netherlands, 1983; pp. 265–280. [Google Scholar]
- Palta, J.A.; Chen, X.; Milroy, S.P.; Rebetzke, G.J.; Dreccer, M.F.; Watt, M. Large root systems: Are they useful in adapting wheat to dry environments? Funct. Plant Biol. 2011, 38, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Wang, Y.; Xu, P.; Zhang, Z. Overexpression of a WRKY transcription factor TaWRKY2 enhances drought stress tolerance in transgenic wheat. Front. Plant Sci. 2018, 9, 997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinclair, T.R. Challenges in breeding for yield increase for drought. Trends Plant Sci. 2011, 16, 289–293. [Google Scholar] [CrossRef]
- Hirte, J.; Leifeld, J.; Abiven, S.; Oberholzer, H.R.; Mayer, J. Below ground carbon inputs to soil via root biomass and rhizodeposition of field-grown maize and wheat at harvest are independent of net primary productivity. Agric. Ecosyst. Environ. 2018, 265, 556–566. [Google Scholar] [CrossRef]
- Du, Y.; Chen, L.; Wang, Y.; Yang, Z.; Saeed, I.; Daoura, B.G.; Hu, Y.G. The combination of dwarfing genes Rht4 and Rht8 reduced plant height, improved yield traits of rainfed bread wheat (Triticum aestivum L.). Field Crops Res. 2018, 215, 149–155. [Google Scholar] [CrossRef]
- Shamuyarira, K.W.; Shimelis, H.A.; Mathew, I.; Tsilo, T.J. Correlation and path coefficient analyses of yield and yield components in drought-tolerant bread wheat populations. S Afr. J. Plant Soil 2019, 36, 367–374. [Google Scholar] [CrossRef]
- Figueroa-Bustos, V.; Palta, J.A.; Chen, Y.; Siddique, K.H. Early season drought largely reduces grain yield in wheat cultivars with smaller root systems. Plants 2019, 8, 305. [Google Scholar] [CrossRef] [Green Version]
- Aslam, M.A.; Ahmed, M.; Stöckle, C.O.; Higgins, S.S.; Hayat, R. Can growing degree days and photoperiod predict spring wheat phenology? Front Environ. Sci. 2017, 5, 57. [Google Scholar] [CrossRef]
- Mathew, I.; Shimelis, H.; Mutema, M.; Clulow, A.; Zengeni, R.; Mbava, N.; Chaplot, V. Selection of wheat genotypes for biomass allocation to improve drought tolerance and carbon sequestration into soils. J. Agron. Crop. Sci. 2019, 205, 385–400. [Google Scholar] [CrossRef]
- Hu, T.; Sørensen, P.; Wahlström, E.M.; Chirinda, N.; Sharif, B.; Li, X.; Olesen, J.E. Root biomass in cereals, catch crops and weeds can be reliably estimated without considering aboveground biomass. Agric. Ecosyst. Environ. 2018, 251, 141–148. [Google Scholar] [CrossRef]
- van Ginkel, M.; Ogbonnaya, F. Novel genetic diversity from synthetic wheats in breeding cultivars for changing production conditions. Field Crops Res. 2007, 104, 86–94. [Google Scholar] [CrossRef]
- Fu, Y.B. Understanding crop genetic diversity under modern plant breeding. Appl. Genet. 2015, 128, 2131–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girma, E. Genetic erosion of wheat (Triticum spp.): Concept, research results and challenges. J. Nat. Sci. Res. 2017, 7, 72–81. [Google Scholar]
- Naderi, S.; Fakheri, B.A.; Maali-Amiri, R.; Mahdinezhad, N. Tolerance responses in wheat landrace Bolani are related to enhanced metabolic adjustments under drought stress. Plant Physiol. Biochem. 2020, 150, 244–253. [Google Scholar] [CrossRef]
- Bhatta, M.; Morgounov, A.; Belamkar, V.; Poland, J.; Baenziger, P.S. Unlocking the novel genetic diversity and population structure of synthetic hexaploid wheat. BMC Genom. 2018, 19, 591. [Google Scholar] [CrossRef] [Green Version]
- Havaux, M.; Ernez, M.; Lannoye, R. Correlation between heat tolerance and drought tolerance in cereals demonstrated by rapid chlorophyll fluorescence tests. J. Plant Physiol. 1988, 133, 555–560. [Google Scholar] [CrossRef]
- Rampino, P.; Mita, G.; Fasano, P.; Borrelli, G.M.; Aprile, A.; Dalessandro, G.; De Bellis, L.; Perrotta, C. Novel durum wheat genes up-regulated in response to a combination of heat and drought stress. Plant Physiol. Biochem. 2012, 56, 72–78. [Google Scholar] [CrossRef]
- Acuña-Galindo, M.A.; Mason, R.E.; Subramanian, N.K.; Hays, D.B. Meta-analysis of wheat QTL regions associated with adaptation to drought and heat stress. Crop. Sci. 2015, 55, 477–492. [Google Scholar] [CrossRef]
- Tricker, P.J.; ElHabti, A.; Schmidt, J.; Fleury, D. The physiological and genetic basis of combined drought and heat tolerance in wheat. J. Exp. Bot. 2018, 69, 3195–3210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ennos, R.A. Maintenance of Genetic Variation in Plant Populations. In Evolutionary Biology; Hecht, M.K., Wallace, B., Prance, G.T., Eds.; Springer: Boston, MA, USA, 1983; Volume 16, pp. 129–155. [Google Scholar]
- Dreisigacker, S.; Zhang, P.; Warburton, M.; Skovmand, B.; Hoisington, D.; Melchinger, A. Genetic diversity among and within CIMMYT wheat landrace accessions investigated with SSRs and implications for plant genetic resources management. Crop Sci. 2005, 45, 653–661. [Google Scholar] [CrossRef] [Green Version]
- Royo, C.; Maccaferri, M.; Álvaro, F.; Moragues, M.; Sanguineti, M.C.; Tuberosa, R.; Maalouf, F.; del Moral, L.F.G.; Demontis, A.; Rhouma, S. Understanding the relationships between genetic and phenotypic structures of a collection of elite durum wheat accessions. Field Crops Res. 2010, 119, 91–105. [Google Scholar] [CrossRef]
- Haghighattalab, A.; Pérez, L.G.; Mondal, S.; Singh, D.; Schinstock, D.; Rutkoski, J.; Ortiz-Monasterio, I.; Singh, R.P.; Goodin, D.; Poland, J. Application of unmanned aerial systems for high throughput phenotyping of large wheat breeding nurseries. Plant Methods 2016, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Saint Pierre, C.; Crossa, J.L.; Bonnett, D.; Yamaguchi-Shinozaki, K.; Reynolds, M.P. Phenotyping transgenic wheat for drought resistance. J. Exp. Bot. 2012, 63, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, K.; Shabbir, G.; Ahmed, M.; Shah, K.N. Phenotyping for drought resistance in bread wheat using physiological and biochemical traits. Sci. Total Environ. 2020, 729, 139082. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tolerance Level | Entry | ||||||
|---|---|---|---|---|---|---|---|
| Tolerant | BW100 | BW111 | BW116 | BW120 | BW147 | BW149 | BW151 |
| BW152 | BW48 | BW63 | LM100 | LM16 | LM17 | LM26 | |
| LM29 | LM37 | LM51 | LM71 | LM76 | LM90 | ||
| Intermediate tolerant | BW103 | BW124 | BW127 | BW129 | BW141 | BW148 | BW157 |
| BW159 | BW162 | BW49 | BW58 | BW71 | BW80 | LM01 | |
| LM12 | LM14 | LM18 | LM19 | LM21 | LM22 | LM25 | |
| LM27 | LM30 | LM31 | LM32 | LM36 | LM39 | LM40 | |
| LM41 | LM42 | LM44 | LM46 | LM47 | LM49 | LM56 | |
| LM58 | LM60 | LM70 | LM72 | LM79 | LM83 | LM85 | |
| LM91 | LM93 | LM97 | LM99 | ||||
| Susceptible | BW128 | BW140 | BW142 | BW145 | BW150 | BW28 | LM15 |
| LM20 | LM23 | LM24 | LM28 | LM33 | LM35 | LM38 | |
| LM43 | LM48 | LM50 | LM52 | LM54 | LM55 | LM57 | |
| LM59 | LM75 | LM77 | LM80 | LM81 | LM82 | LM84 | |
| LM86 | LM96 | LM98 | |||||
| SOV | df | DTH | DTM | PH | TN | PB | SB | RB | RS | GY |
|---|---|---|---|---|---|---|---|---|---|---|
| Block | 19 | 96.00 *** | 225.91 *** | 999.83 *** | 18.00 *** | 7,029,511.00 *** | 2,506,321.00 *** | 66,525.00 *** | 0.03 * | 876,453.00 *** |
| Rep | 1 | 261.72 *** | 552.71 *** | 7673.14 *** | 68.44 *** | 31,575,567.00 *** | 15,118,430.00 *** | 157,718.00 *** | 0.01 | 2,285,866.00 *** |
| Genotype (Gen) | 96 | 197.51 *** | 124.22 *** | 167.49 *** | 16.05 *** | 711,194.00 * | 270,175.00 *** | 15,507.00 *** | 0.03 * | 146,304.00 * |
| Water Regime (WR) | 1 | 53.6 | 28,022.20 *** | 31,765.90 *** | 3358.07 *** | 110,774,907.00 *** | 22,763,489.00 *** | 1,883,093.00 *** | 1.11 *** | 18,109,833.00 *** |
| Site | 2 | 74,612.12 *** | 125,380.64 *** | 134,122.84 *** | 2746.88 *** | 1,594,700,477.00 *** | 617,123,646.00 *** | 11,896,156.00 *** | 35.55 *** | 192,151,512.00 *** |
| Gen*WR | 96 | 22.31 * | 39.93 *** | 30.77 | 5.07 | 437,549 | 144,938 | 11,995 | 0.02 | 82,041 |
| Gen*Site | 192 | 85.02 *** | 61.80 *** | 65.12 *** | 9.69 *** | 657,583.00 * | 267,142.00 *** | 15,107.00 *** | 0.02 ** | 123,754 |
| Gen*WR*Site | 192 | 19.72 | 35.19 *** | 35.91 | 7.42 ** | 495,229 | 156,342 | 11,818 | 0.03 * | 92,026 |
| Residual | 561 | 17.21 | 23.47 | 30.18 | 5.43 | 532,134 | 160,226 | 10,445 | 0.02 | 106,404 |
| ENTRY | DTH | DTM | PH | TN | PB | SB | RB | RS | GY | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DS | NS | DS | NS | DS | NS | DS | NS | DS | NS | DS | NS | DS | NS | DS | NS | DS | NS | |
| Top ten genotypes | ||||||||||||||||||
| LM52 | 56.51 | 64.52 | 103.8 | 118.1 | 60.6 | 75.94 | 9.83 | 15.84 | 2137 | 3089 | 994 | 1487 | 214.2 | 352.4 | 0.43 | 0.53 | 929.4 | 1249.9 |
| LM30 | 66.12 | 66.73 | 109.3 | 116.3 | 67.12 | 82.39 | 11.11 | 12.13 | 2513 | 2600 | 1260 | 1370 | 325.9 | 295.4 | 0.5 | 0.49 | 927.7 | 934.4 |
| BW157 | 65.13 | 65.85 | 104.4 | 114.8 | 65.42 | 75.11 | 7.94 | 10.84 | 2091 | 2145 | 1060 | 1042 | 249.4 | 298.1 | 0.42 | 0.53 | 782 | 802.6 |
| BW152 | 64.39 | 65.53 | 108.6 | 116.8 | 65.09 | 75.73 | 10.36 | 11.5 | 1672 | 2417 | 945 | 1266 | 218.3 | 453.1 | 0.49 | 0.6 | 509.1 | 699.5 |
| BW140 | 68.16 | 57.36 | 108.5 | 115.6 | 69.72 | 66.36 | 7.59 | 15.85 | 1559 | 1978 | 850 | 1168 | 228.4 | 310 | 0.46 | 0.49 | 481.7 | 497.3 |
| LM47 | 69.05 | 67.27 | 112 | 121.1 | 65.35 | 78.57 | 9.1 | 12.25 | 1460 | 2901 | 898 | 1661 | 171.2 | 311.7 | 0.39 | 0.61 | 469 | 926.7 |
| LM70 | 68.25 | 68.83 | 109.2 | 119 | 68.11 | 74.79 | 8.61 | 13.84 | 1478 | 1990 | 839 | 990 | 190.9 | 274.9 | 0.48 | 0.57 | 449.1 | 722.1 |
| LM48 | 71.99 | 65.67 | 112.3 | 118.5 | 63.74 | 83.69 | 9.01 | 10 | 1606 | 2335 | 921 | 1181 | 238.4 | 276.8 | 0.4 | 0.55 | 447.4 | 876.7 |
| BW162 | 63.85 | 63.65 | 108.8 | 114.8 | 66.69 | 78.79 | 8.79 | 12.35 | 1358 | 2639 | 788 | 1288 | 160.7 | 280 | 0.45 | 0.62 | 414.7 | 1068.4 |
| LM54 | 61.5 | 69.71 | 105.8 | 119.7 | 60.62 | 78.49 | 11 | 11.67 | 1157 | 2952 | 653 | 1739 | 136.2 | 424.5 | 0.44 | 0.57 | 368 | 789.4 |
| Bottom five genotypes | ||||||||||||||||||
| LM39 | 69.16 | 68.61 | 109 | 118.5 | 66.39 | 79.89 | 7.68 | 13.03 | 1290 | 2195 | 799 | 1147 | 208.9 | 305.3 | 0.42 | 0.52 | 281.5 | 741.3 |
| LM44 | 70.22 | 66.17 | 102.6 | 113 | 66.61 | 83.28 | 9.2 | 12.84 | 1186 | 2174 | 716 | 1286 | 189.6 | 213.8 | 0.49 | 0.39 | 281.5 | 674.1 |
| BW147 | 73.45 | 67.16 | 111 | 117.4 | 65.07 | 76.27 | 7.34 | 9.7 | 1441 | 2167 | 940 | 1140 | 218.5 | 346 | 0.43 | 0.52 | 278.9 | 680.7 |
| LM55 | 65.43 | 63.15 | 108.1 | 113.7 | 60.29 | 76.94 | 9 | 14.45 | 1127 | 2034 | 778 | 1133 | 131.9 | 210.5 | 0.32 | 0.39 | 216.3 | 679.8 |
| LM29 | 64.83 | 66.39 | 112.8 | 122.9 | 62.22 | 80.86 | 8.5 | 13.08 | 1062 | 2662 | 710 | 1694 | 186.5 | 272.2 | 0.4 | 0.48 | 198 | 870.2 |
| Mean | 65.64 | 65.9 | 107.4 | 117.7 | 65.32 | 78.67 | 9.22 | 12.75 | 1490 | 2298 | 853.5 | 1252 | 206.2 | 295.1 | 0.43 | 0.49 | 444.3 | 757.1 |
| SEM | 0.85 | 0.41 | 0.47 | 0.3 | 0.49 | 0.44 | 0.1 | 0.16 | 23.86 | 37.75 | 13.26 | 24.68 | 4.87 | 6.05 | 0.01 | 0.01 | 12.19 | 15.48 |
| CV (%) | 12.72 | 6.16 | 4.33 | 2.49 | 7.35 | 5.52 | 10.81 | 12.63 | 15.77 | 16.18 | 15.31 | 19.42 | 23.26 | 20.19 | 13.99 | 13.04 | 26.88 | 20.14 |
| Range | 28.19 | 21.45 | 26.1 | 16.7 | 42.73 | 19.84 | 5.22 | 7.74 | 1451 | 2257 | 767 | 1498 | 400 | 268.5 | 0.34 | 0.3 | 731.4 | 825.6 |
| LSD (5%) | 4.3 | 5.29 | 6.61 | 4.17 | 6.68 | 4.87 | 0.35 | 3.16 | 532.7 | 1053 | 274.8 | 591.3 | 74.81 | 148.6 | 0.13 | 0.16 | 274.2 | 449.2 |
| R2 (%) | 80.86 | 72.25 | 77.06 | 82.45 | 76.48 | 74.24 | 44.81 | 33.04 | 79.7 | 76.79 | 78.78 | 73.41 | 14.99 | 20.71 | 19.43 | 42.68 | 62.26 | 68.91 |
| Traits | Drought-Stressed | Non-Stressed | ||||
|---|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC1 | PC2 | PC3 | |
| DTH | 0.35 | 0.83 | 0.05 | 0.14 | 0.88 | 0.10 |
| DTM | 0.40 | 0.74 | −0.08 | 0.19 | 0.74 | 0.01 |
| PH | 0.13 | 0.13 | −0.46 | 0.14 | 0.39 | 0.71 |
| TN | −0.12 | −0.56 | −0.15 | −0.05 | −0.53 | 0.10 |
| PB | 0.94 | −0.29 | −0.15 | 0.97 | −0.17 | 0.12 |
| SB | 0.87 | −0.03 | −0.27 | 0.86 | −0.07 | 0.22 |
| RB | 0.73 | 0.08 | 0.50 | 0.70 | 0.10 | −0.44 |
| RS | 0.18 | −0.19 | 0.87 | 0.22 | 0.29 | −0.74 |
| GY | 0.74 | −0.53 | −0.12 | 0.75 | −0.33 | 0.08 |
| Eigenvalue | 3.05 | 1.97 | 1.36 | 2.86 | 1.98 | 1.33 |
| Explained variance (%) | 33.92 | 21.87 | 15.08 | 31.80 | 21.99 | 14.81 |
| Cumulative variance (%) | 33.92 | 55.79 | 70.86 | 31.80 | 53.79 | 68.60 |
| Population | GD | PIC | MAF | Ho | F |
|---|---|---|---|---|---|
| Drought tolerant | 0.63 | 0.75 | 0.38 | 0.43 | −0.38 |
| Intermediate tolerance | 0.64 | 0.8 | 0.44 | 0.39 | −0.33 |
| Susceptible | 0.63 | 0.81 | 0.47 | 0.38 | −0.32 |
| Range | 0.20–0.88 | 0.24–1.00 | 0.05–0.50 | 0.29–0.58 | 0–−0.60 |
| Source | df | SS | MS | Estimated Variance | Proportion of Variance |
|---|---|---|---|---|---|
| Among Pops | 2 | 7713.672 | 3856.836 | 21.935 | 1% |
| Within Pops | 94 | 299,495.7 | 3186.124 | 3186.124 | 99% |
| Total | 96 | 307,209.3 | 3208.059 | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shamuyarira, K.W.; Shimelis, H.; Mathew, I.; Shayanowako, A.; Zengeni, R.; Chaplot, V. Comparative Genetic Diversity Analysis for Biomass Allocation and Drought Tolerance in Wheat. Agronomy 2022, 12, 1457. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12061457
Shamuyarira KW, Shimelis H, Mathew I, Shayanowako A, Zengeni R, Chaplot V. Comparative Genetic Diversity Analysis for Biomass Allocation and Drought Tolerance in Wheat. Agronomy. 2022; 12(6):1457. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12061457
Chicago/Turabian StyleShamuyarira, Kwame W., Hussein Shimelis, Isack Mathew, Admire Shayanowako, Rebecca Zengeni, and Vincent Chaplot. 2022. "Comparative Genetic Diversity Analysis for Biomass Allocation and Drought Tolerance in Wheat" Agronomy 12, no. 6: 1457. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12061457