
Drench Application of Soy Protein Hydrolysates Increases Tomato Plant Fitness, Fruit Yield, and Resistance to a Hemibiotrophic Pathogen

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Greenhouse Growth Conditions
2.2. Soy Protein Hydrolysate Treatments
2.3. Botrytis Cinerea
2.4. Pseudomonas syringae pv. Tomato DC3000-LUX (Pst-LUX)
2.5. Pathogen Inoculations
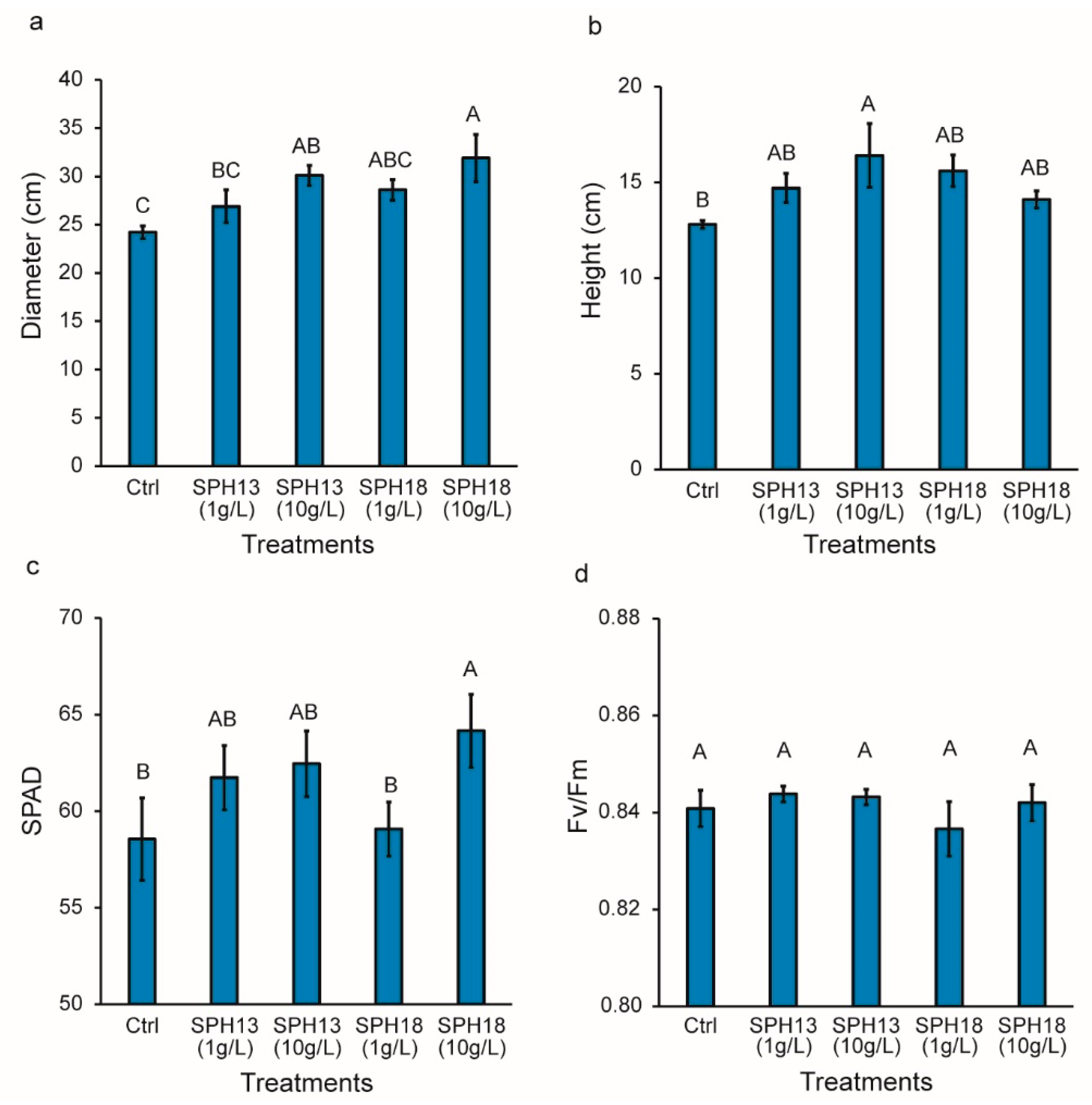
2.6. Plant Measurements
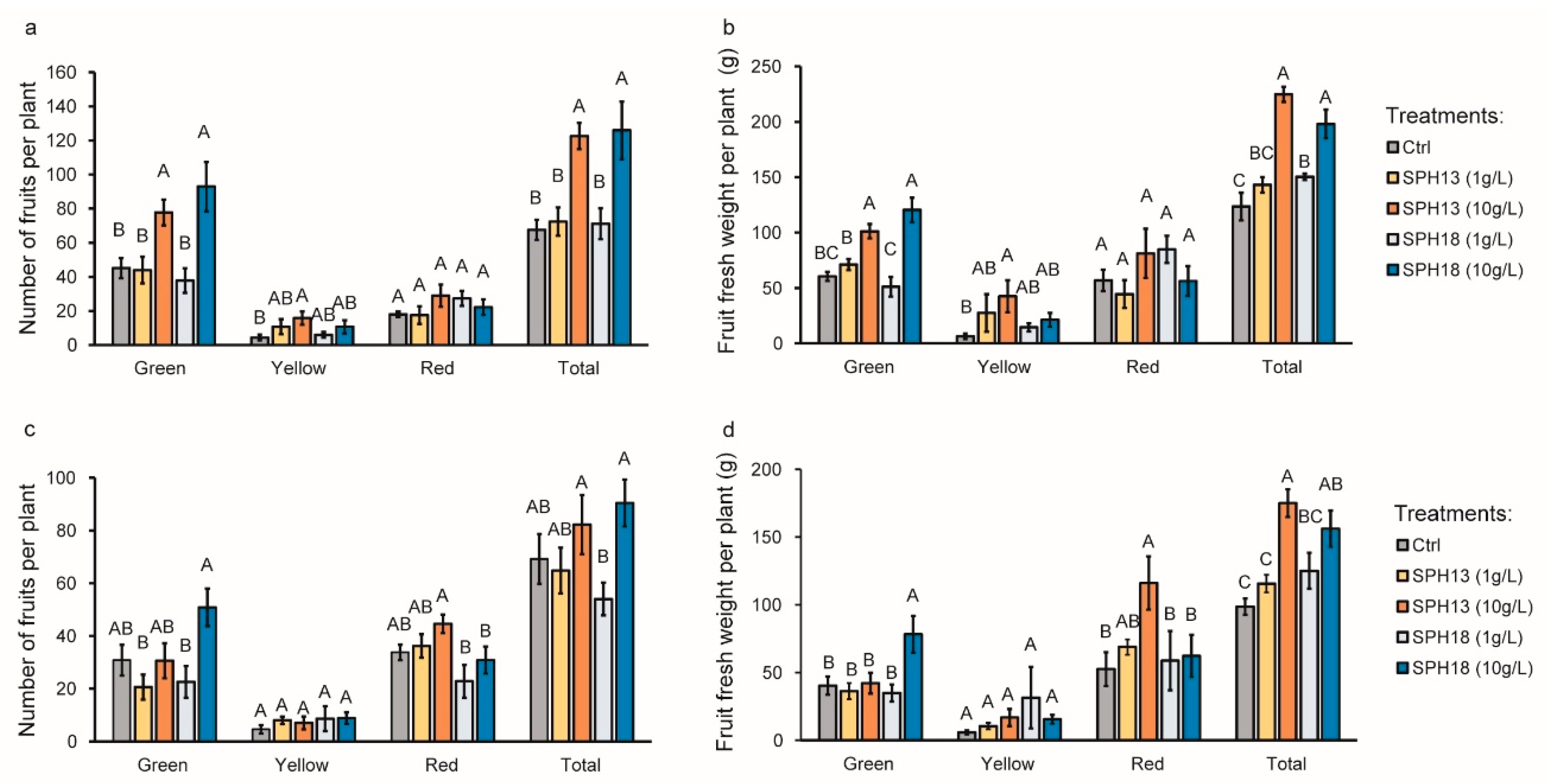
2.7. Fruit Yield Assessment
2.8. RNA Isolation and RT-qPCR
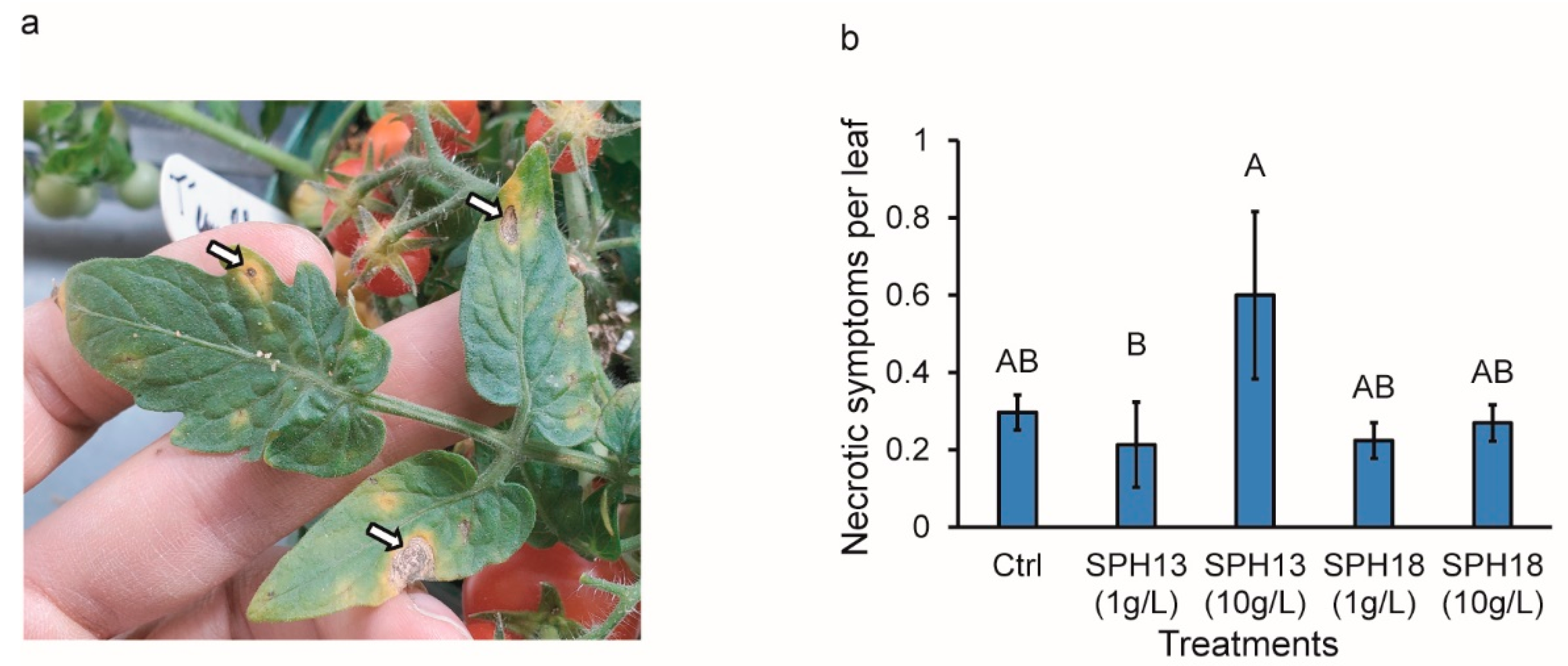
2.9. Disease Severity Assessments
2.10. Luciferase Assay and Bioluminescence Imaging
2.11. Nutrient Quantification
2.12. Statistical Analysis
3. Results
3.1. Soy Protein Hydrolysates Stimulate Growth and Fruit Productivity
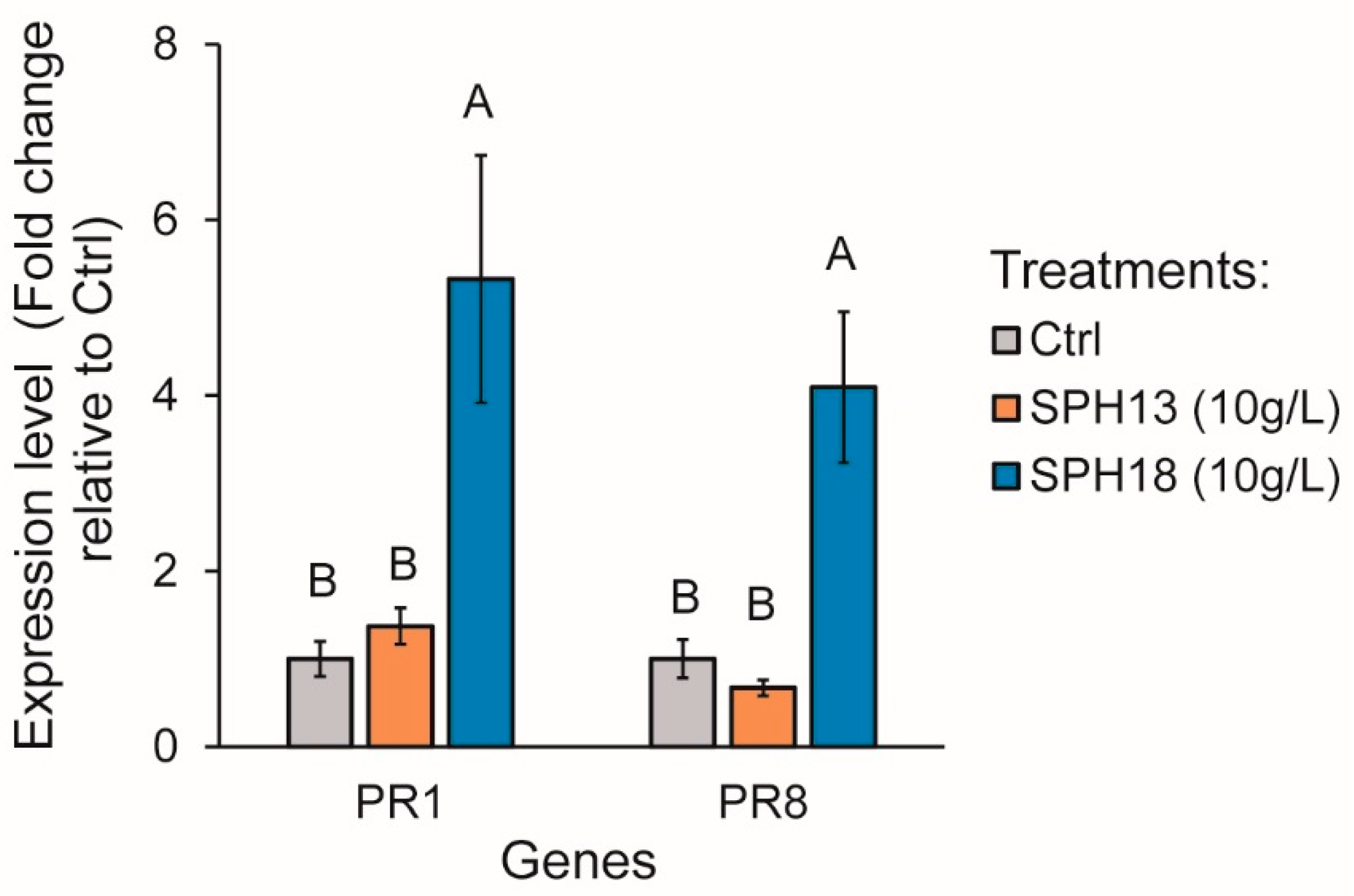
3.2. Soy Protein Hydrolysates Stimulate Defence Gene Expression
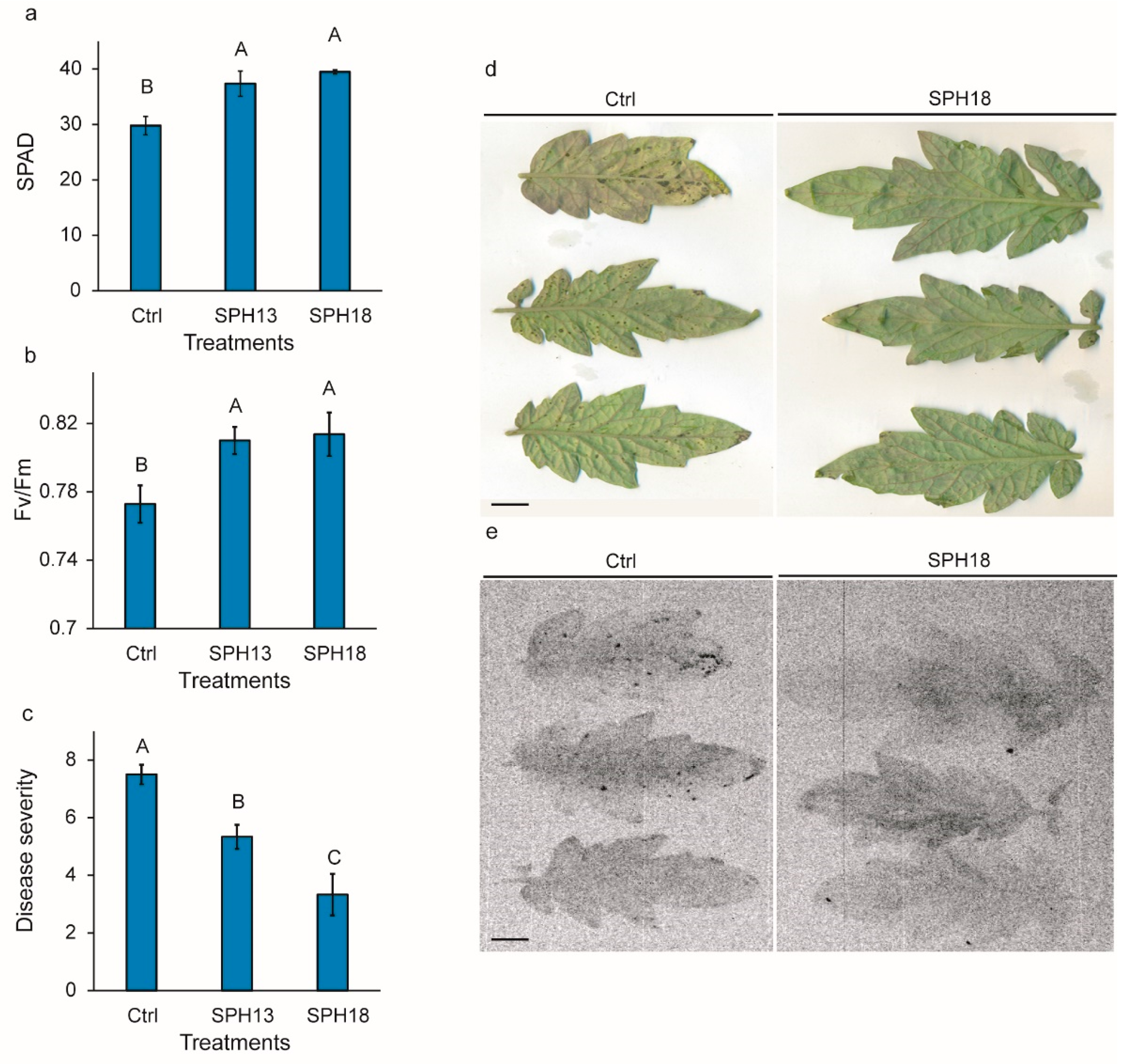
3.3. Soy Protein Hydrolysates Modulate Pathogen Resistance
4. Discussion
4.1. SPHs Increase Tomato Fruit Production
4.2. SPHs as a Nutrient Source
4.3. SPHs as Plant Biostimulants
4.4. SPHs as Stimulators of Plant Defences
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations World Population Prospects 2019-Highlights. In World Population Prospects 2019; Department of Economic and Social Affairs: New York, NY, USA, 2019; pp. 1–39.
- van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar] [CrossRef]
- Pimentel, D. Green revolution agriculture and chemical hazards. Sci. Total Environ. 1996, 188, S86–S98. [Google Scholar] [CrossRef]
- Jenkinson, D.S. The impact of humans on the nitrogen cycle, with focus on temperate arable agriculture. Plant Soil 2001, 228, 3–15. [Google Scholar] [CrossRef]
- Clark, S.; Klonsky, K.; Livingston, P.; Temple, S. Crop-yield and economic comparisons of organic, low-input, and conventional farming systems in California’s Sacramento Valley. Am. J. Altern. Agric. 1999, 14, 109–121. [Google Scholar] [CrossRef]
- Savci, S. Investigation of Effect of Chemical Fertilizers on Environment. APCBEE Procedia 2012, 1, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Davidson, E.A.; Mauzerall, D.L.; Searchinger, T.D.; Dumas, P.; Shen, Y. Managing nitrogen for sustainable development. Nature 2015, 528, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Dion, P.-P.; Jeanne, T.; Thériault, M.; Hogue, R.; Pepin, S.; Dorais, M. Nitrogen release from five organic fertilizers commonly used in greenhouse organic horticulture with contrasting effects on bacterial communities. Can. J. Soil Sci. 2020, 100, 120–135. [Google Scholar] [CrossRef]
- Dion, P.; Thériault, M.; Hunt, D.; Bittman, S.; Pepin, S.; Dorais, M. NLOS-OG: A management tool for nitrogen fertilization in organic greenhouses. Acta Hortic. 2020, 1296, 1093–1098. [Google Scholar] [CrossRef]
- Dorais, M. Advances in organic greenhouse cultivation. In Achieving Sustainable Greenhouse Cultivation; Marcelis, E.L., Heuvelink, E., Eds.; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; Chapter 5; pp. 121–176. [Google Scholar]
- De Pascale, S.; Rouphael, Y.; Colla, G. Plant biostimulants: Innovative tool for enhancing plant nutrition in organic farming. Eur. J. Hortic. Sci. 2018, 82, 277–285. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Hernandez, M.; Macias-Bobadilla, I.; Guevara-Gonzalez, R.G.; Romero-Gomez, S.D.J.; Rico-Garcia, E.; Ocampo-Velazquez, R.V.; Alvarez-Arquieta, L.D.L.; Torres-Pacheco, I. Plant Hormesis Management with Biostimulants of Biotic Origin in Agriculture. Front. Plant Sci. 2017, 8, 1762. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.A.; García-Cristobal, J.; Bonilla, A.; Ramos, B.; Gutierrez-Mañero, J. Beneficial rhizobacteria from rice rhizosphere confers high protection against biotic and abiotic stress inducing systemic resistance in rice seedlings. Plant Physiol. Biochem. 2014, 82, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Demir, S.; Danesh, Y.R.; Boyno, G.; Najafi, S. Arbuscular mycorrhizal fungi in biotic and abiotic stress conditions: Function and management in horticulture. In Sustainable Horticulture, 1st ed.; Seymen, M., Kurtar, E., Erdinc, C., Kumar, A., Eds.; Academic Press: Cambridge, MA, USA, 2022; Chapter 7; pp. 157–183. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sestili, F.; Rouphael, Y.; Cardarelli, M.; Pucci, A.; Bonini, P.; Canaguier, R.; Colla, G. Protein Hydrolysate Stimulates Growth in Tomato Coupled With N-Dependent Gene Expression Involved in N Assimilation. Front. Plant Sci. 2018, 9, 1233. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant Action of Protein Hydrolysates: Unraveling Their Effects on Plant Physiology and Microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, A.; Miras-Moreno, B.; Buffagni, V.; Senizza, B.; Pii, Y.; Cardarelli, M.; Rouphael, Y.; Colla, G.; Lucini, L. Foliar Application of Different Vegetal-Derived Protein Hydrolysates Distinctively Modulates Tomato Root Development and Metabolism. Plants 2021, 10, 326. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Matsumiya, Y.; Kubo, Y.M.A.M. Soybean Peptide: Novel Plant Growth Promoting Peptide from Soybean. In Soybean and Nutrition; El-Shemy, H., Ed.; InTechOpen: London, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Lachhab, N.; Sanzani, S.M.; Adrian, M.; Chiltz, A.; Balacey, S.; Boselli, M.; Ippolito, A.; Poinssot, B. Soybean and Casein Hydrolysates Induce Grapevine Immune Responses and Resistance against Plasmopara Viticola. Front. Plant Sci. 2014, 5, 716. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Fernald, R.D. Comprehensive Algorithm for Quantitative Real-Time Polymerase Chain Reaction. J. Comput. Biol. 2005, 12, 1047–1064. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [Green Version]
- González-Aguilera, K.L.; Saad, C.F.; Montes, R.A.C.; Alves-Ferreira, M.; De Folter, S. Selection of Reference Genes for Quantitative Real-Time RT-PCR Studies in Tomato Fruit of the Genotype MT-Rg1. Front. Plant Sci. 2016, 7, 1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safaie-Farahani, A.; Taghavi, S. Transcript analysis of some defense genes of tomato in response to host and non-host bacterial pathogens. Mol. Biol. Res. Commun. 2017, 6, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Diaz, A.H.; Kovács, I.; Lindermayr, C. Inducible Expression of the De-Novo Designed Antimicrobial Peptide SP1-1 in Tomato Confers Resistance to Xanthomonas campestris pv. vesicatoria. PLoS ONE 2016, 11, e0164097. [Google Scholar] [CrossRef]
- Stavridou, E.; Giannakis, I.; Karamichali, I.; Kamou, N.N.; Lagiotis, G.; Madesis, P.; Emmanouil, C.; Kungolos, A.; Nianiou-obeidat, I.; Lagopodi, A.L. Biosolid-amended Soil Enhances Defense Responses in Tomato Based on Metagenomic Profile and Expression of Pathogenesis-related Genes. Plants 2021, 10, 2789. [Google Scholar] [CrossRef]
- Isaac, R.A.; Johnson, W.C. Determination of Total Nitrogen in Plant Tissue, Using a Block Digestor. J. AOAC Int. 1976, 59, 98–100. [Google Scholar] [CrossRef]
- Analytical Methods for Atomic Absorption Spectroscopy. 1996. Available online: http://www1.lasalle.edu/~prushan/Intrumental%20Analysis_files/AA-Perkin%20Elmer%20guide%20to%20all!.pdf (accessed on 15 June 2022).
- Nkonge, C.; Ballance, G.M. A sensitive colorimetric procedure for nitrogen determination in micro-Kjeldahl digests. J. Agric. Food Chem. 1982, 30, 416–420. [Google Scholar] [CrossRef]
- Tandon, H.L.S.; Cescas, M.P.; Tyner, E.H. An Acid-Free Vanadate-Molybdate Reagent for the Determination of Total Phosphorus in Soils. Soil Sci. Soc. Am. J. 1968, 32, 48–51. [Google Scholar] [CrossRef]
- Reidinger, S.; Ramsey, M.H.; Hartley, S.E. Rapid and accurate analyses of silicon and phosphorus in plants using a portable X-ray fluorescence spectrometer. New Phytol. 2012, 195, 699–706. [Google Scholar] [CrossRef]
- Kiba, T.; Krapp, A. Plant Nitrogen Acquisition Under Low Availability: Regulation of Uptake and Root Architecture. Plant Cell Physiol. 2016, 57, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Serrani, J.C.; Fos, M.; Atarés, A.; García-Martínez, J.L. Effect of Gibberellin and Auxin on Parthenocarpic Fruit Growth Induction in the cv Micro-Tom of Tomato. J. Plant Growth Regul. 2007, 26, 211–221. [Google Scholar] [CrossRef]
- Matsuo, S.; Kikuchi, K.; Fukuda, M.; Honda, I.; Imanishi, S. Roles and regulation of cytokinins in tomato fruit development. J. Exp. Bot. 2012, 63, 5569–5579. [Google Scholar] [CrossRef] [PubMed]
- Bartels, S.; Boller, T. Quo vadis, Pep? Plant elicitor peptides at the crossroads of immunity, stress, and development. J. Exp. Bot. 2015, 66, 5183–5193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlot, A.C.; Sales, J.H.; Lenk, M.; Bauer, K.; Brambilla, A.; Sommer, A.; Chen, Y.; Wenig, M.; Nayem, S. Systemic propagation of immunity in plants. New Phytol. 2020, 229, 1234–1250. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Name | Primer Sequence | References |
|---|---|---|---|
| ACTIN2 | Act2_Fwd | CATTGTGCTCAGTGGTGGTTC | [26] |
| Act2_Rev | TCTGCTGGAAGGTGCTAAGTG | ||
| TUBULIN | Tub_Fwd | AACCTCCATTCAGGAGATGTTT | [27] |
| Tub_Rev | TCTGCTGTAGCATCCTGGTATT | ||
| UBIQUITIN | UBQ_Fwd | TCGTAAGGAGTGCCCTAATGCTGA | [28] |
| UBQ_Rev | CAATCGCCTCCAGCCTTGTTGTAA | ||
| SAND | SAND_Fwd | TTGCTTGGAGGAACAGACG | [26] |
| SAND_Rev | GCAAACAGAACCCCTGAATC | ||
| PR1 | PR1_Fwd | TCTTGTGAGGCCCAAAATTC | [29] |
| PR1_Rev | ATAGTCTGGCCTCTCGGACA | ||
| PR8/ CHITINASE 3 | PR8_Fwd | TGCAGGAACATTCACTGGAG | [29] |
| PR8_Rev | TAACGTTGTGGCATGATGGT |
| Treatment | |||
|---|---|---|---|
| Element | SPH13 | SPH18 | Unit |
| N | 13.551 (±0.053) | 18.660 (±0.018) | % |
| P | 0.213 (±0.004) | <0.01 | % |
| K | 0.364 (±0.023) | <0.01 | % |
| Ca | 0.025 (±0.004) | <0.01 | % |
| Mg | 0.040 (±0.004) | <0.01 | % |
| S | 1.204 (±0.003) | <0.01 | % |
| Cl | 1.421 (±0.001) | 0.577 (±0.061) | % |
| Si | 0.067 (±0.002) | 0.032 (±0.000) | % |
| Al | <0.17 | <0.17 | % |
| Mo | 5.035 (± 0.055) | 6.195 (±0.565) | mg/Kg |
| Sr | 4.405 (± 0.165) | 2.350 (±0.020) | mg/Kg |
| Rb | 6.920 (± 0.170) | <1 | mg/Kg |
| Pb | <1 | <1 | mg/Kg |
| Se | <2 | <2 | mg/Kg |
| As | <2 | <2 | mg/Kg |
| Hg | <3 | <3 | mg/Kg |
| Zn | <3 | <3 | mg/Kg |
| Cu | <9 | <9 | mg/Kg |
| Fe | 30.920 (±2.250) | <26 | mg/Kg |
| Mn | <17 | <17 | mg/Kg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrada, A.; Delisle-Houde, M.; Nguyen, T.T.A.; Tweddell, R.J.; Dorais, M. Drench Application of Soy Protein Hydrolysates Increases Tomato Plant Fitness, Fruit Yield, and Resistance to a Hemibiotrophic Pathogen. Agronomy 2022, 12, 1761. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12081761
Barrada A, Delisle-Houde M, Nguyen TTA, Tweddell RJ, Dorais M. Drench Application of Soy Protein Hydrolysates Increases Tomato Plant Fitness, Fruit Yield, and Resistance to a Hemibiotrophic Pathogen. Agronomy. 2022; 12(8):1761. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12081761
Chicago/Turabian StyleBarrada, Adam, Maxime Delisle-Houde, Thi Thuy An Nguyen, Russell J. Tweddell, and Martine Dorais. 2022. "Drench Application of Soy Protein Hydrolysates Increases Tomato Plant Fitness, Fruit Yield, and Resistance to a Hemibiotrophic Pathogen" Agronomy 12, no. 8: 1761. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy12081761