
Effect of Branch Bending on the Canopy Characteristics and Growth of Peach (Prunus persica (L.) Batsch)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design
2.3. Determination of Flower Bud Quality
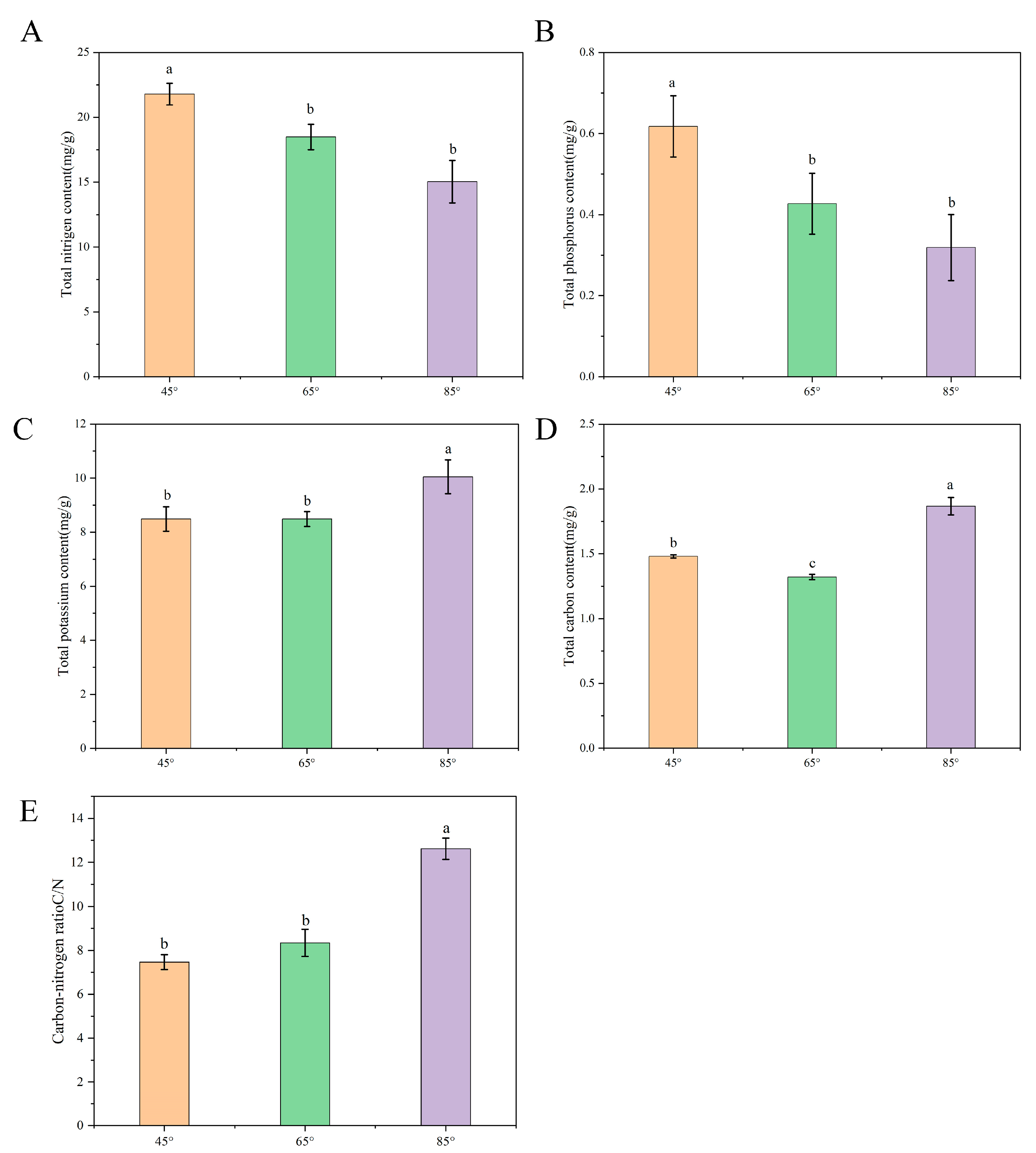
2.4. Determination of the Leaf Photosynthetic Rate and Nutrients
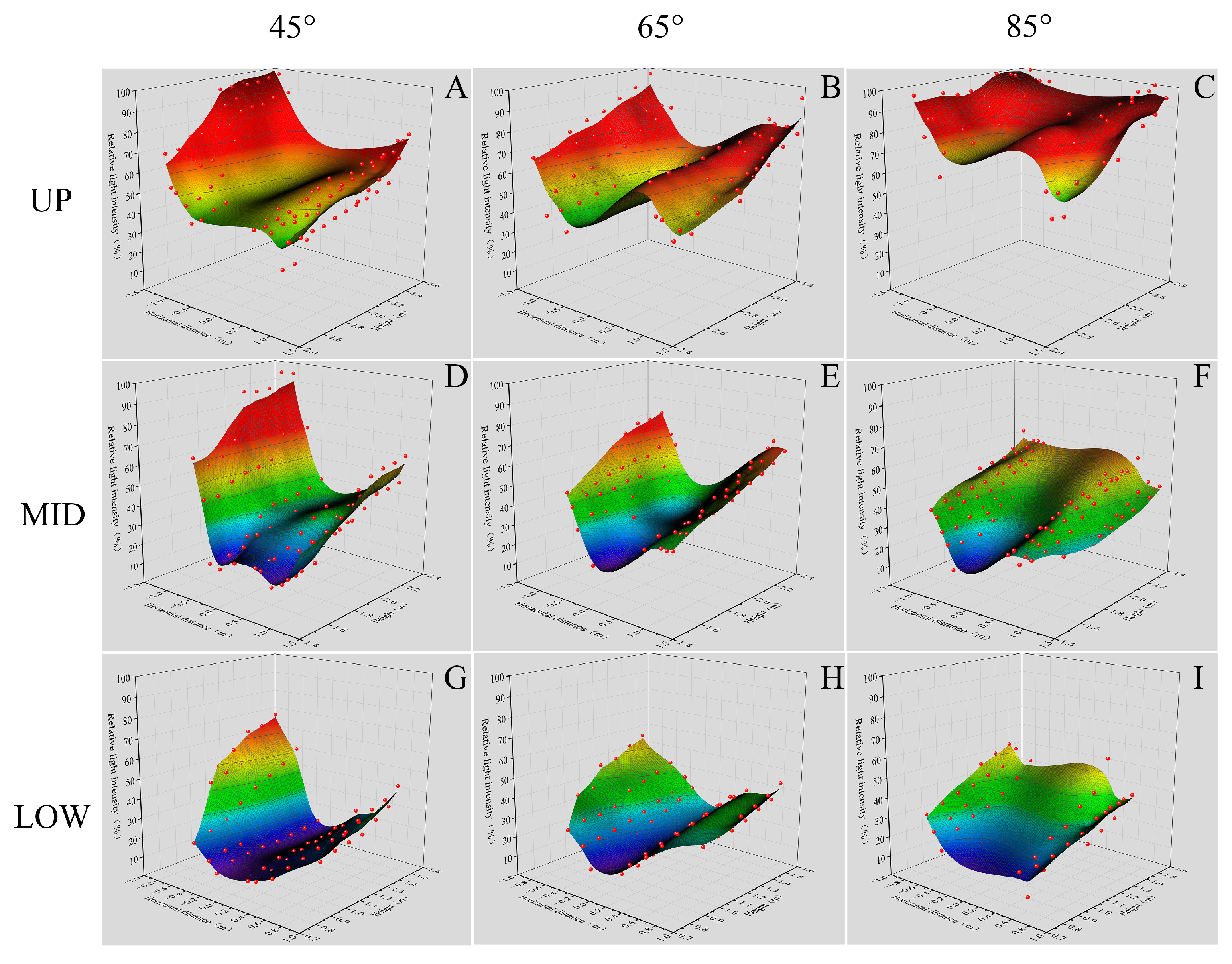
2.5. Measurement of the Canopy Light Intensity
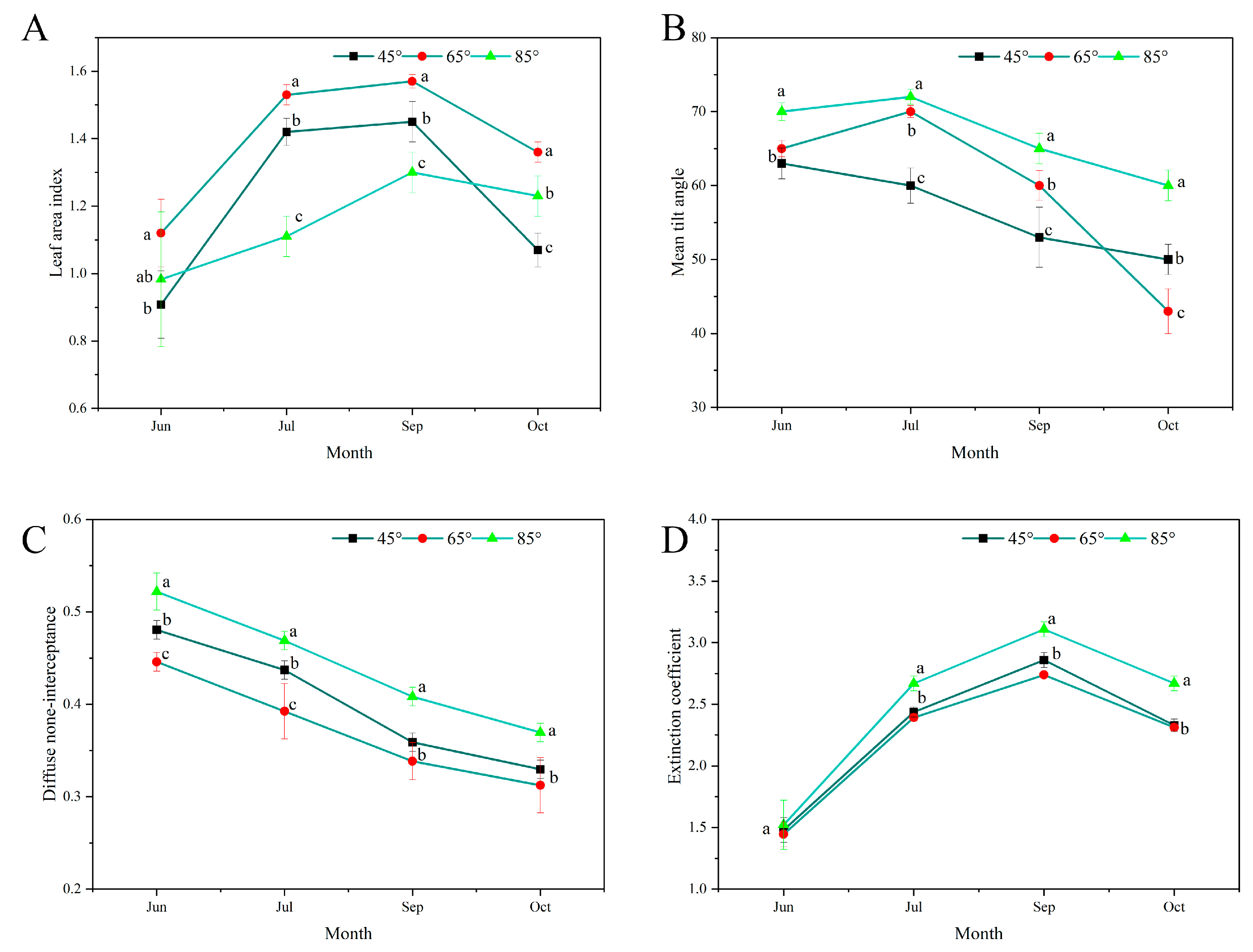
2.6. Measurement of Canopy Characteristics
2.7. Stomatal Aperture Assay
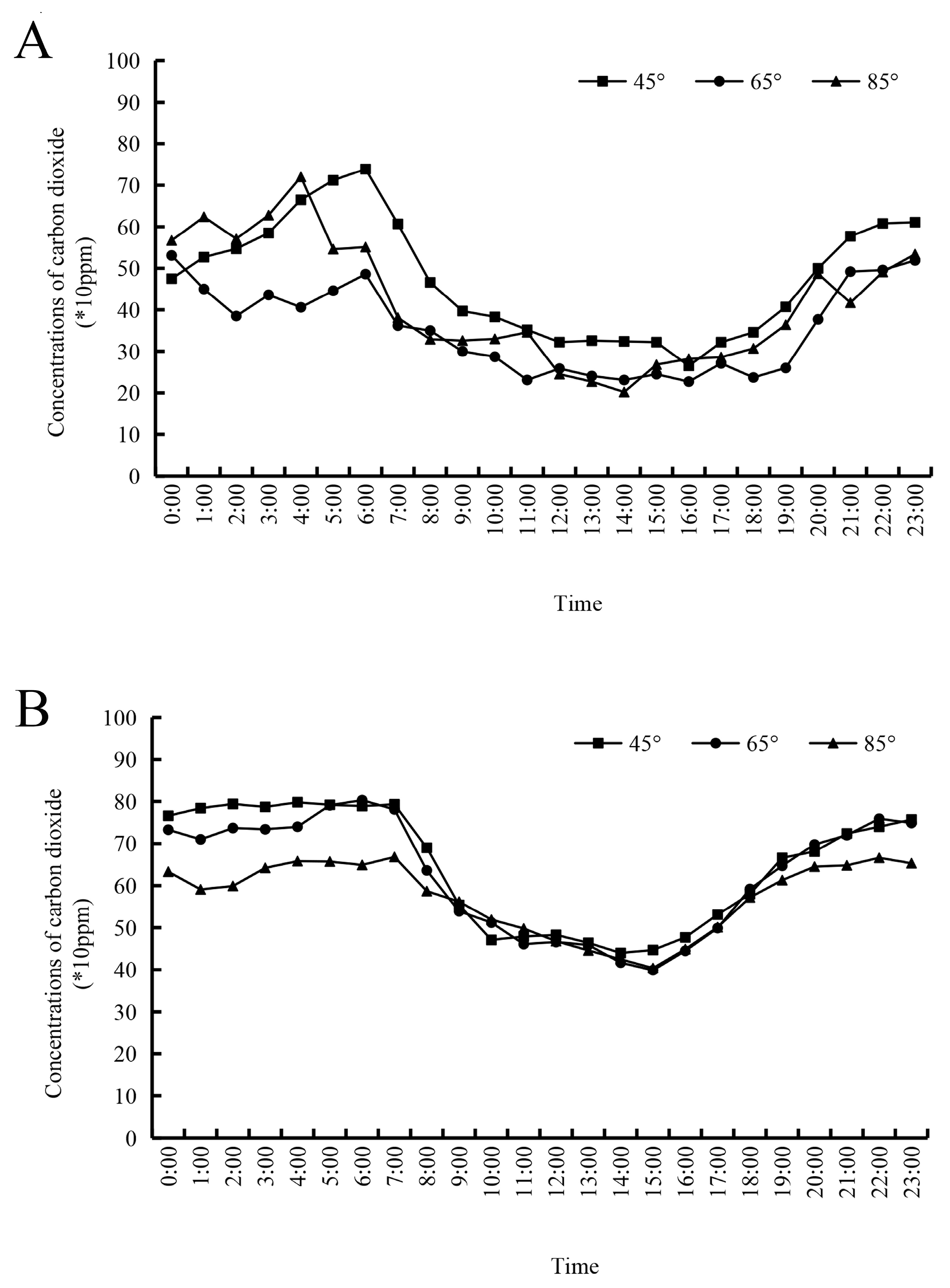
2.8. The Daily Change in the Carbon Dioxide Concentration in the Canopy during Different Seasons
2.9. Determination of the Growth of New Shoots and Main Branches
2.10. Determination of Fruit Quality
2.11. Statistical Analysis
3. Results
3.1. The Quality Difference in Peach Buds with Different Main Branch Opening Angles
3.2. Differences in the Canopy Characteristics among the Different Main Branch Opening Angles
3.3. Daily Variation Pattern of Carbon Dioxide with Different Main Branch Opening Angles
3.4. Effect of Different Main Branch Opening Angles on the Canopy Growth of Peach Trees
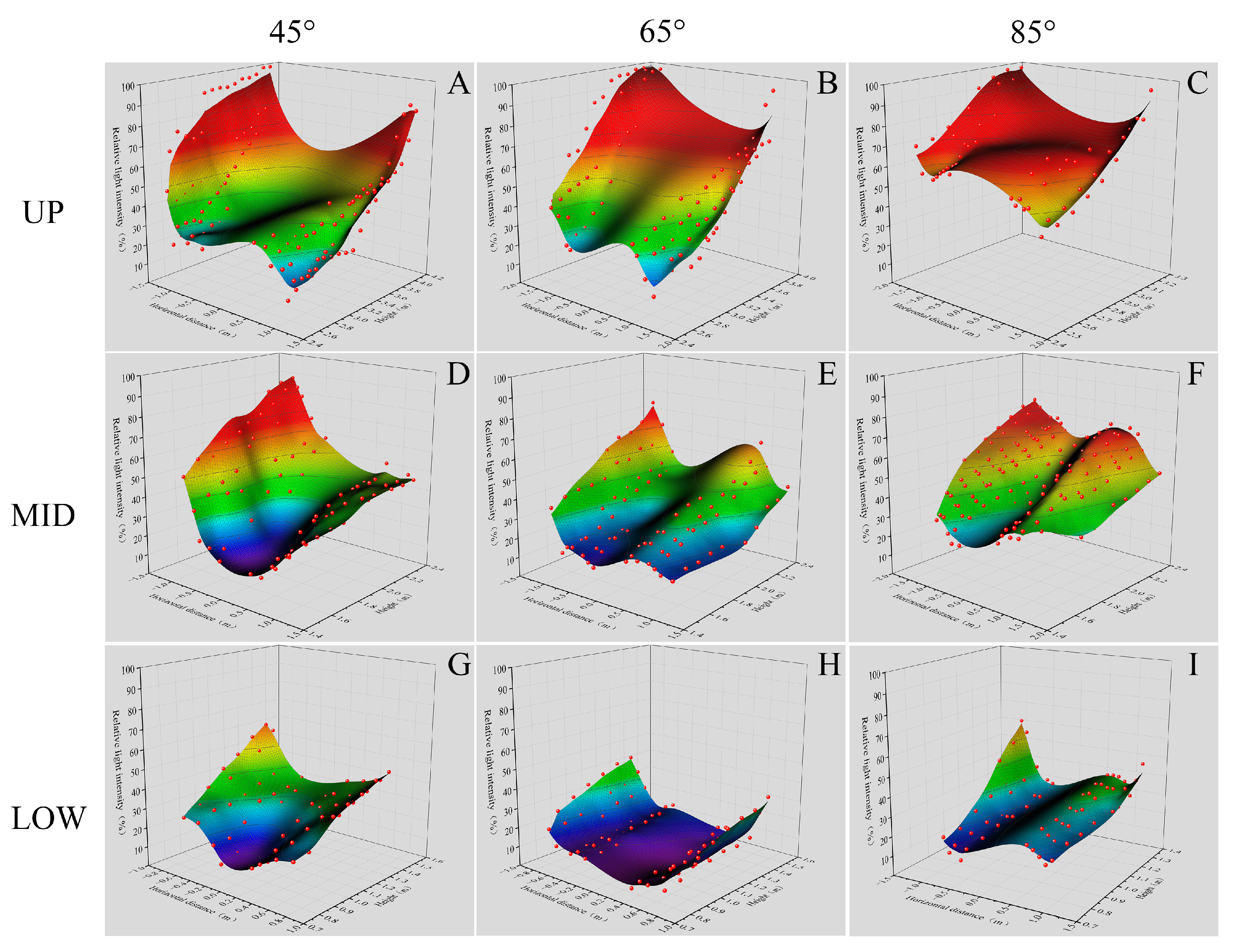
3.5. Differences in Light Distribution in the Canopy
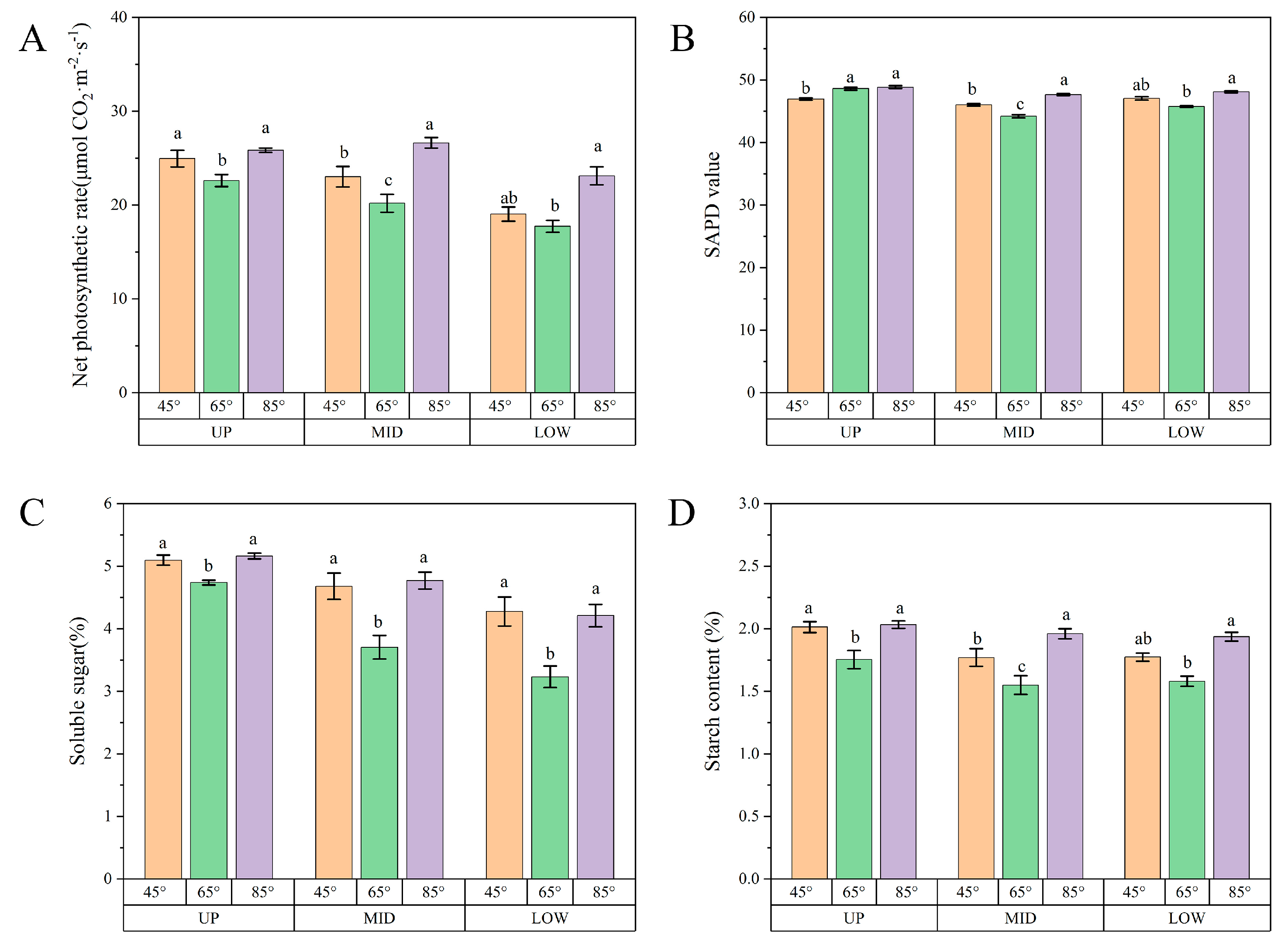
3.6. The Photosynthetic Rate and Photosynthates of Leaves Differed among Different Parts
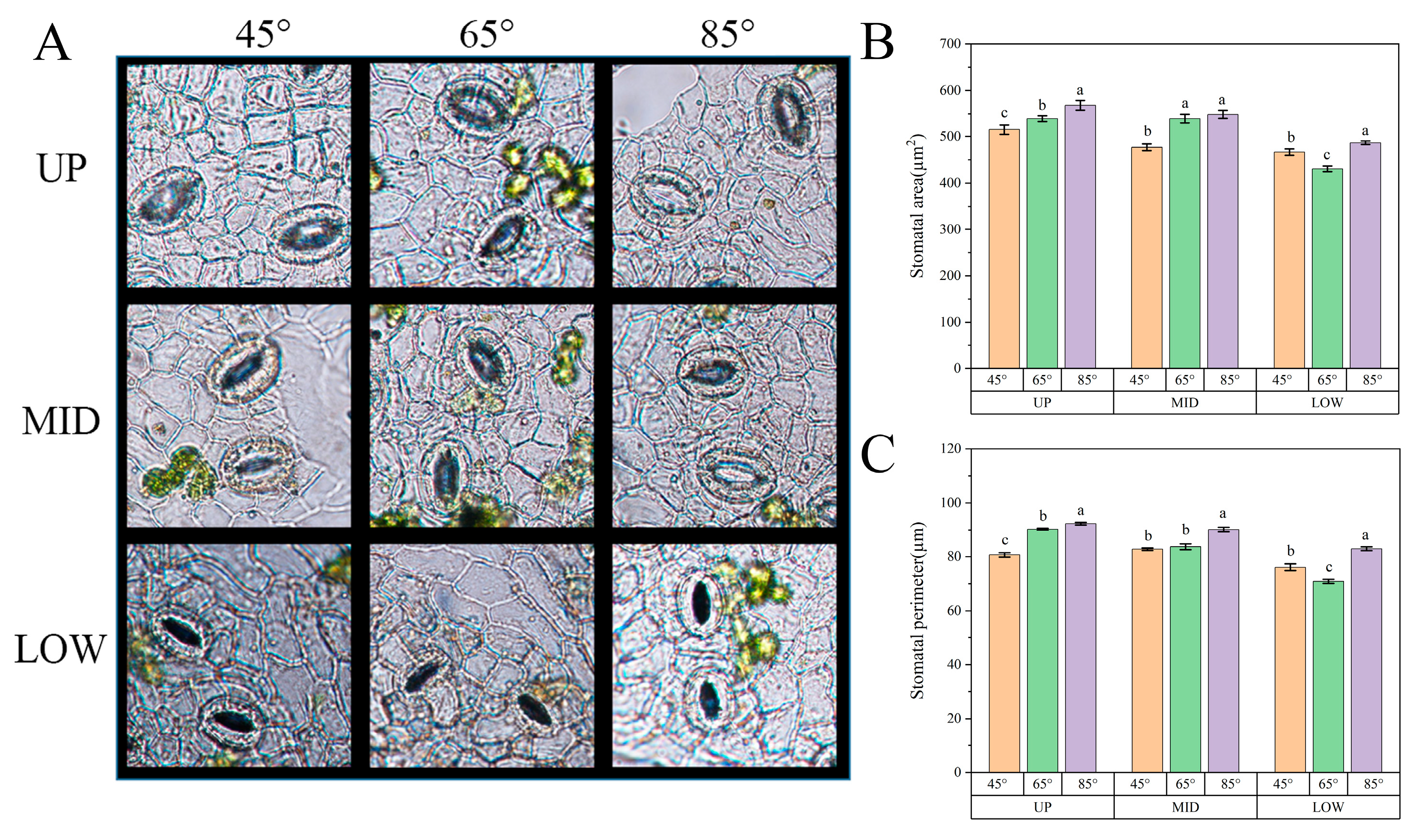
3.7. Leaf Stomatal Morphology Was Different in Different Parts of the Canopy
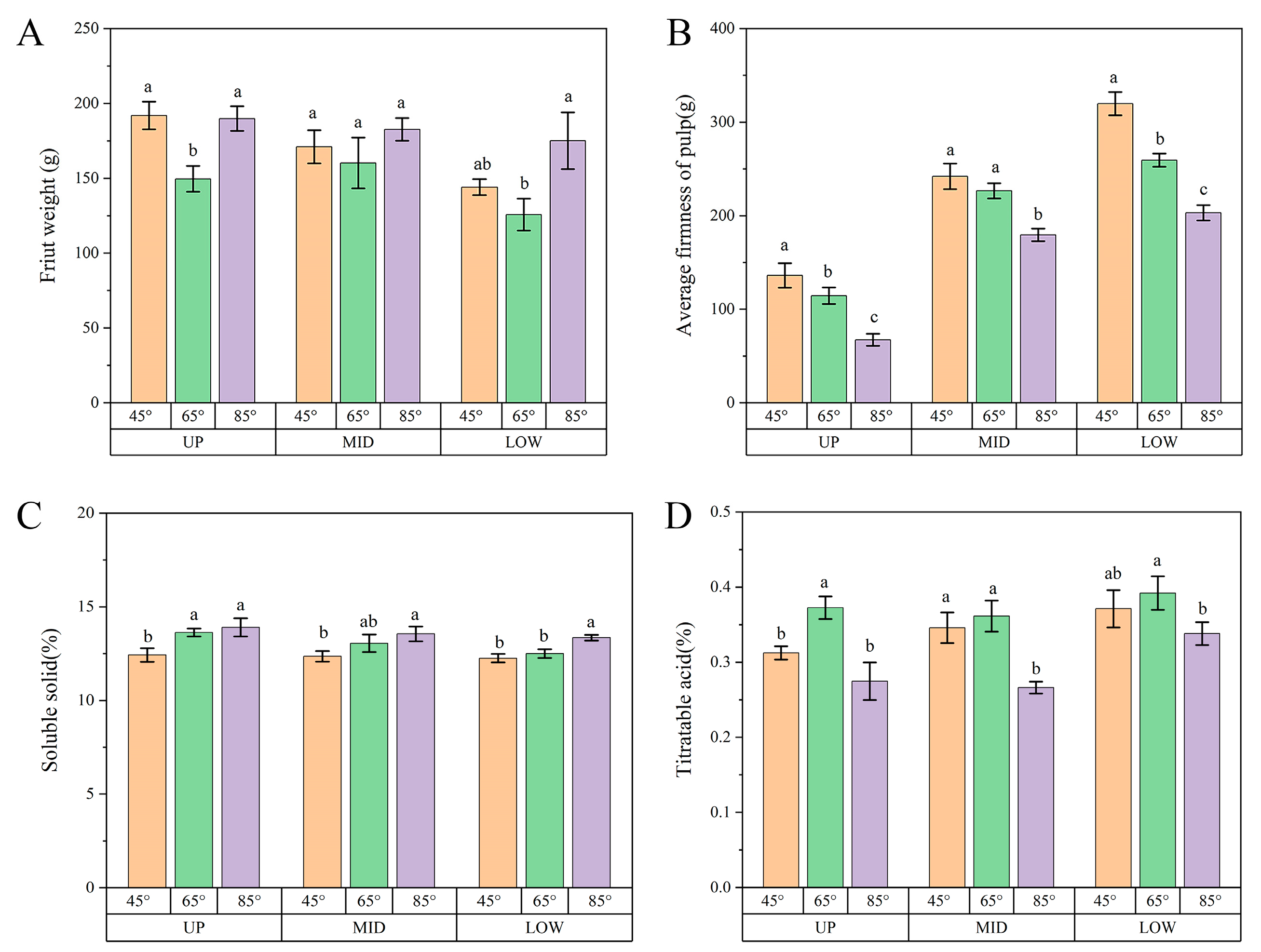
3.8. Differences in Fruit Quality in Different Fruiting Parts
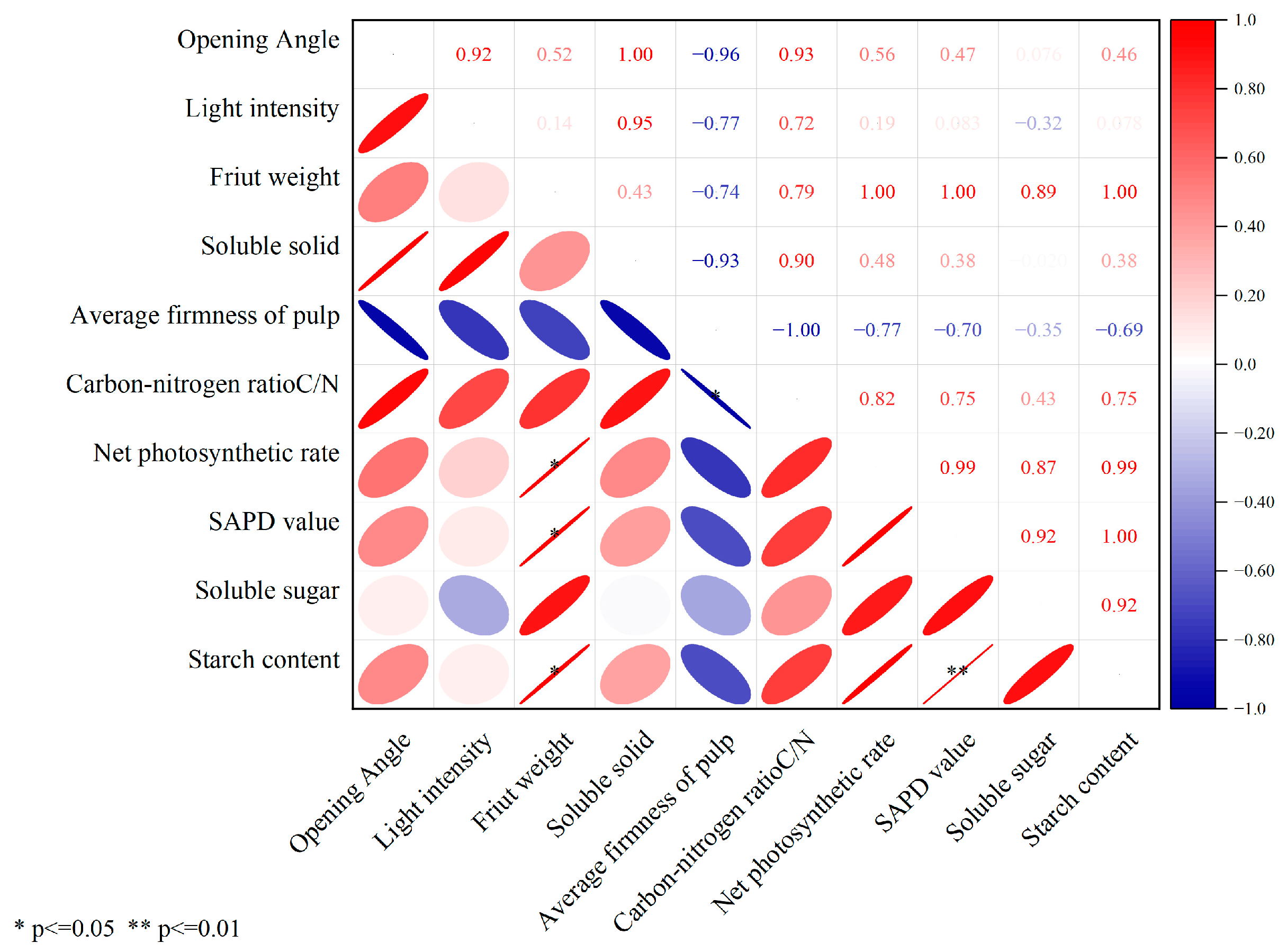
3.9. Correlation Analysis of Canopy Characteristic Indices and Fruit Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bartolini, S.; Leccese, A.; Andreini, L. Influence of canopy fruit location on morphological, histochemical and biochemical changes in two oil olive cultivars. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2014, 148, 1221–1230. [Google Scholar] [CrossRef]
- Farina, V.; Bianco, R.L.; Inglese, P. Vertical Distribution of Crop Load and Fruit Quality within Vase-and Y-shaped Canopies of ‘Elegant Lady’ Peach. HortScience 2005, 40, 587–591. [Google Scholar] [CrossRef] [Green Version]
- Anthony, B.M.; Minas, I.S. Optimizing peach tree canopy architecture for efficient light use, increased productivity and improved fruit quality. Agronomy 2021, 11, 1961. [Google Scholar] [CrossRef]
- Manganaris, G.A.; Minas, I.; Cirilli, M.; Torres, R.; Bassi, D.; Costa, G. Peach for the future: A specialty crop revisited. Sci. Hortic. 2022, 305, 111390. [Google Scholar] [CrossRef]
- Minas, I.S.; Tanou, G.; Molassiotis, A. Environmental and orchard bases of peach fruit quality. Sci. Hortic. 2018, 235, 307–322. [Google Scholar] [CrossRef]
- De Macedo, T.A.; da Silva, P.S.; Sander, G.F.; Welter, J.F.; Rufato, L.; de Rossi, A. Productivity and quality of ‘Fuji Suprema’ apple fruit in different rootstocks and growing conditions. Sci. Hortic. 2019, 256, 5. [Google Scholar] [CrossRef]
- Lauzike, K.; Uselis, N.; Samuoliene, G. The Influence of Rootstock and High-Density Planting on Apple cv. Auksis Fruit Quality. Plants 2021, 10, 11. [Google Scholar] [CrossRef]
- Legua, P.; Martinez-Cuenca, M.R.; Bellver, R.; Forner-Giner, M.A. Rootstock’s and scion’s impact on lemon quality in southeast Spain. Int. Agrophys. 2018, 32, 325–333. [Google Scholar] [CrossRef]
- Lu, X.; Gao, Y.; Wang, K.; Sun, S.M.; Liu, Z.; Yan, P.; Feng, J.R.; Li, Q.S.; Li, L.W.; Wang, D.J. Dwarf Interstocks Improve Aroma Quality of ‘Huahong’ Apple (Malus × domestica). Agriculture 2022, 12, 13. [Google Scholar] [CrossRef]
- Musacchi, S.; Gagliardi, F.; Serra, S. New training systems for high-density planting of sweet cherry. HortScience 2015, 50, 59–67. [Google Scholar] [CrossRef]
- Reig, G.; Lordan, J.; Fazio, G.; Grusak, M.A.; Hoying, S.; Cheng, L.L.; Francescatto, P.; Robinson, T. Horticultural performance and elemental nutrient concentrations on ‘Fuji’ grafted on apple rootstocks under New York State climatic conditions. Sci. Hortic. 2018, 227, 22–37. [Google Scholar] [CrossRef]
- Stachowiak, A.; Bosiacki, M.; Swierczynski, S.; Kolasinski, M. Influence of rootstocks on different sweet cherry cultivars and accumulation of heavy metals in leaves and fruit. Hortic. Sci. 2015, 42, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.-K.; Choi, H.-S. Light penetration, growth, and fruit productivity in ‘Fuji’ apple trees trained to four growing systems. Sci. Hortic. 2010, 125, 672–678. [Google Scholar] [CrossRef]
- Jung, S.-K.; Ryu, H.-U.; Gu, M.; Choi, H.-S.; Ryu, J.-H.; Cho, Y.-K. Growth and fruit production in young ‘Fuji’ apple trees as affected by time of downward bending of branches. J. Hortic. Sci. Biotechnol. 2012, 87, 519–523. [Google Scholar] [CrossRef]
- Jung, J. Effect of Pruning Methods on the Tree Growth, Yield and Quality of Fruits in Dwarf Apple Trees (‘Fuji’/M. 26/Malus prunifolia seedlings). Ph.D. Thesis, Kyung Hee University, Yongin, Korea, 1993. [Google Scholar]
- Zhang, M.; Han, M.; Ma, F.; Shu, H. Effect of bending on the dynamic changes of endogenous hormones in shoot terminals of ‘Fuji’ and ‘Gala’ apple trees. Acta Physiol. Plant. 2015, 37, 76. [Google Scholar] [CrossRef]
- Zhang, M.; Ma, F.; Shu, H.; Han, M. Branch bending affected floral bud development and nutrient accumulation in shoot terminals of ‘Fuji’ and ‘Gala’ apples. Acta Physiol. Plant. 2017, 39, 156. [Google Scholar] [CrossRef]
- Robinson, T.L.; Lakso, A.N.; Ren, Z.-b. Modifying apple tree canopies for improved production efficiency. HortScience 1991, 26, 1005–1012. [Google Scholar] [CrossRef] [Green Version]
- Lakso, A.N.; Robinson, T.L. Sunlight, yield, and productivity of apples. N. Y. Fruit Q. 2014, 22, 7. [Google Scholar]
- Zhang, Y.-Q.; Wen, Y.; Bai, Q.; Ma, Z.; Ye, H.-L.; Su, S.-C. Spatio-temporal effects of canopy microclimate on fruit yield and quality of Sapindus mukorossi Gaertn. Sci. Hortic. 2019, 251, 136–149. [Google Scholar] [CrossRef]
- Yue, W.; Shu-chai, S.; Ma, L.-y.; Shao-yan, Y.; Yu-wei, W.; Wang, X.-n. Effects of canopy microclimate on fruit yield and quality of Camellia oleifera. Sci. Hortic. 2018, 235, 132–141. [Google Scholar] [CrossRef]
- Lakso, A.; Corelli Grappadelli, L. Implications of pruning and training practices to carbon partitioning and fruit development in apple. In International Symposium on Training and Pruning of Fruit Trees; ISHS: Sheprdstown, WV, USA, 1991; Volume 322, pp. 231–240. [Google Scholar]
- Buler, Z.; Mika, A. The influence of canopy architecture on light interception and distribution in ‘Sampion’ apple trees. J. Fruit Ornam. Plant Res. 2009, 17, 45–52. [Google Scholar]
- Han, H.-H.; Coutand, C.; Cochard, H.; Trottier, C.; Lauri, P.-E. Effects of shoot bending on lateral fate and hydraulics: Invariant and changing traits across five apple genotypes. J. Exp. Bot. 2007, 58, 3537–3547. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.; Hoying, S.; Barritt, H. Descriptions of orchard planting systems. Compact. Fruit Tree 2003, 36, 50–64. [Google Scholar]
- Costes, E.; Lauri, P.-E.; Regnard, J.-L. Analyzing fruit tree architecture: Implications for tree management and fruit production. Hortic. Rev. 2006, 32, 1–61. [Google Scholar]
- Lauri, P.; Laurens, F. Architectural types in apple (Malus × domestica Borkh.). In Crops Growth Quality Biotechnology; World Food Limited: Helsinki, Finland, 2005; pp. 1300–1314. [Google Scholar]
- Xing, L.; Zhang, D.; Zhao, C.; Li, Y.; Ma, J.; An, N.; Han, M. Shoot bending promotes flower bud formation by mi RNA-mediated regulation in apple (Malus domestica Borkh.). Plant Biotechnol. J. 2016, 14, 749–770. [Google Scholar] [CrossRef] [Green Version]
- Ball, R.A.; Purcell, L.C.; Carey, S.K. Evaluation of solar radiation prediction models in North America. Agron. J. 2004, 96, 391–397. [Google Scholar] [CrossRef]
- De Bruin, J.L.; Pedersen, P. Effect of row spacing and seeding rate on soybean yield. Agron. J. 2008, 100, 704–710. [Google Scholar] [CrossRef]
- Giunta, F.; Motzo, R.; Pruneddu, G. Has long-term selection for yield in durum wheat also induced changes in leaf and canopy traits? Field Crops Res. 2008, 106, 68–76. [Google Scholar] [CrossRef]
- Feng, G.; Luo, H.; Zhang, Y.; Gou, L.; Yao, Y.; Lin, Y.; Zhang, W. Relationship between plant canopy characteristics and photosynthetic productivity in diverse cultivars of cotton (Gossypium hirsutum L.). Crop J. 2016, 4, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Liu, Y.; Guo, Y.; Peng, C.; Tan, W.; Zhang, M.; Li, Z.; Zhou, Y.; Duan, L. A novel plant growth regulator improves the grain yield of high-density maize crops by reducing stalk lodging and promoting a compact plant type. Field Crops Res. 2021, 260, 107982. [Google Scholar] [CrossRef]
- Tabarzad, A.; Ghaemi, A.A.; Zand-Parsa, S. Extinction coefficients and radiation use efficiency of barley under different irrigation regimes and sowing dates. Agric. Water Manag. 2016, 178, 126–136. [Google Scholar] [CrossRef]
- Sharma, Y.; Singh, H.; Singh, S. Effect of light interception and penetration at different levels of fruit tree canopy on quality of peach. Curr. Sci. 2018, 115, 1562–1566. [Google Scholar] [CrossRef]
- Lu, Y.; Si, Y.; Zhang, L.; Sun, Y.; Su, S. Effects of Canopy Position and Microclimate on Fruit Development and Quality of Camellia oleifera. Agronomy 2022, 12, 2158. [Google Scholar] [CrossRef]
- Xie, T.-T.; Su, P.-X.; An, L.-Z.; Shan, L.-S.; Zhou, Z.-J.; Chai, Z.-P. Physiological characteristics of high yield under cluster planting: Photosynthesis and canopy microclimate of cotton. Plant Prod. Sci. 2016, 19, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Kostopoulou, P.; Karatassiou, M. Lotus corniculatus L. response to carbon dioxide concentration and radiation level variations. Photosynthetica 2017, 55, 522–531. [Google Scholar] [CrossRef]
- Hampson, C.R.; Quamme, H.A.; Brownlee, R.T. Canopy growth, yield, and fruit quality of ‘Royal Gala’ apple trees grown for eight years in five tree training systems. HortScience 2002, 37, 627–631. [Google Scholar] [CrossRef] [Green Version]
- Zhaoyuan, W.; Qihang, T.; Yong, G.; Chang, R.; Guojian, L.; Hu, C.; Yonghong, L. The Distribution of Relative Light Intensity in a Peach Tree Canopy Affects Fruit Yield and Texture. Agric. Biotechnol. 2021, 10, 2164–4993. [Google Scholar]
- Lewallen, K.S.; Marini, R.P. Relationship between flesh firmness and ground color in peach as influenced by light and canopy position. J. Am. Soc. Hortic. Sci. 2003, 128, 163–170. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Angle | Height (m) | East–West Crown Diameter (m) | North–South Crown Diameter (m) |
|---|---|---|---|
| 45° | 4.08 ± 0.07 a | 2.56 ± 0.15 b | 2.20 ± 0.10 a |
| 65° | 3.82 ± 0.06 b | 3.10 ± 0.26 ab | 2.16 ± 0.25 a |
| 85° | 3.01 ± 0.06 c | 3.36 ± 0.50 a | 2.46 ± 0.06 a |
| Angle | Short Fruiting Branch | Medium Fruiting Branch | Long Fruiting Branch | Epicormic Branch | New Shoot Length (cm) | Main Branch Length (cm) | Main Trunk Length (cm) |
|---|---|---|---|---|---|---|---|
| 45° | 79.3 ± 11.0 a | 59.3 ± 1.8 a | 60.3 ± 3.9 a | 5.0 ± 0.6 b | 37.8 ± 2.27 a | 4.07 ± 0.3 a | 21.67 ± 1.2 a |
| 65° | 58.7 ± 5.2 ab | 54.7 ± 5.4 ab | 49.3 ± 1.5 b | 12.0 ± 2.1 a | 36.1 ± 1.89 ab | 3.83 ± 0.18 a | 16.67 ± 0.35 b |
| 85° | 43.0 ± 6.1 b | 44.0 ± 2.6 b | 38.0 ± 2.6 c | 8.3 ± 0.9 ab | 29.56 ± 2.5 b | 2.73 ± 0.23 b | 9.33 ± 0.17 c |
| Angle | Location | <30% | 30–60% | >60% |
|---|---|---|---|---|
| Upper | 15.2 | 49.1 | 35.7 | |
| 45° | Middle | 33.8 | 52.4 | 13.8 |
| Lower | 56.5 | 35.2 | 8.3 | |
| Upper | 22.2 | 40.9 | 36.9 | |
| 65° | Middle | 39.5 | 52.7 | 7.8 |
| Lower | 59.8 | 36.9 | 3.3 | |
| Upper | 3.4 | 16.9 | 79.7 | |
| 85° | Middle | 13.2 | 76.9 | 9.9 |
| Lower | 39.5 | 58.1 | 2.4 |
| Angle | Location | <30% | 30–60% | >60% |
|---|---|---|---|---|
| Upper | 33.9 | 47.2 | 18.9 | |
| 45° | Middle | 39.5 | 47.8 | 12.7 |
| Lower | 56.1 | 37.8 | 6.1 | |
| Upper | 25.4 | 40.9 | 33.7 | |
| 65° | Middle | 30.9 | 57.8 | 11.3 |
| Lower | 74.5 | 25.2 | 0.3 | |
| Upper | 5.9 | 30.8 | 63.3 | |
| 85° | Middle | 13.9 | 58.6 | 27.5 |
| Lower | 54.5 | 44 | 1.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Zheng, F.; Geng, W.; Du, H.; Xiao, Y.; Peng, F. Effect of Branch Bending on the Canopy Characteristics and Growth of Peach (Prunus persica (L.) Batsch). Agronomy 2023, 13, 1058. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13041058
Zhang B, Zheng F, Geng W, Du H, Xiao Y, Peng F. Effect of Branch Bending on the Canopy Characteristics and Growth of Peach (Prunus persica (L.) Batsch). Agronomy. 2023; 13(4):1058. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13041058
Chicago/Turabian StyleZhang, Binbin, Fengshi Zheng, Wenwen Geng, Hao Du, Yuansong Xiao, and Futian Peng. 2023. "Effect of Branch Bending on the Canopy Characteristics and Growth of Peach (Prunus persica (L.) Batsch)" Agronomy 13, no. 4: 1058. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13041058