
Genome-Wide Identification and Expression Pattern Analysis of the F5H Gene Family in Flax (Linum usitatissimum L.)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of F5H Genes in Linum usitatissimum
2.2. Sequence Analysis and Structural Characterization of LuF5Hs
2.3. Phylogenetic Analysis of the LuF5H Gene Family
2.4. Analysis of cis-Acting Elements in the LuF5H Gene Promoter
2.5. PPI Network of F5H Proteins in Linum usitatissimum
2.6. Analysis of Expression Pattern of LuF5Hs
2.7. Drought, NaCl and Brassinosteroid (BR) Treatments
2.8. RNA Extraction
2.9. DGE Library Preparation and Sequencing
2.10. Real-Time Quantitative PCR Analysis
2.11. Determination of Lignin Content and Correlation Analysis
3. Results
3.1. Identification and Basic Information of F5H Genes in Linum usitatissimum
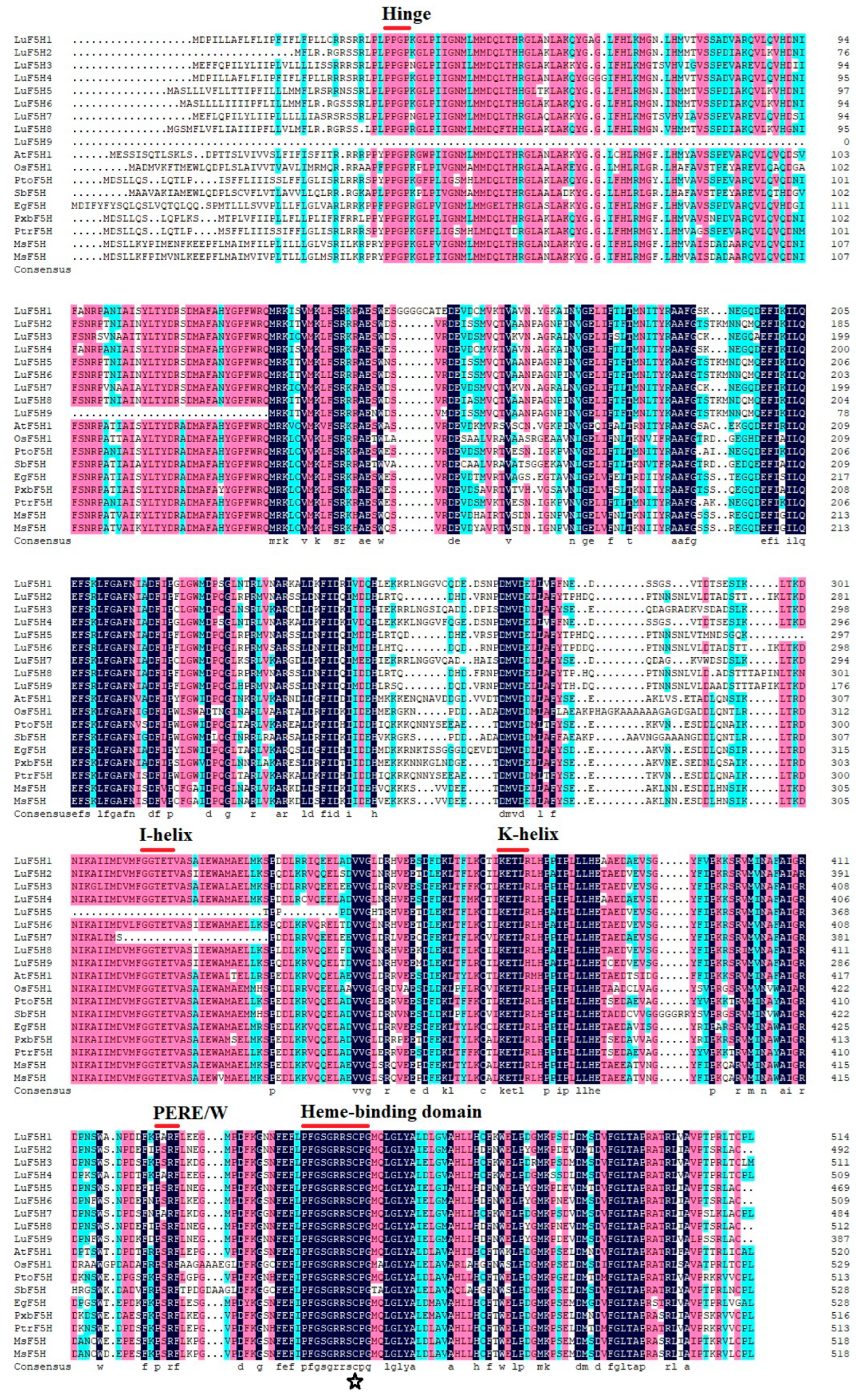
3.2. Multiple Sequence Alignment of LuF5H Proteins
3.3. LuF5H Gene Structures and Conserved Motifs
3.4. Phylogenetic Analysis and Classification of the F5H Gene Family in Linum usitatissimum
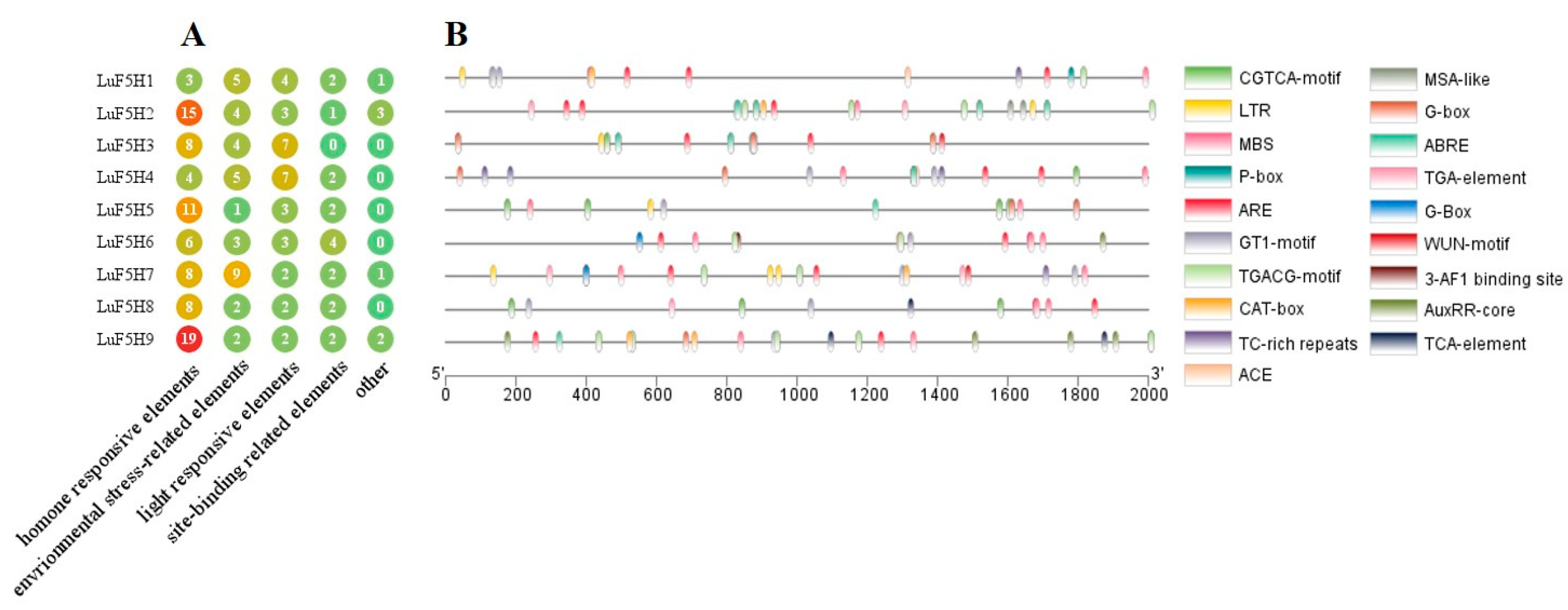
3.5. Cis-Element Analysis in the Promoters of LuF5H Genes
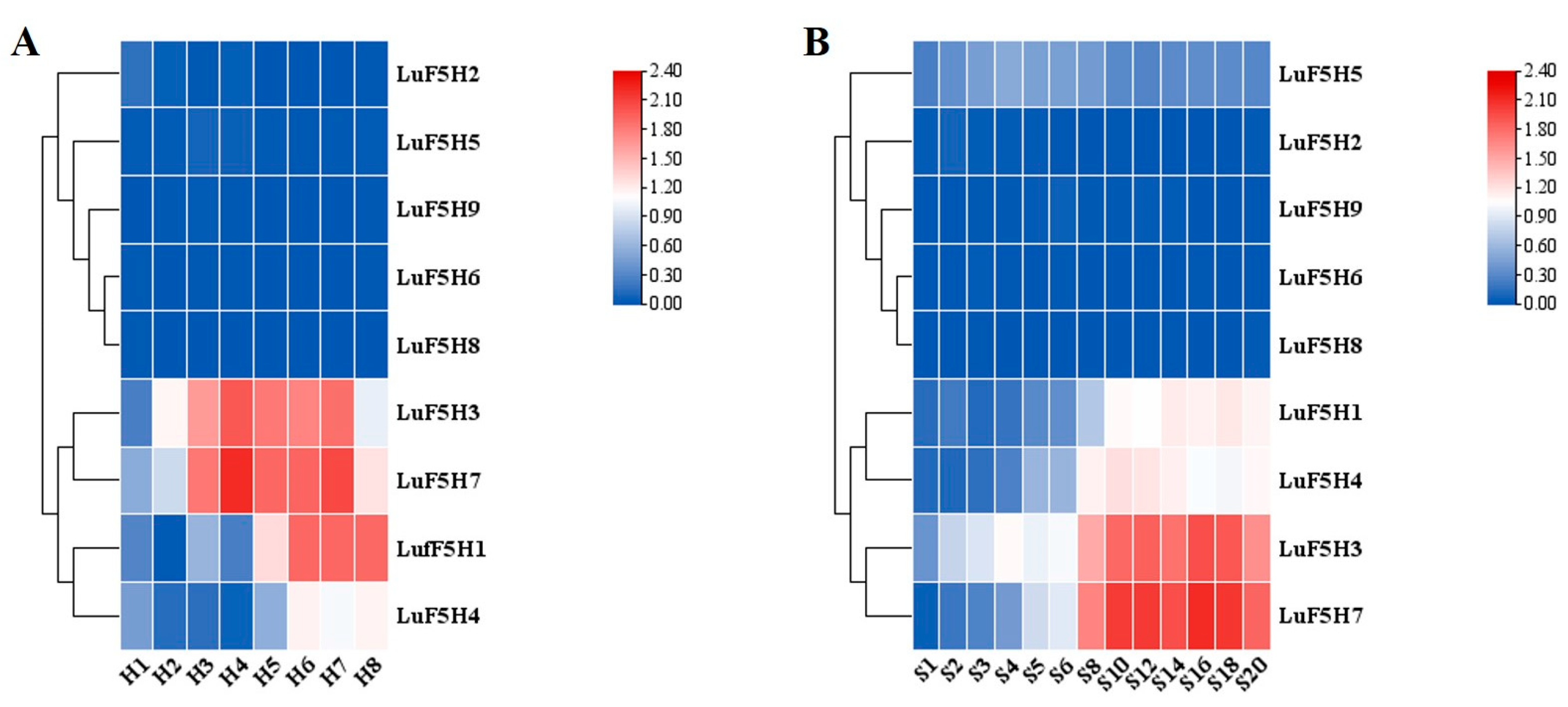
3.6. Analysis of Expression Pattern of LuF5H Genes
3.7. Interaction Analysis between F5H Proteins in Linum usitatissimum
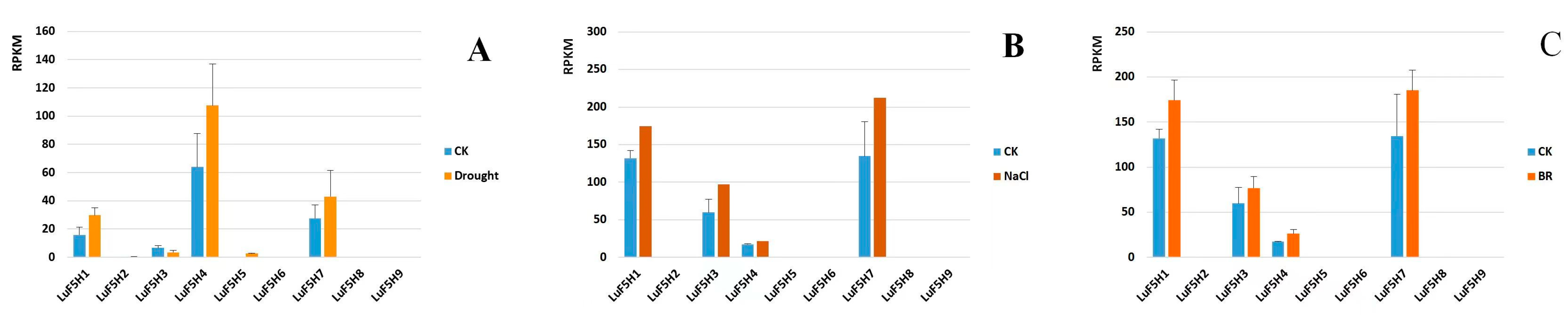
3.8. Expression Profiles of LuF5H Genes in Response to Drought, NaCl and BR Treatments
3.9. Correlation Analysis between Lignin Content and LuF5H Gene Expression Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sa, R.; Yi, L.X.; Siqin, B.; An, M.; Bao, H.Z.; Song, X.L.; Wang, S.Y.; Li, Z.W.; Zhang, Z.; Hazaisi, H.; et al. Chromosome-Level Genome Assembly and Annotation of the Fiber Flax (Linum usitatissimum) Genome. Front. Genet. 2021, 12, 735690. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.M.; Guo, W.D.; Zhao, L.J.; Yu, Y.; Chen, S.; Tao, L.; Cheng, L.L.; Kang, Q.H.; Song, X.X.; Wu, J.Z.; et al. Genome-wide identification and expression analysis of the WRKY transcription factor family in flax (Linum usitatissimum L.). BMC Genom. 2021, 22, 375. [Google Scholar] [CrossRef]
- Yuan, H.M.; Guo, W.D.; Zhao, L.J.; Yu, Y.; Chen, S.; Cheng, L.L.; Kang, Q.H.; Song, X.X.; Wu, J.Z.; Huang, W.G.; et al. Expression Characteristics Analysis of CesA/Csl genes in Flax during the Fast Growing Stage. J. Nat. Fibers 2021, 19, 7864–7877. [Google Scholar] [CrossRef]
- Zimniewska, M. Antioxidant Activity of Fibres Originating from Traditional Varieties of Polish Flax Plants. Fibres Text. 2015, 23, 41–47. [Google Scholar] [CrossRef]
- Zimniewska, M.; Rozanska, W.; Gryszczynska, A.; Romanowska, B.; Kicinska-Jakubowska, A. Antioxidant Potential of Hemp and Flax Fibers Depending on Their Chemical Composition. Molecules 2018, 23, 1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, S.W.; Xu, B.R.; Cheng, L.F.; Feng, X.Y.; Yang, Q.; Zheng, K.; Gao, M.Q.; Liu, Z.Y.; Liu, C.J.; Peng, Y.D. Bacterial strain for bast fiber crops degumming and its bio-degumming technique. Bioprocess Biosyst. Eng. 2021, 44, 2503–2512. [Google Scholar] [CrossRef]
- Kim, J.; Choi, B.; Park, Y.H.; Cho, B.K.; Lim, H.S.; Natarajan, S.; Park, S.U.; Bae, H. Molecular Characterization of Ferulate 5-Hydroxylase Gene from Kenaf (Hibiscus cannabinus L.). Sci. World J. 2013, 2013, 421578. [Google Scholar] [CrossRef] [Green Version]
- Shafrin, F.; Das, S.S.; Sanan-Mishra, N.; Khan, H. Artificial miRNA-mediated down-regulation of two monolignoid biosynthetic genes (C3H and F5H) cause reduction in lignin content in jute. Plant Mol. Biol. 2015, 89, 511–527. [Google Scholar] [CrossRef]
- Takeda, Y.; Koshiba, T.; Tobimatsu, Y.; Suzuki, S.; Murakami, S.; Yamamura, M.; Rahman, M.M.; Takano, T.; Hattori, T.; Sakamoto, M.; et al. Regulation of CONIFERALDEHYDE 5-HYDROXYLASE expression to modulate cell wall lignin structure in rice. Planta 2017, 246, 337–349. [Google Scholar] [CrossRef]
- Tetreault, H.M.; Gries, T.; Palmer, N.A.; Funnell-Harris, D.L.; Satoz, S.; Ge, Z.X.; Sarath, G.; Sattler, S.E. Overexpression of ferulate 5-hydroxylase increases syringyl units in Sorghum bicolor. Plant Mol. Biol. 2020, 103, 269–285. [Google Scholar] [CrossRef]
- Huntley, S.K.; Ellis, D.; Gilbert, M.; Chapple, C.; Mansfield, S.D. Significant increases in pulping efficiency in C4H-F5H-transformed poplars: Improved chemical savings and reduced environmental toxins. J. Agric. Food Chem. 2003, 51, 6178–6183. [Google Scholar] [CrossRef] [PubMed]
- Welker, C.M.; Balasubramanian, V.K.; Petti, C.; Rai, K.M.; DeBolt, S.; Mendu, V. Engineering Plant Biomass Lignin Content and Composition for Biofuels and Bioproducts. Energies 2015, 8, 7654–7676. [Google Scholar] [CrossRef] [Green Version]
- Bewg, W.P.; Poovaiah, C.; Lan, W.; Ralph, J.; Coleman, H.D. RNAi downregulation of three key lignin genes in sugarcane improves glucose release without reduction in sugar production. Biotechnol. Biofuels 2016, 9, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphreys, J.M.; Chapple, C. Rewriting the lignin roadmap. Curr. Opin. Plant Biol. 2002, 5, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Kwon, S.J.; Sung, S.Y.; Kim, W.J.; Kim, D.S.; Ahn, J.W.; Kim, J.B.; Kim, S.H.; Ha, B.K.; Kang, S.Y. Molecular cloning, characterization, and expression analysis of lignin biosynthesis genes from kenaf (Hibiscus cannabinus L.). Genes Genom. 2016, 38, 59–67. [Google Scholar] [CrossRef]
- Tang, Y.H.; Liu, F.; Xing, H.C.; Mao, K.Q.; Chen, G.; Guo, Q.Q.; Chen, J.R. Correlation Analysis of Lignin Accumulation and Expression of Key Genes Involved in Lignin Biosynthesis of Ramie (Boehmeria nivea). Genes 2019, 10, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin Biosynthesis and Structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Ralph, J.; Lundquist, K.; Brunow, G.; Lu, F.; Kim, H.; Schatz, P.F.; Marita, J.M.; Hatfield, R.D.; Ralph, S.A.; Christensen, J.H.; et al. Lignins: Natural polymers from oxidative coupling of 4-hydroxyphenyl-propanoids. Phytochem. Rev. 2004, 3, 29–60. [Google Scholar] [CrossRef]
- Ciesielski, P.N.; Resch, M.G.; Hewetson, B.; Killgore, J.P.; Curtin, A.; Anderson, N.; Chiaramonti, A.N.; Hurley, D.C.; Sanders, A.; Himmel, M.E.; et al. Engineering plant cell walls: Tuning lignin monomer composition for deconstructable biofuel feedstocks or resilient biomaterials. Green Chem. 2014, 16, 2627–2635. [Google Scholar] [CrossRef]
- Stewart, J.J.; Akiyama, T.; Chapple, C.; Ralph, J.; Mansfield, S.D. The Effects on Lignin Structure of Overexpression of Ferulate 5-Hydroxylase in Hybrid Poplar. Plant Physiol. 2009, 150, 621–635. [Google Scholar] [CrossRef] [Green Version]
- Bhinu, V.S.; Li, R.; Huang, J.; Kaminskyj, S.; Sharpe, A.; Hannoufa, A. Perturbation of lignin biosynthesis pathway in Brassica napus (canola) plants using RNAi. Can. J. Plant Sci. 2009, 89, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.R.; Yan, X.Y.; Ran, S.Y.; Ralph, J.; Smith, R.A.; Chen, X.P.; Qu, C.M.; Li, J.N.; Liu, L.Z. Knockout of the lignin pathway gene BnF5H decreases the S/G lignin compositional ratio and improves Sclerotinia sclerotiorum resistance in Brassica napus. Plant Cell Environ. 2022, 45, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Franke, R.; McMichael, C.M.; Meyer, K.; Shirley, A.M.; Cusumano, J.C.; Chapple, C. Modified lignin in tobacco and poplar plants over-expressing the Arabidopsis gene encoding ferulate 5-hydroxylase. Plant J. 2000, 22, 223–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Zhou, Y.H.; Cheng, X.F.; Sun, J.Y.; Marita, J.M.; Ralph, J.; Chiang, V.L. Combinatorial modification of multiple lignin traits in trees through multigene cotransformation. Proc. Natl. Acad. Sci. USA 2003, 100, 4939–4944. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Yang, D.H.; Kim, J.S.; Baek, M.H.; Park, Y.M.; Wi, S.G.; Cho, J.Y.; Chung, B.Y. Cloning, characterization, and expression of two cDNA clones for a rice ferulate-5-hydroxylase gene, a cytochrorne P450-dependent monooxygenase. J. Plant Biol. 2006, 49, 200–204. [Google Scholar] [CrossRef]
- Garcia, J.R.; Anderson, N.; Le-Feuvre, R.; Iturra, C.; Elissetche, J.; Chapple, C.; Valenzuela, S. Rescue of syringyl lignin and sinapate ester biosynthesis in Arabidopsis thaliana by a coniferaldehyde 5-hydroxylase from Eucalyptus globulus. Plant Cell Rep. 2014, 33, 1263–1274. [Google Scholar] [CrossRef]
- Humphreys, J.M.; Hemm, M.R.; Chapple, C. New routes for lignin biosynthesis defined by biochemical characterization of recombinant ferulate 5-hydroxylase, a multifunctional cytochrome P450-dependent monooxygenase. Proc. Natl. Acad. Sci. USA 1999, 96, 10045–10050. [Google Scholar] [CrossRef] [Green Version]
- Meyer, K.; Cusumano, J.C.; Somerville, C.; Chapple, C.C.S. Ferulate-5-hydroxylase from Arabidopsis thaliana defines a new family of cytochrome P450-dependent monooxygenases. Proc. Natl. Acad. Sci. USA 1996, 93, 6869–6874. [Google Scholar] [CrossRef] [Green Version]
- Meyer, K.; Shirley, A.M.; Cusumano, J.C.; Bell-Lelong, D.A.; Chapple, C. Lignin monomer composition is determined by the expression of a cytochrome P450-dependent monooxygenase in Arabidopsis. Proc. Natl. Acad. Sci. USA 1998, 95, 6619–6623. [Google Scholar] [CrossRef] [Green Version]
- Marita, J.M.; Ralph, J.; Hatfield, R.D.; Chapple, C. NMR characterization of lignins in Arabidopsis altered in the activity of ferulate 5-hydroxylase. Proc. Natl. Acad. Sci. USA 1999, 96, 12328–12332. [Google Scholar] [CrossRef] [Green Version]
- Sibout, R.; Baucher, M.; Gatineau, M.; Doorsselaere, J.V.; Mila, I.; Pollet, B.; Maba, B.; Pilate, G.; Lapierre, C.; Boerjan, W.; et al. Expression of a poplar cDNA encoding a ferulate-5-hydroxylase/coniferaldehyde 5-hydroxylase increases S lignin deposition in Arabidopsis thaliana. Plant Physiol. Biochem. 2002, 40, 1087–1096. [Google Scholar] [CrossRef]
- Wang, Z.W.; Hobson, N.; Galindo, L.; Zhu, S.L.; Shi, D.; McDill, J.; Yang, L.F.; Hawkins, S.; Neutelings, G.; Datla, R.; et al. The genome of flax (Linum usitatissimum) assembled de novo from short shotgun sequence reads. Plant J. Cell Mol. Biol. 2012, 72, 461–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, D.W.; Dai, Z.G.; Yang, Z.M.; Sun, J.; Zhao, D.B.; Yang, X.; Zhang, L.G.; Tang, Q.; Su, J.G. Genome-Wide Association Study Identifying Candidate Genes Influencing Important Agronomic Traits of Flax (Linum usitatissimum L.) Using SLAF-seq. Front. Plant Sci. 2018, 8, 22–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, D.L.; Jiang, H.X.; Yan, W.L.; Yang, L.J.; Ye, J.L.; Wang, Y.; Yan, Q.C.; Chen, J.X.; Gao, Y.F.; Duan, L.P. Resequencing 200 Flax Cultivated Accessions Identifies Candidate Genes Related to Seed Size and Weight and Reveals Signatures of Artificial Selection. Front. Plant Sci. 2020, 10, 1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tombuloglu, H. Genome-wide identification and expression analysis of R2R3, 3R- and 4R-MYB transcription factors during lignin biosynthesis in flax (Linum usitatissimum). Genomics 2020, 112, 782–795. [Google Scholar] [CrossRef]
- Gao, Y.; Chen, X.B.; Long, S.H.; Deng, X.; Zou, J.; Liu, A.L. Cloning and Sequence Analysis of Partial Fragments of Critical Genes Involved in Flax Lignin Synthesis. Acta Agron. Sin. 2008, 34, 337–340. [Google Scholar] [CrossRef]
- Jiang, W.T.; Zeng, Q.Q.; Jiang, Y.; Gai, Y.; Jiang, X.N. Molecular and functional characterization of ferulate-5-hydroxylase in Populus tomentosa. J. Plant Biochem. Biotechnol. 2021, 30, 92–98. [Google Scholar] [CrossRef]
- Franke, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, the Swiss bioinformatics resource portal, as designed by its users. Nucleic Acids Res. 2021, 49, 216–227. [Google Scholar] [CrossRef]
- Ayaz, A.; Saqib, S.; Huang, H.D.; Zaman, W.; Lü, S.Y.; Zhao, H.Y. Genome-wide comparative analysis of long-chain acyl-CoA synthetases (LACSs) gene family: A focus on identification, evolution and expression profiling related to lipid synthesis. Plant Physiol. Biochem. 2021, 161, 1–11. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef]
- Li, D.H.; Zaman, W.; Lu, J.J.; Niu, Q.Q.; Zhang, X.H.; Ayaz, A.; Saqib, S.; Yang, B.M.; Zhang, J.X.; Zhao, H.Y.; et al. Natural lupeol level variation among castor accessions and the upregulation of lupeol synthesis in response to light. Ind. Crops Prod. 2023, 192, 116090. [Google Scholar] [CrossRef]
- Shen, S.Y.; Zhang, Q.R.; Shi, Y.; Sun, Z.M.; Zhang, Q.Q.; Hou, S.J.; Wu, R.L.; Jiang, L.B.; Zhao, X.Y.; Guo, Y.Q. Genome-Wide Analysis of the NAC Domain Transcription Factor Gene Family in Theobroma cacao. Genes 2020, 11, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Peer, Y.V.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Li, R.Q.; Li, Y.R.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.L.; Wang, F.Q.; Li, D.H. Cloning and molecular characterization of a ferulate-5-hydroxylase gene from water chestnuts (Trapa bicornis Osbeck.). J. Plant Biochem. Biotechnol. 2018, 27, 139–146. [Google Scholar] [CrossRef]
- del Río, J.C.; Rencoret, J.; Marques, G.; Li, J.; Gellerstedt, G.; Jiménez-Barbero, J.; Martínez, A.T.; Gutiérrez, A. Structural characterization of the lignin from jute (Corchorus capsularis) fibers. J. Agric. Food Chem. 2009, 57, 10271–10281. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, A.; Rodríguez, I.M.; del Río, J.C. Chemical characterization of lignin and lipid fractions in industrial hemp bast fibers used for manufacturing high-quality paper pulps. J. Agric. Food Chem. 2006, 54, 2138–2144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- del Río, J.C.; Rencoret, J.; Gutiérrez, A.; Nieto, L.; Jiménez-Barbero, J.; Martínez, Á.T. Structural characterization of guaiacyl-rich lignins in flax (Linum usitatissimum) fibers and shives. J. Agric. Food Chem. 2011, 59, 11088–11099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapple, C. Molecular-genetic analysis of plant cytochrome P450-dependent monooxygenases. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 311–343. [Google Scholar] [CrossRef]
- Dueholm, B.; Krieger, C.; Drew, D.; Olry, A.; Kamo, T.; Taboureau, O.; Weitzel, C.; Bourgaud, F.; Hehn, A.; Simonsen, H.T. Evolution of substrate recognition sites (SRSs) in cytochromes P450 from Apiaceae exemplified by the CYP71AJ subfamily. BMC Evol. Biol. 2015, 15, 122. [Google Scholar] [CrossRef] [Green Version]
- Bai, S.; Yin, M.J.; Lyu, Q.; Jiang, B.; Li, L.X. Cytochrome P450 BsCYP99A44 and BsCYP704A177 Confer Metabolic Resistance to ALS Herbicides in Beckmannia syzigachne. Int. J. Mol. Sci. 2022, 23, 12175. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.P.; Edwards, M.; Wang, Q.Z.; Zhao, H.J.; Fu, H.W.; Huang, J.Z.; Gatehouse, A.; Shu, Q.Y. Expression of cytochrome P450 CYP81A6 in rice: Tissue specificity, protein subcellular localization, and response to herbicide application. J. Zhejiang Univ. Sci. B 2015, 16, 113–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruegger, M.; Meyer, K.; Cusumano, J.C.; Chapple, C. Regulation of Ferulate-5-Hydroxylase Expression in Arabidopsis in the Context of Sinapate Ester Biosynthesis. Plant Physiol. 1999, 119, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Neves, G.Y.S.; Marchiosi, R.; Ferrarese, M.L.L.; Siqueira-Soares, R.C.; Ferrarese-Filho, O. Root Growth Inhibition and Lignification Induced by Salt Stress in Soybean. J. Agron. Crop Sci. 2010, 196, 467–473. [Google Scholar] [CrossRef]
- Krishna, P. Brassinosteroid-Mediated Stress Responses. J. Plant Growth Regul. 2003, 22, 289–297. [Google Scholar] [CrossRef]
- Wang, L.; Xu, Y.Y.; Li, J.; Powell, R.A.; Xu, Z.H.; Chong, K. Transgenic rice plants ectopically expressing AtBAK1 are semi-dwarfed and hypersensitive to 24-epibrassinolide. J. Plant Physiol. 2007, 164, 655–664. [Google Scholar] [CrossRef]
- Mahesh, K.; Balaraju, P.; Ramakrishna, B.; Rao, S.S.R. Effect of brassinosteroids on germination and seedling growth of radish (Raphanus sativus L.) under PEG-6000 induced water stress. Am. J. Plant Sci. 2013, 4, 2305–2313. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Kumar, A.; Bhardwaj, R. Plant steroidal hormone epibrassinolide regulate- heavy metal stress tolerance in Oryza sativa L. by modulating antioxidant defense expression. Environ. Exp. Bot. 2016, 122, 1–9. [Google Scholar] [CrossRef]
- Otie, V.; Udo, I.; Shao, Y.; Itam, M.; Okamoto, H.; An, P.; Eneji, E. Salinity effects on morpho-physiological and yield traits of soybean (Glycine max L.) as mediated by foliar spray with brassinolide. Plants 2021, 10, 541. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | aa | MV (Da) | pI | GRAVY | Aliphatic Index | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| LuF5H1 | Lus10014273 | 515 | 57,508.51 | 5.75 | −0.087 | 89.20 | PM |
| LuF5H2 | Lus10041511 | 493 | 55,865.44 | 6.37 | −0.210 | 85.45 | PM |
| LuF5H3 | Lus10028361 | 512 | 57,743.95 | 6.07 | −0.111 | 92.54 | PM |
| LuF5H4 | Lus10025975 | 510 | 57,051.18 | 6.16 | −0.072 | 89.69 | PM |
| LuF5H5 | Lus10012582 | 470 | 53,319.63 | 6.68 | −0.177 | 85.67 | PM |
| LuF5H6 | Lus10022303 | 510 | 57,656.91 | 6.43 | −0.096 | 91.98 | PM |
| LuF5H7 | Lus10041811 | 485 | 54,456.20 | 5.99 | −0.079 | 95.10 | PM |
| LuF5H8 | Lus10034300 | 513 | 58,072.41 | 6.68 | −0.054 | 90.68 | PM |
| LuF5H9 | Lus10000326 | 388 | 43,893.39 | 5.12 | −0.184 | 84.68 | PM |
| LuF5H1 | LuF5H3 | LuF5H4 | LuF5H7 | Lignin Content | |
|---|---|---|---|---|---|
| LuF5H1 | 1 | 0.090 | 0.982 ** | 0.103 | 0.475 |
| LuF5H3 | 0.090 | 1 | −0.204 | 0.982 ** | 0.478 |
| LuF5H4 | 0.982 ** | −0.204 | 1 | −0.003 | 0.453 |
| LuF5H7 | 0.103 | 0.982 ** | −0.003 | 1 | 0.591 |
| lignin content | 0.475 | 0.478 | 0.453 | 0.591 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Yuan, H.; Yao, Y.; Cheng, L.; Tang, L.; Kang, Q.; Song, X.; Chen, S.; Wu, G. Genome-Wide Identification and Expression Pattern Analysis of the F5H Gene Family in Flax (Linum usitatissimum L.). Agronomy 2023, 13, 1108. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13041108
Liu D, Yuan H, Yao Y, Cheng L, Tang L, Kang Q, Song X, Chen S, Wu G. Genome-Wide Identification and Expression Pattern Analysis of the F5H Gene Family in Flax (Linum usitatissimum L.). Agronomy. 2023; 13(4):1108. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13041108
Chicago/Turabian StyleLiu, Dandan, Hongmei Yuan, Yubo Yao, Lili Cheng, Lili Tang, Qinghua Kang, Xixia Song, Si Chen, and Guangwen Wu. 2023. "Genome-Wide Identification and Expression Pattern Analysis of the F5H Gene Family in Flax (Linum usitatissimum L.)" Agronomy 13, no. 4: 1108. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13041108