
Effect of Biostimulants on Leafy Vegetables (Baby Leaf Lettuce and Batavia Lettuce) Exposed to Abiotic or Biotic Stress under Two Different Growing Systems
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pythium ultimum
2.2. Plant Material and Growth Conditions
2.3. Biostimulants
2.4. Effect of Biostimulants on Baby Leaf Lettuce Grown in a Substrate Colonized with P. ultimum
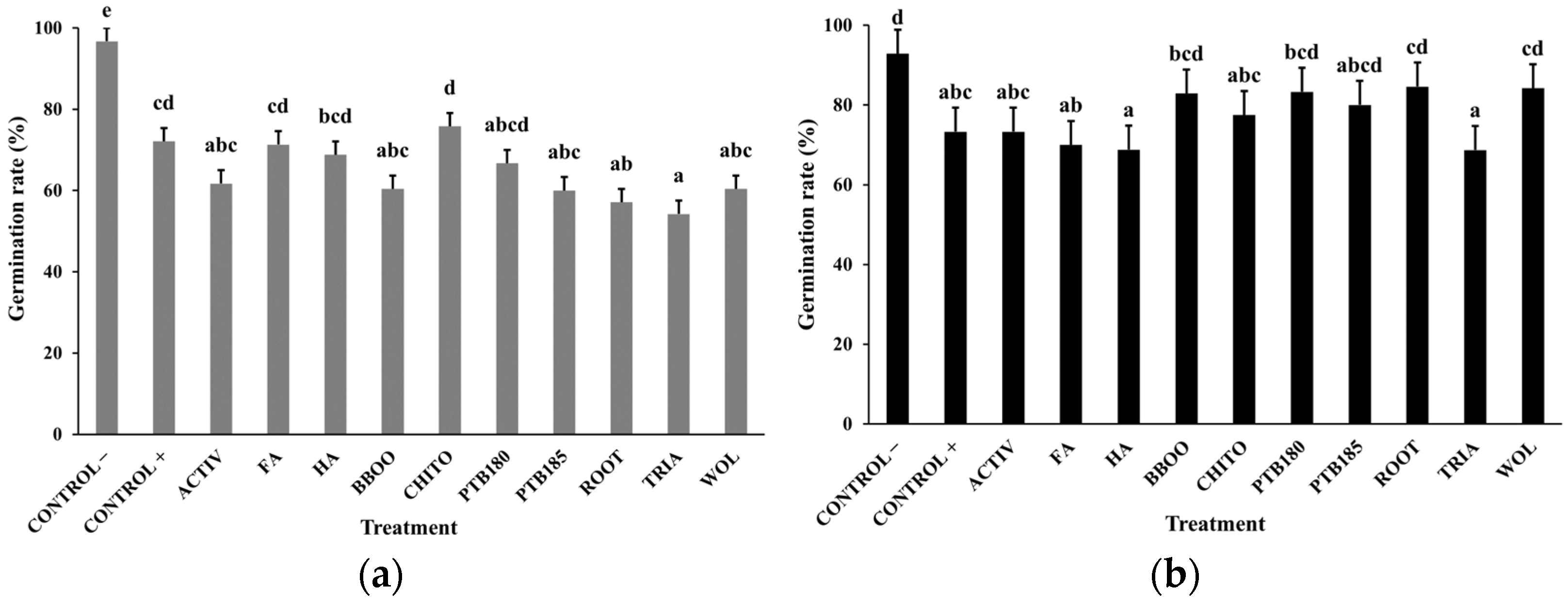
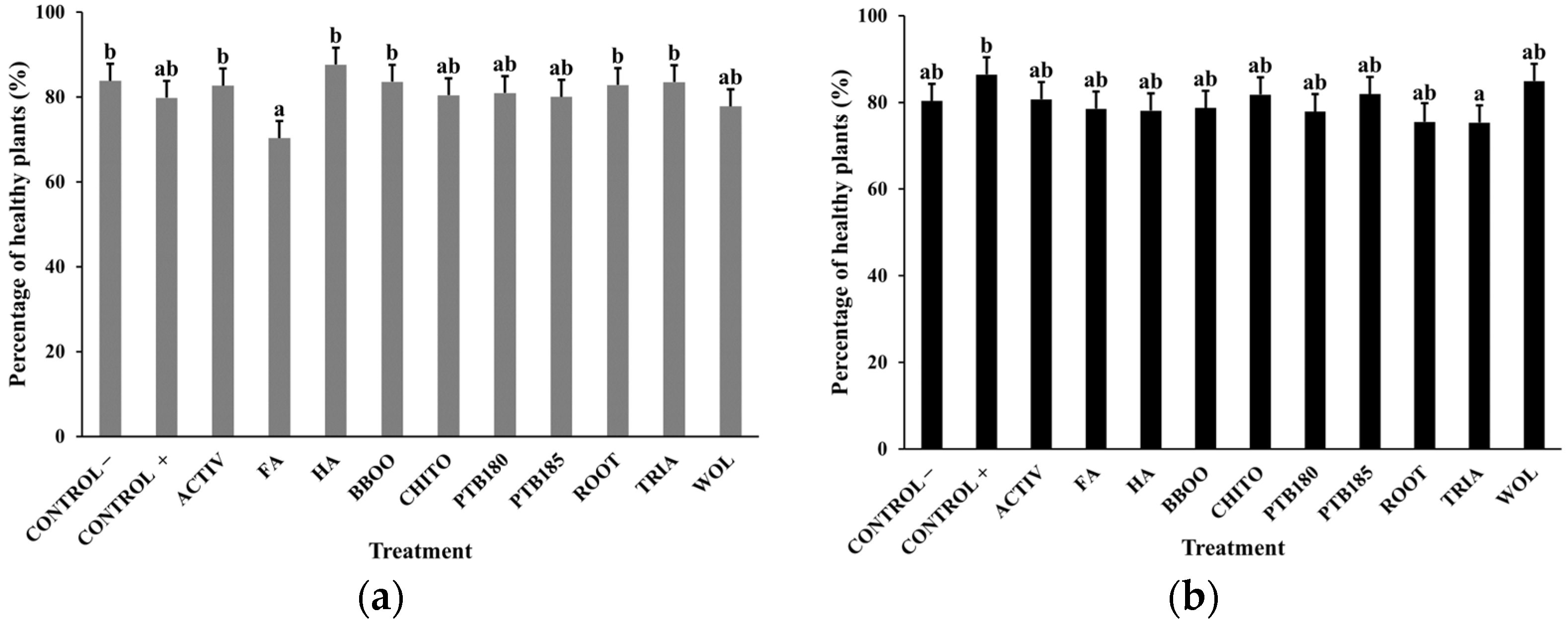
2.4.1. Germination Rate and Percentage of Healthy Plants
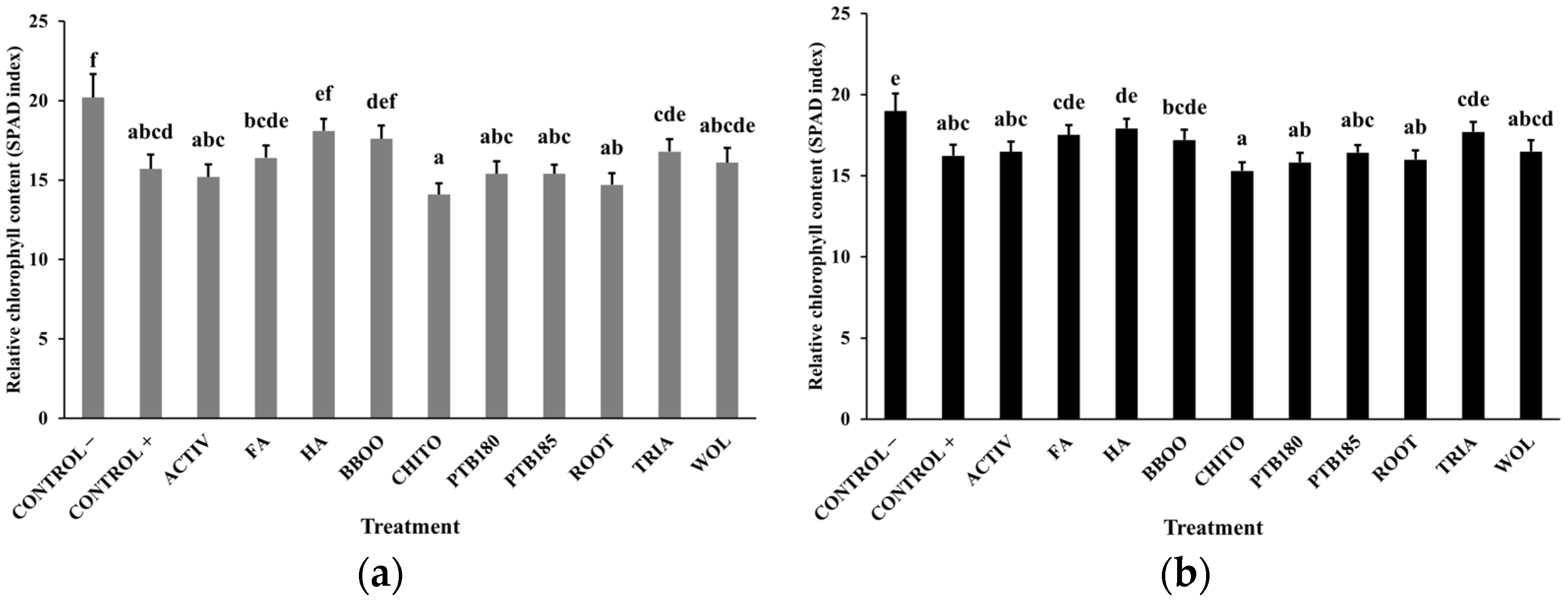
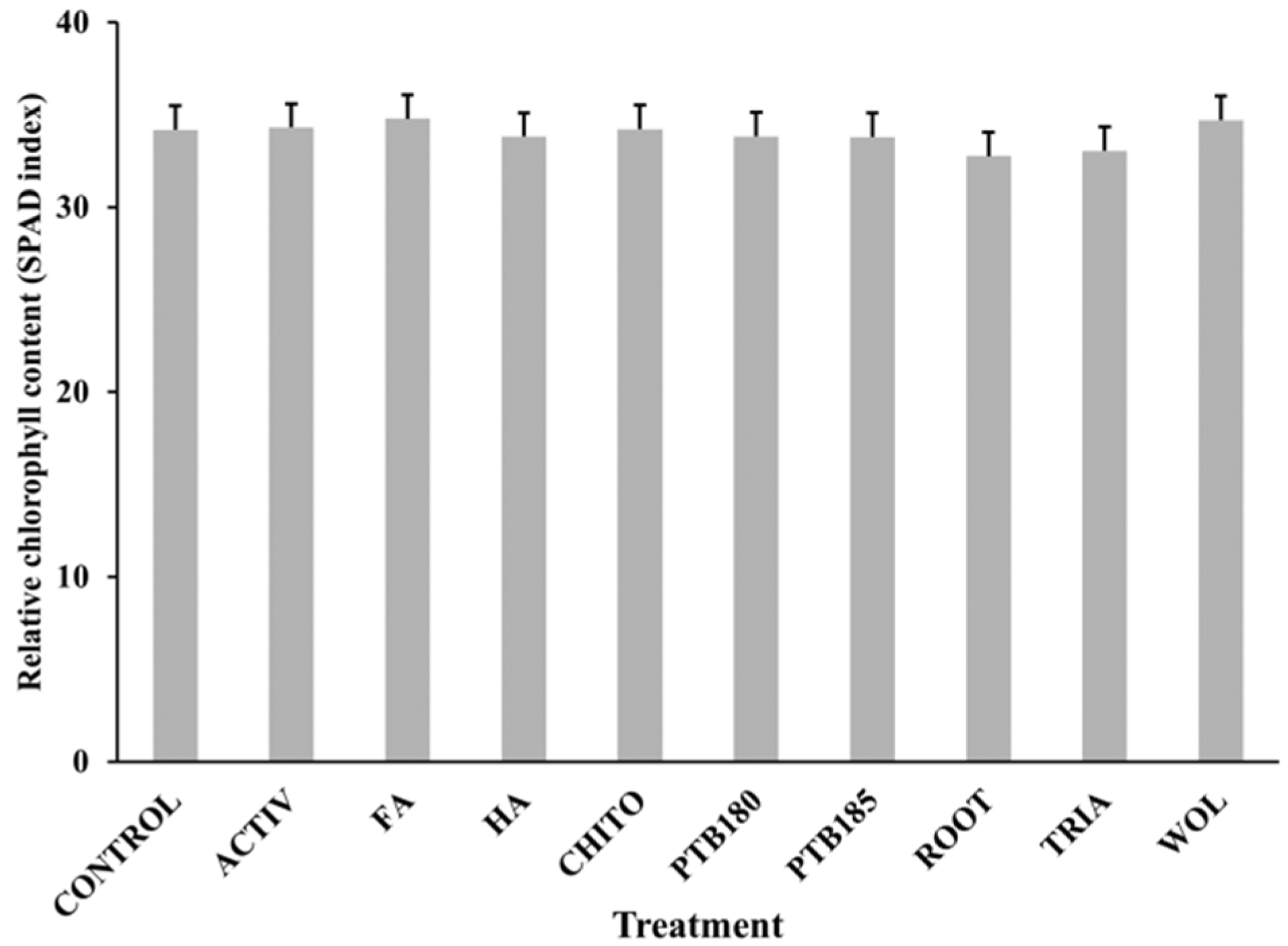
2.4.2. Chlorophyll Content (SPAD Index)
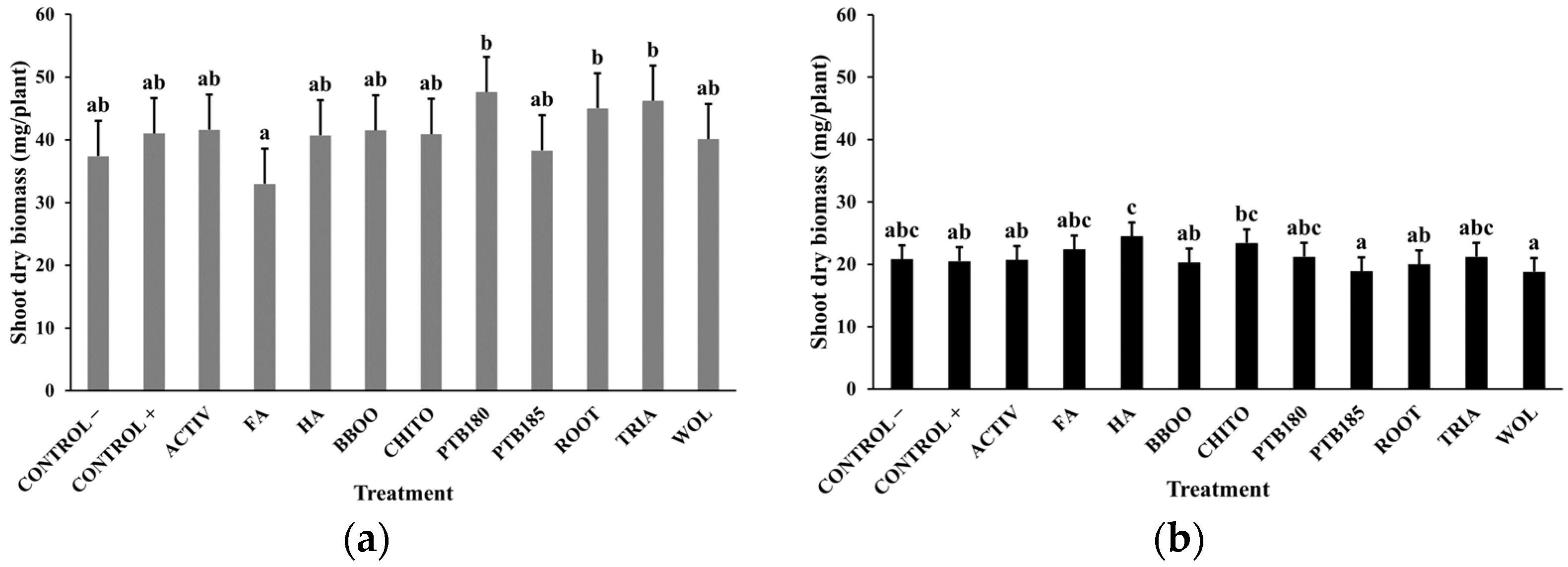
2.4.3. Shoot Dry Biomass
2.5. Effect of Biostimulants on Baby Leaf Lettuce Grown under Salinity Stress
2.6. Effect of Biostimulants on Lettuce Grown under Water Stress
2.7. Statistical Analysis
3. Results
3.1. Effect of Biostimulants on Baby Leaf Lettuce Grown in a Substrate Colonized with P. ultimum
3.2. Effect of Biostimulants on Baby Leaf Lettuce Grown under Salinity Stress
3.3. Effect of Biostimulants on Batavia Lettuce Grown under Water Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The Role of Biostimulants and Bioeffectors as Alleviators of Abiotic Stress in Crop Plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Del Buono, D. Can Biostimulants Be Used to Mitigate the Effect of Anthropogenic Climate Change on Agriculture? It Is Time to Respond. Sci. Total Environ. 2021, 751, 141763. [Google Scholar] [CrossRef] [PubMed]
- du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and Fulvic Acids as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein Hydrolysates as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Barrada, A.; Delisle-Houde, M.; Nguyen, T.T.A.; Tweddell, R.J.; Dorais, M. Drench Application of Soy Protein Hydrolysates Increases Tomato Plant Fitness, Fruit Yield, and Resistance to a Hemibiotrophic Pathogen. Agronomy 2022, 12, 1761. [Google Scholar] [CrossRef]
- Sharma, H.S.S.; Fleming, C.; Selby, C.; Rao, J.R.; Martin, T. Plant Biostimulants: A Review on the Processing of Macroalgae and Use of Extracts for Crop Management to Reduce Abiotic and Biotic Stresses. J. Appl. Phycol. 2014, 26, 465–490. [Google Scholar] [CrossRef]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed Extracts as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum-Based Biostimulants: Sustainable Applications in Agriculture for the Stimulation of Plant Growth, Stress Tolerance, and Disease Management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Ruiz, J.; Arnao, M.B. Relationship of Melatonin and Salicylic Acid in Biotic/Abiotic Plant Stress Responses. Agronomy 2018, 8, 33. [Google Scholar] [CrossRef] [Green Version]
- Zulfiqar, F.; Casadesús, A.; Brockman, H.; Munné-Bosch, S. An Overview of Plant-Based Natural Biostimulants for Sustainable Horticulture with a Particular Focus on Moringa Leaf Extracts. Plant Sci. 2020, 295, 110194. [Google Scholar] [CrossRef]
- Islam, S.; Zaid, A.; Mohammad, F. Role of Triacontanol in Counteracting the Ill Effects of Salinity in Plants: A Review. J. Plant Growth Regul. 2021, 40, 1–10. [Google Scholar] [CrossRef]
- El Hadrami, A.; Adam, L.R.; El Hadrami, I.; Daayf, F. Chitosan in Plant Protection. Mar. Drugs 2010, 8, 968–987. [Google Scholar] [CrossRef]
- Pichyangkura, R.; Chadchawan, S. Biostimulant Activity of Chitosan in Horticulture. Sci. Hortic. 2015, 196, 49–65. [Google Scholar] [CrossRef]
- Malerba, M.; Cerana, R. Chitosan Effects on Plant Systems. IJMS 2016, 17, 996. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Sun, W.; Zhu, Y.-G.; Christie, P. Mechanisms of Silicon-Mediated Alleviation of Abiotic Stresses in Higher Plants: A Review. Environ. Pollut. 2007, 147, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Merino, F.C.; Trejo-Téllez, L.I. Biostimulant Activity of Phosphite in Horticulture. Sci. Hortic. 2015, 196, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Franzoni, G.; Cocetta, G.; Prinsi, B.; Ferrante, A.; Espen, L. Biostimulants on Crops: Their Impact under Abiotic Stress Conditions. Horticulturae 2022, 8, 189. [Google Scholar] [CrossRef]
- Chérif, M.; Menzies, J.G.; Benhamou, N.; Bélanger, R.R. Studies of Silicon Distribution in Wounded and Pythium Ultimum Infected Cucumber Plants. Physiol. Mol. Plant Pathol. 1992, 41, 371–385. [Google Scholar] [CrossRef]
- Ma, J.F. Role of Silicon in Enhancing the Resistance of Plants to Biotic and Abiotic Stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Johri, B.N. Interactions of Bacillus spp. and Plants—With Special Reference to Induced Systemic Resistance (ISR). Microbiol. Res. 2009, 164, 493–513. [Google Scholar] [CrossRef]
- Sofo, A.; Nuzzaci, M.; Vitti, A.; Tataranni, G.; Scopa, A. Control of Biotic and Abiotic Stresses in Cultivated Plants by the Use of Biostimulant Microorganisms. In Improvement of Crops in the Era of Climatic Changes; Ahmad, P., Wani, M.R., Azooz, M.M., Tran, L.-S.P., Eds.; Springer: New York, NY, USA, 2014; pp. 107–117. ISBN 978-1-4614-8829-3. [Google Scholar]
- López-Bucio, J.; Pelagio-Flores, R.; Herrera-Estrella, A. Trichoderma as Biostimulant: Exploiting the Multilevel Properties of a Plant Beneficial Fungus. Sci. Hortic. 2015, 196, 109–123. [Google Scholar] [CrossRef]
- Ruzzi, M.; Aroca, R. Plant Growth-Promoting Rhizobacteria Act as Biostimulants in Horticulture. Sci. Hortic. 2015, 196, 124–134. [Google Scholar] [CrossRef]
- Grewal, A.; Abbey, L.; Gunupuru, L.R. Production, Prospects and Potential Application of Pyroligneous Acid in Agriculture. J. Anal. Appl. Pyrolysis 2018, 135, 152–159. [Google Scholar] [CrossRef]
- Gull, A.; Lone, A.A.; Wani, N.U.I. Biotic and Abiotic Stresses in Plants. In Abiotic and Biotic Stress in Plants; Bosco de Oliveira, A., Ed.; IntechOpen: London, UK, 2019; pp. 3–8. ISBN 978-1-78932-811-2. [Google Scholar]
- Subbarao, K.V.; Davis, R.M.; Gilbertson, R.L.; Raid, R.N. Compendium of Lettuce Diseases and Pests, 2nd ed.; The American Phytopathological Society: Saint Paul, MN, USA, 2017; pp. 31–45. ISBN 978-0-89054-578-2. [Google Scholar]
- Tziros, G.T.; Karaoglanidis, G.S. Molecular Identification and Pathogenicity of Rhizoctonia Solani and Pythium Spp. Associated with Damping-off Disease on Baby Leafy Vegetables in Greece. Plant Pathol. 2022, 71, 1381–1391. [Google Scholar] [CrossRef]
- Gullino, M.L.; Gilardi, G.; Garibaldi, A. Ready-to-Eat Salad Crops: A Plant Pathogen’s Heaven. Plant Dis. 2019, 103, 2153–2170. [Google Scholar] [CrossRef] [Green Version]
- Machado, R.M.A.; Serralheiro, R.P. Soil Salinity: Effect of Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Khalid, M.F.; Huda, S.; Yong, M.; Li, L.; Li, L.; Chen, Z.-H.; Ahmed, T. Alleviation of Drought and Salt Stress in Vegetables: Crop Responses and Mitigation Strategies. Plant Growth Regul. 2023, 99, 177–194. [Google Scholar] [CrossRef]
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, E.; Blaszczak, A.; Wiatrak, P.; Canady, M. Biostimulant Mode of Action. In The Chemical Biology of Plant Biostimulants; Geelen, D., Xu, L., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 205–227. ISBN 978-1-119-35725-4. [Google Scholar]
- van der Plaats-Niterink, A.J. Monograph of the Genus. Pythium. Stud. Mycol. 1981, 21, 1–244. [Google Scholar]
- Bouchard-Rochette, M.; Machrafi, Y.; Cossus, L.; Nguyen, T.T.A.; Antoun, H.; Droit, A.; Tweddell, R.J. Bacillus pumilus PTB180 and Bacillus subtilis PTB185: Production of Lipopeptides, Antifungal Activity, and Biocontrol Ability against Botrytis cinerea. Biol. Control. 2022, 170, 104925. [Google Scholar] [CrossRef]
- Dembélé, D.M. Production de Verdurettes Biologiques: Niveaux de Fertilisation et Biostimulants. Master’s Thesis, Université Laval, Québec, QC, Canada, 2021. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar] [CrossRef]
- Giménez, A.; Fernández, J.A.; Pascual, J.A.; Ros, M.; Egea-Gilabert, C. Application of Directly Brewed Compost Extract Improves Yield and Quality in Baby Leaf Lettuce Grown Hydroponically. Agronomy 2020, 10, 370. [Google Scholar] [CrossRef] [Green Version]
- Forotaghe, Z.A.; Souri, M.K.; Jahromi, M.G.; Torkashvand, A.M. Influence of Humic Acid Application on Onion Growth Characteristics under Water Deficit Conditions. J. Plant Nutr. 2022, 45, 1030–1040. [Google Scholar] [CrossRef]
- Hassanein, R.A.; Hussein, O.S.; Farag, I.A.; Hassan, Y.E.; Abdelkader, A.F.; Ibrahim, M. Salt-Stressed Coriander (Coriandrum sativum L.) Responses to Potassium Silicate, Humic Acid and Gamma Irradiation Pretreatments. Agronomy 2022, 12, 2268. [Google Scholar] [CrossRef]
- Turan, M.; Ekinci, M.; Kul, R.; Kocaman, A.; Argin, S.; Zhirkova, A.M.; Perminova, I.V.; Yildirim, E. Foliar Applications of Humic Substances Together with Fe/Nano Fe to Increase the Iron Content and Growth Parameters of Spinach (Spinacia oleracea L.). Agronomy 2022, 12, 2044. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, R.; Liang, Y.; Zhang, S.; Zhang, Z.; Sun, C.; Li, J.; Qi, Z.; Yang, Q. Comparing Efficacy of Different Biostimulants for Hydroponically Grown Lettuce (Lactuca sativa L.). Agronomy 2022, 12, 786. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Gangwar, S.; Prasad, S.M.; Maurya, J.N.; Chauhan, D.K. Role of Silicon in Enrichment of Plant Nutrients and Protection from Biotic and Abiotic Stresses. In Improvement of Crops in the Era of Climatic Changes; Ahmad, P., Wani, M.R., Azooz, M.M., Tran, L.-S.P., Eds.; Springer: New York, NY, USA, 2014; pp. 39–56. ISBN 978-1-4614-8829-3. [Google Scholar]
- Verma, K.K.; Song, X.-P.; Zeng, Y.; Guo, D.-J.; Singh, M.; Rajput, V.D.; Malviya, M.K.; Wei, K.-J.; Sharma, A.; Li, D.-P.; et al. Foliar Application of Silicon Boosts Growth, Photosynthetic Leaf Gas Exchange, Antioxidative Response and Resistance to Limited Water Irrigation in Sugarcane (Saccharum officinarum L.). Plant Physiol. Biochem. 2021, 166, 582–592. [Google Scholar] [CrossRef]
- Chaski, C.; Petropoulos, S.A. The Alleviation Effects of Biostimulants Application on Lettuce Plants Grown under Deficit Irrigation. Horticulturae 2022, 8, 1089. [Google Scholar] [CrossRef]
- Avis, T.J.; Gravel, V.; Antoun, H.; Tweddell, R.J. Multifaceted Beneficial Effects of Rhizosphere Microorganisms on Plant Health and Productivity. Soil Biol. Biochem. 2008, 40, 1733–1740. [Google Scholar] [CrossRef]
- Tarigan, S.I.; Toth, S.; Szalai, M.; Kiss, J.; Turoczi, G.; Toepfer, S. Biological Control Properties of Microbial Plant Biostimulants. A Review. Biocontrol Sci. Technol. 2022, 32, 1351–1371. [Google Scholar] [CrossRef]
- Xu, C.; Leskovar, D.I. Effects of A. nodosum Seaweed Extracts on Spinach Growth, Physiology and Nutrition Value under Drought Stress. Sci. Hortic. 2015, 183, 39–47. [Google Scholar] [CrossRef]
- Guinan, K.J.; Sujeeth, N.; Copeland, R.B.; Jones, P.W.; O’Brien, N.M.; Sharma, H.S.S.; Prouteau, P.F.J.; O’Sullivan, J.T. Discrete Roles for Extracts of Ascophyllum nodosum in Enhancing Plant Growth and Tolerance to Abiotic and Biotic Stresses. Acta Hortic. 2013, 127–135. [Google Scholar] [CrossRef]
- Rose, M.T.; Patti, A.F.; Little, K.R.; Brown, A.L.; Jackson, W.R.; Cavagnaro, T.R. A Meta-Analysis and Review of Plant-Growth Response to Humic Substances: Practical Implications for Agriculture. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2014; Volume 124, pp. 37–89. ISBN 978-0-12-800138-7. [Google Scholar]
- Gómez, S.; Gómez, C. Evaluating the Use of Biostimulants for Indoor Hydroponic Lettuce Production. HortTechnology 2022, 32, 348–355. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic Fertilizer [Manufacturer] | Rate for Baby Leaf Lettuce (g L−1 of Substrate) | Rate for Batavia Lettuce (g L−1 of Substrate) |
|---|---|---|
| Pure hen manure pellets 5-3-2 (Acti-Sol inc., Notre-Dame-du-Bon-Conseil, QC, Canada) | 4.8 | 6.8 |
| Feather meal 13-0-0 (Les Engrais Naturels McInnes, Stanstead, QC, Canada) | 2.5 | 1.3 |
| Potassium sulfate (Tessenderlo Group N.V., Troonstraat, Belgium) | 0.7 | 0.9 |
| Epsom salt (EPSO Top®, K + S, Kassel, Germany) | 0.3 | 0.1 |
| Nutrient | Concentration for Baby Leaf Lettuce (mg L−1) | Concentration for Batavia Lettuce (mg L−1) |
|---|---|---|
| N-NO3 | 200 | 255.6 |
| N-NH4 | 10 | 14.8 |
| P | 50 | 80.6 |
| K | 400 | 437.7 |
| Ca | 200 | 208.6 |
| Mg | 55.4 | 40.8 |
| S | 135.1 | 179.5 |
| Fe | 6 | 0.8 |
| Mn | 1 | 0.5 |
| Zn | 0.5 | 0.1 |
| B | 0.5 | 0.2 |
| Cu | 0.1 | 0.2 |
| Mo | 0.1 | 0.03 |
| Product Name (Active Ingredient) | Manufacturer | Concentration for Drench Application (per L of Water) | Concentration for Seed Priming (per L of Water) |
|---|---|---|---|
| ACTIV 0-0-5 (Ascophyllum nodosum extract) | OrganicOcean Inc. [Rimouski, QC, Canada] | 0.6 mL | 0.03 mL |
| Organic BamBoo Power BBP No. 11 (bamboo vinegar) | Seek Bio-Technology [Shanghai, China] | 1.25 mL | - |
| EZ-Gro Chitosan (4% chitosan) | EZ-Gro Inc. [Kingston, ON, Canada] | 20 mL | 0.007 mL |
| Fulvic Acid 0-0-3 (70% fulvic acid) | EZ-Gro Inc. | 0.144 g | 50 g |
| Humic Acid 0-0-5 (80% humic acid) | EZ-Gro Inc. | 0.24 g | 50 g |
| PTB180 (Bacillus pumilus PTB180) | Premier Tech [Rivière-du-Loup, QC, Canada] | 1 × 1010 CFU a | 1 × 1010 CFU |
| PTB185 (Bacillus subtilis PTB185) | Premier Tech | 1 × 1010 CFU | 1 × 1010 CFU |
| RootShield® (Trichoderma harzianum T-22) | BioWorks® [Victor, NY, USA] | 1 × 107 CFU | 1 × 108 CFU |
| Nutri-Stim Triacontanol™ (2.5% triacontanol) | Nutri-Tech Solutions® [Yandina, Australia] | 0.004 mL | 0.004 mL |
| Wollastonite (calcium silicate; 55% SiO2) | Canadian Wollastonite [Seeleys Bay, ON, Canada] | 4 g | 4 g |
| Germination Rate (%) | ||||
|---|---|---|---|---|
| 0 mM NaCl L−1 | 40 mM NaCl L−1 | 80 mM NaCl L−1 | 120 mM NaCl L−1 | |
| CONTROL | 75.8 a | 77.5 ab | 54.3 ab | 16.8 ab |
| ACTIV | 80.0 a | 67.3 ab | 50.0 ab | 20.0 ab |
| FA | 84.2 a | 74.3 ab | 63.3 bc | 20.0 ab |
| HA | 82.3 a | 84.3 b | 62.5 abc | 49.3 c |
| CHITO | 76.8 a | 63.3 a | 47.5 ab | 22.5 ab |
| PTB180 | 81.8 a | 60.8 a | 43.3 a | 7.5 a |
| PTB185 | 80.8 a | 76.8 ab | 51.0 ab | 15.3 ab |
| ROOT | 79.3 a | 83.5 b | 76.8 c | 43.5 c |
| TRIA | 80.8 a | 67.0 ab | 62.8 abc | 31.0 bc |
| TRIA † | 68.3 a | 85.0 b | 74.3 c | 74.0 d |
| WOL | 78.3 a | 67.3 ab | 66.8 bc | 34.3 bc |
| WOL † | 85.0 a | 84.3 b | 76.8 c | 70.0 d |
| Germination Rate (%) | ||||
|---|---|---|---|---|
| 0 mM NaCl L−1 | 40 mM NaCl L−1 | 80 mM NaCl L−1 | 120 mM NaCl L−1 | |
| CONTROL | 51.0 ab | 21.8 abc | 7.5 a | 0.8 a |
| ACTIV | 63.5 abc | 29.3 abc | 2.5 a | 1.5 a |
| FA | 45.0 a | 11.8 a | 6.8 a | 0.0 a |
| HA | 64.5 abc | 39.3 cd | 14.0 a | 0.8 a |
| CHITO | 60.0 abc | 29.3 abc | 4.8 a | 4.3 a |
| PTB180 | 55.0 ab | 17.5 ab | 3.3 a | 0.0 a |
| PTB185 | 67.5 bcd | 29.0 abc | 7.3 a | 0.8 a |
| ROOT | 60.5 abc | 53.3 de | 16.0 a | 0.0 a |
| TRIA | 54.8 ab | 33.3 abc | 11.5 a | 15.0 ab |
| TRIA † | 79.0 cd | 65.8 e | 53.3 b | 32.5 bc |
| WOL | 62.5 abc | 36.8 bcd | 8.3 a | 3.3 a |
| WOL † | 85.8 d | 67.5 e | 59.3 b | 42.5 c |
| Shoot Dry Biomass (mg/Plant) | |||
|---|---|---|---|
| 0 mM NaCl L−1 | 40 mM NaCl L−1 | 80 mM NaCl L−1 | |
| CONTROL | 40.8 c | 29.0 bc | 33.2 c |
| ACTIV | 39.1 bc | 42.1 e | 22.0 ab |
| FA | 42.4 c | 30.0 bcd | 26.4 bc |
| HA | 42.0 c | 37.2 cde | 31.8 bc |
| CHITO | 36.3 bc | 35.9 cde | 31.5 bc |
| PTB180 | 38.6 bc | 35.0 cde | 28.7 bc |
| PTB185 | 42.6 c | 36.6 cde | 29.2 bc |
| ROOT | 34.0 bc | 39.5 de | 30.7 bc |
| TRIA | 44.0 c | 35.8 cde | 28.7 bc |
| TRIA † | 8.9 a | 22.9 ab | 12.1 a |
| WOL | 44.1 c | 37.4 cde | 28.3 bc |
| WOL † | 28.8 b | 18.4 a | 21.8 ab |
| Shoot Dry Biomass (mg/Plant) | |||
|---|---|---|---|
| 0 mM NaCl L−1 | 40 mM NaCl L−1 | 80 mM NaCl L−1 | |
| CONTROL | 38.4 ab | 44.3 a | 43.2 b |
| ACTIV | 34.1 a | 42.2 a | 32.2 ab |
| FA | 49.6 b | 44.5 a | 24.1 a |
| HA | 34.9 a | 40.8 a | 20.5 a |
| CHITO | 34.0 a | 39.9 a | 15.7 a |
| PTB180 | 39.5 ab | 40.8 a | 44.0 b |
| PTB185 | 32.9 a | 39.3 a | 42.0 b |
| ROOT | 40.0 ab | 31.3 a | 25.6 a |
| TRIA | 40.6 ab | 36.8 a | 25.1 a |
| TRIA † | 33.0 a | 35.1 a | 30.0 ab |
| WOL | 36.0 ab | 40.7 a | 30.0 ab |
| WOL † | 31.6 a | 35.7 a | 31.2 ab |
| 0 mM NaCl L−1 | 40 mM NaCl L−1 | 80 mM NaCl L−1 | ||||
|---|---|---|---|---|---|---|
| Fv/Fm | PI | Fv/Fm | PI | Fv/Fm | PI | |
| CONTROL | 0.8445 a | 4.51 ab | 0.8392 abc | 3.52 a | 0.8440 a | 5.22 abc |
| ACTIV | 0.8405 a | 3.75 ab | 0.8210 ab | 2.46 a | 0.8287 a | 4.03 abc |
| FA | 0.8430 a | 2.92 a | 0.8652 c | 4.37 a | 0.8344 a | 6.08 c |
| HA | 0.8397 a | 3.56 ab | 0.8275 ab | 2.64 a | 0.8277 a | 3.84 abc |
| CHITO | 0.8432 a | 3.84 ab | 0.8405 bc | 3.48 a | 0.8357 a | 4.78 abc |
| PTB180 | 0.8110 a | 2.97 ab | 0.8297 abc | 3.55 a | 0.8362 a | 4.86 abc |
| PTB185 | 0.8137 a | 2.48 a | 0.8142 ab | 2.57 a | 0.8330 a | 4.15 abc |
| ROOT | 0.8432 a | 3.46 ab | 0.8177 ab | 3.16 a | 0.8337 a | 3.32 ab |
| TRIA | 0.8222 a | 2.89 a | 0.8260 ab | 4.50 a | 0.8320 a | 4.09 abc |
| TRIA † | 0.8430 a | 2.77 a | 0.8300 abc | 2.51 a | 0.8150 a | 2.92 a |
| WOL | 0.8455 a | 5.58 b | 0.8320 abc | 3.21 a | 0.8447 a | 5.92 bc |
| WOL † | 0.8539 a | 4.00 ab | 0.7952 a | 1.74 a | 0.8067 a | 4.12 abc |
| 0 mM NaCl L−1 | 40 mM NaCl L−1 | 80 mM NaCl L−1 | ||||
|---|---|---|---|---|---|---|
| Fv/Fm | PI | Fv/Fm | PI | Fv/Fm | PI | |
| CONTROL | 0.8247 ab | 3.57 a | 0.8402 a | 4.03 a | 0.8396 ab | 4.74 a |
| ACTIV | 0.8200 ab | 1.72 a | 0.8352 a | 4.14 a | 0.8317 ab | 3.50 a |
| FA | 0.8370 b | 2.90 a | 0.8390 a | 3.24 a | 0.8250 ab | 3.73 a |
| HA | 0.8235 ab | 1.87 a | 0.8295 a | 3.56 a | 0.8410 b | 4.27 a |
| CHITO | 0.8337 b | 3.10 a | 0.8352 a | 3.44 a | 0.8372 ab | 4.80 a |
| PTB180 | 0.7975 a | 1.56 a | 0.8137 a | 3.50 a | 0.8335 ab | 3.84 a |
| PTB185 | 0.8137 a | 2.31 a | 0.8365 a | 3.17 a | 0.8357 ab | 3.94 a |
| ROOT | 0.8242 ab | 2.76 a | 0.8292 a | 3.87 a | 0.8395 ab | 4.30 a |
| TRIA | 0.8342 b | 2.78 a | 0.8340 a | 2.95 a | 0.8375 ab | 4.65 a |
| TRIA † | 0.8190 ab | 2.78 a | 0.8222 a | 3.24 a | 0.7985 a | 4.18 a |
| WOL | 0.8245 ab | 2.35 a | 0.8312 a | 3.43 a | 0.8438 b | 3.81 a |
| WOL † | 0.8279 ab | 3.00 a | 0.8140 a | 3.25 a | 0.8377 ab | 5.08 a |
| Shoot Dry Biomass (mg/Plant) | ||||
|---|---|---|---|---|
| −0.5 kPa | −2 kPa | −4 kPa | −6 kPa | |
| CONTROL | 6934 abc | 4858 a | 4931 a | 5453 a |
| ACTIV | 6284 ab | 6310 b | 4399 a | 5220 a |
| FA | 6163 ab | 6343 b | 4303 a | 5584 a |
| HA | 5677 a | 6607 b | 4840 a | 5131 a |
| CHITO | 7401 bc | 6026 ab | 4065 a | 5390 a |
| PTB180 | 6221 ab | 6709 b | 4598 a | 5193 a |
| PTB185 | 7780 c | 7064 b | 4397 a | 5539 a |
| ROOT | 7224 bc | 6546 b | 4170 a | 5519 a |
| TRIA | 6310 abc | 5959 ab | 4655 a | 5504 a |
| WOL | 6559 abc | 5628 ab | 4673 a | 5477 a |
| Shoot Dry Biomass (mg/Plant) | ||||
|---|---|---|---|---|
| −0.5 kPa | −2 kPa | −4 kPa | −6 kPa | |
| CONTROL | 8231 a | 7552 a | 4919 a | 2755 a |
| ACTIV | 8682 a | 7130 a | 5102 a | 3148 a |
| FA | 9272 a | 7650 a | 5291 a | 2745 a |
| HA | 8149 a | 7515 a | 4942 a | 4550 b |
| CHITO | 8568 a | 7080 a | 4948 a | 2802 a |
| PTB180 | 8296 a | 7276 a | 4735 a | 2762 a |
| PTB185 | 9149 a | 7778 a | 5079 a | 3014 a |
| ROOT | 8807 a | 7529 a | 5151 a | 2214 a |
| TRIA | 9265 a | 7616 a | 5118 a | 1929 a |
| WOL | 8524 a | 7737 a | 5026 a | 2626 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clément, J.; Delisle-Houde, M.; Nguyen, T.T.A.; Dorais, M.; Tweddell, R.J. Effect of Biostimulants on Leafy Vegetables (Baby Leaf Lettuce and Batavia Lettuce) Exposed to Abiotic or Biotic Stress under Two Different Growing Systems. Agronomy 2023, 13, 879. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13030879
Clément J, Delisle-Houde M, Nguyen TTA, Dorais M, Tweddell RJ. Effect of Biostimulants on Leafy Vegetables (Baby Leaf Lettuce and Batavia Lettuce) Exposed to Abiotic or Biotic Stress under Two Different Growing Systems. Agronomy. 2023; 13(3):879. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13030879
Chicago/Turabian StyleClément, Justine, Maxime Delisle-Houde, Thi Thuy An Nguyen, Martine Dorais, and Russell J. Tweddell. 2023. "Effect of Biostimulants on Leafy Vegetables (Baby Leaf Lettuce and Batavia Lettuce) Exposed to Abiotic or Biotic Stress under Two Different Growing Systems" Agronomy 13, no. 3: 879. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13030879