
Cultivar Susceptibility to Olive Knot Disease and Association with Endophytic Microbiota Community
 ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Orchard, Plant Material, and Field Monitoring
2.2. Pseudomonas savastanoi pv. savastanoi Isolation and Molecular Confirmation
2.3. Sample Collection and Processing for Microbiome Analysis
2.4. 16S and ITS Metagenomic Sequencing Library Preparation, Sequencing, and Analysis
2.5. Statistical Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Disease Intensity * | Cluster ** |
|---|---|---|
| Giarraffa | 3.92 A | Highly susceptible |
| Nocellara del Belice, Gioconda, Randazzese | 3.58 AB | Highly susceptible |
| Riondello | 3.42 ABC | Highly susceptible |
| Nerba Catanese | 3.33 ABCD | Highly susceptible |
| Buscionetto | 3.29 ABCD | Highly susceptible |
| Nasitana | 3.21 ABCD | Susceptible |
| Cerasuola | 3.17 ABCD | Highly susceptible |
| Luminario, Tortella, San Benedetto | 3.00 ABCD | Highly susceptible |
| Biancolilla nana (992) | 2.79 BCD | Susceptible |
| Bottone di Gallo, Aitana | 2.46 CDE | Susceptible |
| Castriciana2 | 2.38 DEF | Susceptible |
| Marmorina, Tonda Iblea | 1.75 EFG | Low resistant |
| Leucocarpa | 1.54 FGH | Low resistant |
| Minuta | 1.50 EFGHI | Low resistant |
| Virdisa | 1.46 FGHI | Low resistant |
| Santagatese | 1.38 GHI | Low resistant |
| Vaddarica | 1.33 GHI | Low resistant |
| Verdella | 1.21 GHI | Resistant |
| Biancolilla nana (1064), Biancolilla | 1.08 GHI | Low resistant |
| Nocellara messinese | 1.04 GHI | Resistant |
| Castriciana1, Calatina | 1.00 GHI | Resistant |
| Passalunara | 0.96 GHIJ | Resistant |
| Rizza | 0.79 GHIJ | Resistant |
| Calamignara | 0.71 HIJ | Resistant |
| Montonica, Zaituna | 0.54 IJ | Highly resistant |
| Verdello grosso, Turdunazza | 0.00 J | Highly resistant |
3. Results
3.1. Evaluation of Olive Knot Disease Susceptibility in Open Field
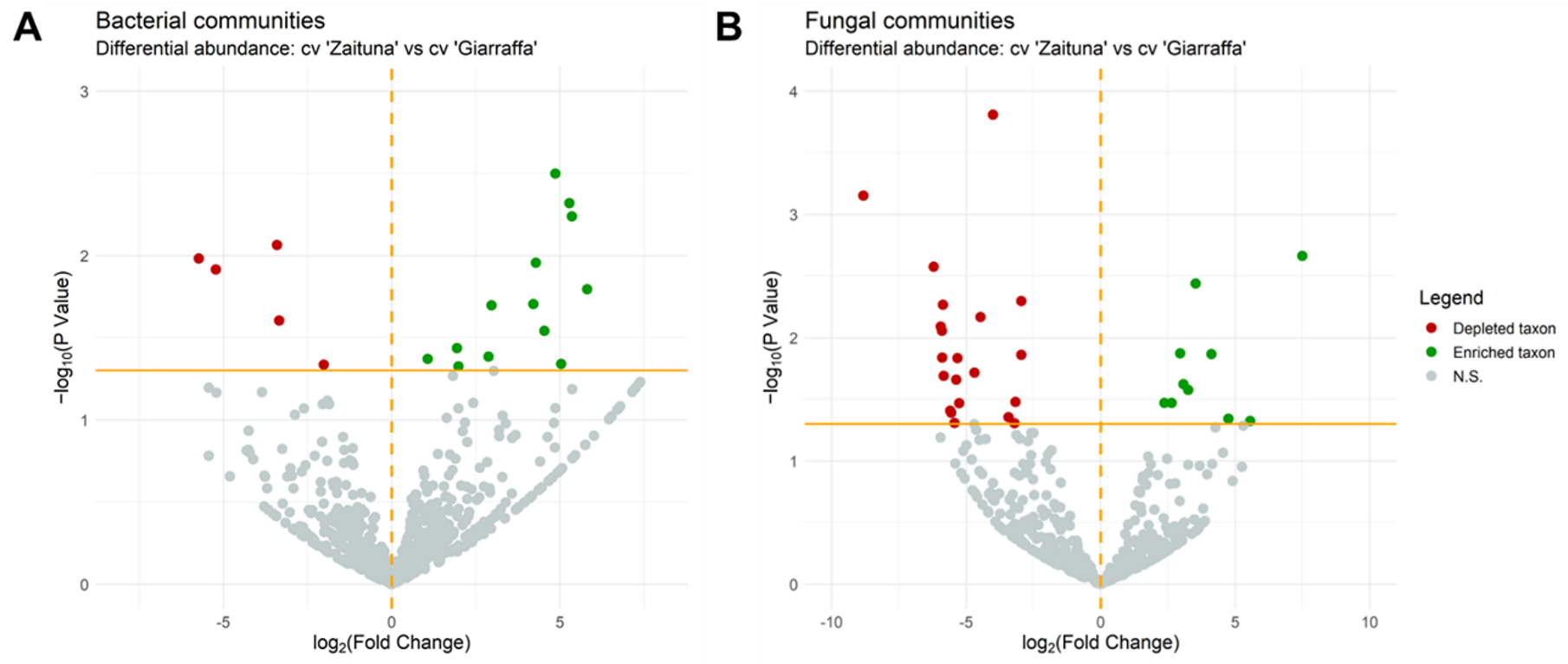
3.2. Endophytic Microbial Community in Susceptible and Resistant Cultivars
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schroth, M.N.; Osgood, J.W.; Miller, T.D. Quantitative assessment of the effect of the olive knot disease on olive yield and quality. Phytopathology 1973, 63, 1064–1065. [Google Scholar] [CrossRef]
- Wilson, E.E. The Olive knot disease: Its inception, development, and control. Hilgardia 1935, 9, 234–266. [Google Scholar] [CrossRef]
- Gardan, L.; Bollet, C.; Ghorrah, M.A.; Grimont, F.; Grimont, P.A.D. DNA Relatedness among the pathovar strains of Pseudomonas syringae subsp. savastanoi Janse (1982) and proposal of Pseudomonas savastanoi sp. nov. Int. J. Syst. Evol. Microbiol. 1992, 42, 606–612. [Google Scholar] [CrossRef]
- Quesada, J.M.; Penyalver, R.; López, M.M. Epidemiology and control of plant diseases caused by phytopathogenic bacteria: The case of olive knot disease caused by Pseudomonas savastanoi pv. savastanoi. In Plant Pathology; Cumagun, C.J., Ed.; Intech: Kayl, Luxembourg, 2012; pp. 299–326. [Google Scholar]
- Quesada, J.M.; Penyalver, R.; Pérez-Panadés, J.; Salcedo, C.I.; Carbonell, E.A.; López, M.M. Dissemination of Pseudomonas savastanoi pv. savastanoi populations and subsequent appearance of olive knot disease. Plant Pathol. 2010, 59, 262–269. [Google Scholar] [CrossRef]
- Lavermicocca, P.; Surico, G. Presenza epifitica di Pseudomonas syringae pv. savastanoi e di altri batteri sull’Olivo e sull’Oleandro. Phytopathol. Mediterr. 1987, 26, 137–141. [Google Scholar]
- Teviotdale, B.L.; Krueger, W.H. Effects of timing of copper sprays, defoliation, rainfall, and inoculum concentration on incidence of olive knot disease. Plant Dis. 2004, 88, 131–135. [Google Scholar] [CrossRef]
- Larcher, W. Temperature stress and survival ability of Mediterranean sclerophyllous plants. Plant Biosyst. 2000, 134, 279–295. [Google Scholar] [CrossRef]
- Ercolani, G.L. Pseudomonas savastanoi and other bacteria colonizing the surface of olive leaves in the field. Microbiology 1978, 109, 245–257. [Google Scholar] [CrossRef]
- Quesada, J.M.; Garcia, A.; Bertolini, E.; López, M.M.; Penyalver, R. Recovery of Pseudomonas savastanoi pv. savastanoi from symptomless shoots of naturally infected olive trees. Int. Microbiol. 2007, 10, 77. [Google Scholar]
- Hosni, T.; Moretti, C.; Devescovi, G.; Suarez-Moreno, Z.R.; Fatmi, M.; Guarnaccia, C.; Pongor, S.; Onofri, A.; Buonaurio, R.; Venturi, V. Sharing of quorum-sensing signals and role of interspecies communities in a bacterial plant disease. ISME J. 2011, 5, 1857–1870. [Google Scholar] [CrossRef]
- Buonaurio, R.; Moretti, C.; da Silva, D.P.; Cortese, C.; Ramos, C.; Venturi, V. The olive knot disease as a model to study the role of interspecies bacterial communities in plant disease. Front. Plant Sci. 2015, 6, 434. [Google Scholar] [CrossRef] [PubMed]
- European Commission Commission. Implementing Regulation (EU) 2018/785 of 29 May 2018 amending Implementing Regulation (EU) No 540/2011 as regards the conditions of approval of the active substance thiamethoxam. Off. J. Eur. Union. 2015, 132, 40–44. [Google Scholar]
- Sisto, A.; Iacobellis, N.S. Olive knot disease: Pathogenic and epidemiological aspects and defence strategies. Olivo Olio 1999, 2, 32–38. [Google Scholar]
- Young, J.M. Olive knot and its pathogens. Australas. Plant Pathol. 2004, 33, 33–39. [Google Scholar] [CrossRef]
- Hassani, D.; Buonaurio, R.; Tombesi, A. Response of some olive cultivars, hybrid and open pollinated seedlings to Pseudomonas savastanoi pv. savastanoi. In Pseudomonas syringae and Related Pathogens; Springer: Berlin/Heidelberg, Germany, 2003; pp. 489–494. [Google Scholar]
- Marcelo, A.; Fernandes, M.; Fatima Potes, M.; Serrano, J.F. Reactions of some cultivars of Olea europaea L. to experimental inoculation with Pseudomonas syringae pv. savastanoi. In Proceedings of the III International Symposium on Olive Growing, Manila, Philippines, 6–8 October 1997; Volume 474, pp. 581–584. [Google Scholar]
- Varvaro, L.; Surico, G. Behaviour of different olive (Olea europaea L.) cultivars to inoculation with Pseudomonas savastanoi (EF Smith) Stevens. Phytopathol. Mediterr. 1978, 17, 174–177. [Google Scholar]
- Penyalver, R.; García, A.; Ferrer, A.; Bertolini, E.; Quesada, J.M.; Salcedo, C.I.; Piquer, J.; Pérez-Panadés, J.; Carbonell, E.A.; Del Rio, C. Factors affecting Pseudomonas savastanoi pv. savastanoi plant inoculations and their use for evaluation of olive cultivar susceptibility. Phytopathology 2006, 96, 313–319. [Google Scholar] [CrossRef]
- Catara, V.; Bella, P.; Tessitori, M.; Tirrò, A.; Colina, P. Variabilità di Pseudomonas savastanoi pv. savastanoi in un’area olivicola della Sicilia e comportamento di alcune varietà di olivo alle inoculazioni [Olea europaea L.]. Tec. Agric. 2005, 57, 41–52. [Google Scholar]
- Salman, M.; Greenhut, R.; Preece, J.; Ferguson, L.; Kluepfel, D. Field evaluation of olive (Olea europaea) genotypes for resistance to Pseudomonas savastanoi pv. savastanoi. J. Plant Pathol. 2020, 102, 663–670. [Google Scholar] [CrossRef]
- Valverde, P.; Zucchini, M.; Polverigiani, S.; Lodolini, E.M.; López-Escudero, F.J.; Neri, D. Olive knot damages in ten olive cultivars after late-winter frost in central Italy. Sci. Hortic. 2020, 266, 109274. [Google Scholar] [CrossRef]
- Mancuso, S. Electrical resistance changes during exposure to low temperature measure chilling and freezing tolerance in olive tree (Olea europaea L.) plants. Plant. Cell Environ. 2000, 23, 291–299. [Google Scholar] [CrossRef]
- Petruccelli, R.; Bartolini, G.; Ganino, T.; Zelasco, S.; Lombardo, L.; Perri, E.; Durante, M.; Bernardi, R. Cold stress, freezing adaptation, varietal susceptibility of Olea europaea L.: A Review. Plants 2022, 11, 1367. [Google Scholar] [CrossRef] [PubMed]
- Lodolini, E.M.; Alfei, B.; Santinelli, A.; Cioccolanti, T.; Polverigiani, S.; Neri, D. Frost tolerance of 24 olive cultivars and subsequent vegetative re-sprouting as indication of recovery ability. Sci. Hortic. 2016, 211, 152–157. [Google Scholar] [CrossRef]
- Bass, D.; Stentiford, G.D.; Wang, H.-C.; Koskella, B.; Tyler, C.R. The pathobiome in animal and plant diseases. Trends Ecol. Evol. 2019, 34, 996–1008. [Google Scholar] [CrossRef] [PubMed]
- Lata, R.; Chowdhury, S.; Gond, S.K.; White, J.F., Jr. Induction of abiotic stress tolerance in plants by endophytic microbes. Lett. Appl. Microbiol. 2018, 66, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zhu, J.; Zhao, X.; Shi, J.; Jiang, C.; Shao, D. Beneficial effects of endophytic fungi colonization on plants. Appl. Microbiol. Biotechnol. 2019, 103, 3327–3340. [Google Scholar] [CrossRef]
- Nicoletti, R.; Di Vaio, C.; Cirillo, C. Endophytic fungi of olive tree. Microorganisms 2020, 8, 1321. [Google Scholar] [CrossRef]
- Gomes, T.; Pereira, J.A.; Lino-Neto, T.; Bennett, A.E.; Baptista, P. Bacterial disease induced changes in fungal communities of olive tree twigs depend on host genotype. Sci. Rep. 2019, 9, 5882. [Google Scholar] [CrossRef]
- Mina, D.; Pereira, J.A.; Lino-Neto, T.; Baptista, P. Impact of plant genotype and plant habitat in shaping bacterial pathobiome: A comparative study in olive tree. Sci. Rep. 2020, 10, 3475. [Google Scholar] [CrossRef]
- Giampetruzzi, A.; Baptista, P.; Morelli, M.; Cameirão, C.; Lino Neto, T.; Costa, D.; D’Attoma, G.; Abou Kubaa, R.; Altamura, G.; Saponari, M. Differences in the endophytic microbiome of olive cultivars infected by Xylella fastidiosa across seasons. Pathogens 2020, 9, 723. [Google Scholar] [CrossRef]
- Pyrowolakis, E.; Weltzien, H.C. Studies on the distribution of Olive knot, induced by Pseudomonas savastanoi (Sm.) Stev. in the Greek island of Crete. Phytopathol. Mediterr. 1974, 13, 118–120. [Google Scholar]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar]
- Penyalver, R.; García, A.; Ferrer, A.; Bertolini, E.; López, M.M. Detection of Pseudomonas savastanoi pv. savastanoi in olive plants by enrichment and PCR. Appl. Environ. Microbiol. 2000, 66, 2673–2677. [Google Scholar] [CrossRef]
- Bella, P.; Licciardello, G.; Tessitori, M.; Catara, V. A real-time PCR quantitative detection assay for Pseudomonas savastanoi pv. nerii in Nerium oleander. Phytopathol. Mediterr. 2008, 47, 204–213. [Google Scholar]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Paytuví, A.; Battista, E.; Scippacercola, F.; Cigliano, R.A.; Sanseverino, W. GAIA: An integrated metagenomics suite. bioRxiv 2019, 804690. [Google Scholar]
- R Core Team. R Development Core Team. R A Lang. Environ. Stat. Comput. 2016, 55, 275–286. [Google Scholar]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Love, M.; Anders, S.; Huber, W. Differential analysis of count data–the DESeq2 package. Genome Biol. 2014, 15, 10–1186. [Google Scholar]
- Wickham, H. Data analysis. In ggplot2; Springer: Berlin/Heidelberg, Germany, 2016; pp. 189–201. [Google Scholar]
- Weiß, C.H. StatSoft, Inc., Tulsa, Ok.: Statistica, Version 8. AStA Adv. Stat. Anal. 2007, 91, 339–341. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Package ‘factoextra’. Extr. Vis. Results Multivar. Data Anal. 2017, 76. [Google Scholar]
- Las Casas, G.; Scollo, F.; Distefano, G.; Continella, A.; Gentile, A.; La Malfa, S. Molecular characterization of olive (Olea europaea L.) Sicilian cultivars using SSR markers. Biochem. Syst. Ecol. 2014, 57, 15–19. [Google Scholar] [CrossRef]
- Mantia, L.; Lain, T.; Caruso, T.; Testolin, R. SSR-based DNA fingerprints reveal the genetic diversity of Sicilian olive (Olea europaea L.) germplasm. J. Hortic. Sci. Biotechnol. 2005, 80, 628–632. [Google Scholar] [CrossRef]
- Bottari, V.; Spina, P. Le varietà di olivo coltivate in Sicilia. Ann. Ist. Sper. Agron. 1953, 7, 937–1004. [Google Scholar]
- Mousavi, S.; De la Rosa, R.; Moukhli, A.; El Riachy, M.; Mariotti, R.; Torres, M.; Pierantozzi, P.; Stanzione, V.; Mastio, V.; Zaher, H. Plasticity of fruit and oil traits in olive among different environments. Sci. Rep. 2019, 9, 16968. [Google Scholar] [CrossRef] [Green Version]
- Licciardello, G.; Moral, J.; Strano, M.C.; Caruso, P.; Sciara, M.; Bella, P.; Sorrentino, G.; Di Silvestro, S. Characterization of Colletotrichum strains associated with olive anthracnose in Sicily. Phytopathol. Mediterr. 2022, 61, 139–151. [Google Scholar] [CrossRef]
- Tugizimana, F.; Mhlongo, M.I.; Piater, L.A.; Dubery, I.A. Metabolomics in plant priming research: The way forward? Int. J. Mol. Sci. 2018, 19, 1759. [Google Scholar] [CrossRef]
- Moral, J.; De la Rosa, R.; León, L.; Barranco, D.; Michailides, T.J.; Trapero, A. High susceptibility of olive cultivar FS-17 to Alternaria alternata in Southern Spain. Plant Dis. 2008, 92, 1252. [Google Scholar] [CrossRef]
- Basım, E.; Basım, H.; Abdulai, M.; Baki, D.; Öztürk, N. Identification and characterization of Alternaria alternata causing leaf spot of olive tree (Olea europaea) in Turkey. Crop Prot. 2017, 92, 79–88. [Google Scholar] [CrossRef]
- Tziros, G.T.; Karpouzis, A.; Lagopodi, A.L. Alternaria alternata as the cause of decline and necrosis on olive tree cuttings in Greece. Australas. Plant Dis. Notes 2021, 16, 7. [Google Scholar] [CrossRef]
- Berardo, C.; Bulai, I.M.; Venturino, E.; Baptista, P.; Gomes, T. Modeling the endophytic fungus Epicoccum nigrum action to fight the “olive knot” disease caused by Pseudomonas savastanoi pv. savastanoi (Psv) bacteria in Olea europaea L. Trees. Trends Biomath. Model. Optim. Comput. Probl. 2018, 189–207. [Google Scholar]
- Hanani, A.; Valentini, F.; Sanzani, S.M.; Santoro, F.; Minutillo, S.A.; Gallo, M.; Cavallo, G.; Mourou, M.; El Moujabber, M.; D’Onghia, A.M. Community analysis of culturable sapwood endophytes from apulian olive varieties with different susceptibility to Xylella fastidiosa. Agronomy 2021, 12, 9. [Google Scholar] [CrossRef]
- Tuppad, D.S.; Shishupala, S. Evaluation of endophytic fungi from Butea monosperma for antimicrobial and enzyme activity. J. Med. Plants Stud. 2014, 2, 38–45. [Google Scholar]
- Frey-Klett, P.; Burlinson, P.; Deveau, A.; Barret, M.; Tarkka, M.; Sarniguet, A. Bacterial-fungal interactions: Hyphens between agricultural, clinical, environmental, and food microbiologists. Microbiol. Mol. Biol. Rev. 2011, 75, 583–609. [Google Scholar] [CrossRef]
- Valenzuela-Lopez, N.; Cano-Lira, J.F.; Stchigel, A.M.; Rivero-Menendez, O.; Alastruey-Izquierdo, A.; Guarro, J. Neocucurbitaria keratinophila: An emerging opportunistic fungus causing superficial mycosis in Spain. Med. Mycol. 2019, 57, 733–738. [Google Scholar] [CrossRef]
- Jayawardena, R.S.; Ariyawansa, H.A.; Singtripop, C.; Li, Y.M.; Yan, J.; Li, X.; Nilthong, S.; Hyde, K.D. A re-assessment of Elsinoaceae (Myriangiales, Dothideomycetes). Phytotaxa 2014, 176, 120–138. [Google Scholar] [CrossRef]






| OTUs | Phylum | Class | Order | Family | Genus |
|---|---|---|---|---|---|
| 16S | 34 | 65 | 141 | 286 | 678 |
| ITS | 5 | 22 | 64 | 144 | 292 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Licciardello, G.; Mosca, A.; Di Silvestro, S.; Puglisi, D.; Russo, M.P.; Catara, V.; Caruso, P. Cultivar Susceptibility to Olive Knot Disease and Association with Endophytic Microbiota Community. Agronomy 2023, 13, 468. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13020468
Licciardello G, Mosca A, Di Silvestro S, Puglisi D, Russo MP, Catara V, Caruso P. Cultivar Susceptibility to Olive Knot Disease and Association with Endophytic Microbiota Community. Agronomy. 2023; 13(2):468. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13020468
Chicago/Turabian StyleLicciardello, Grazia, Alexandros Mosca, Silvia Di Silvestro, Damiano Puglisi, Maria Patrizia Russo, Vittoria Catara, and Paola Caruso. 2023. "Cultivar Susceptibility to Olive Knot Disease and Association with Endophytic Microbiota Community" Agronomy 13, no. 2: 468. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13020468