
Determination of Physiochemical Characteristics Associated with Various Degrees of Cadmium Tolerance in Barley Accessions

 ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Germination Test and Phenotypic Character Measurements under Cd Stress Conditions
2.3. Physiochemical Measurements
2.4. Statistical Data Analysis
3. Results
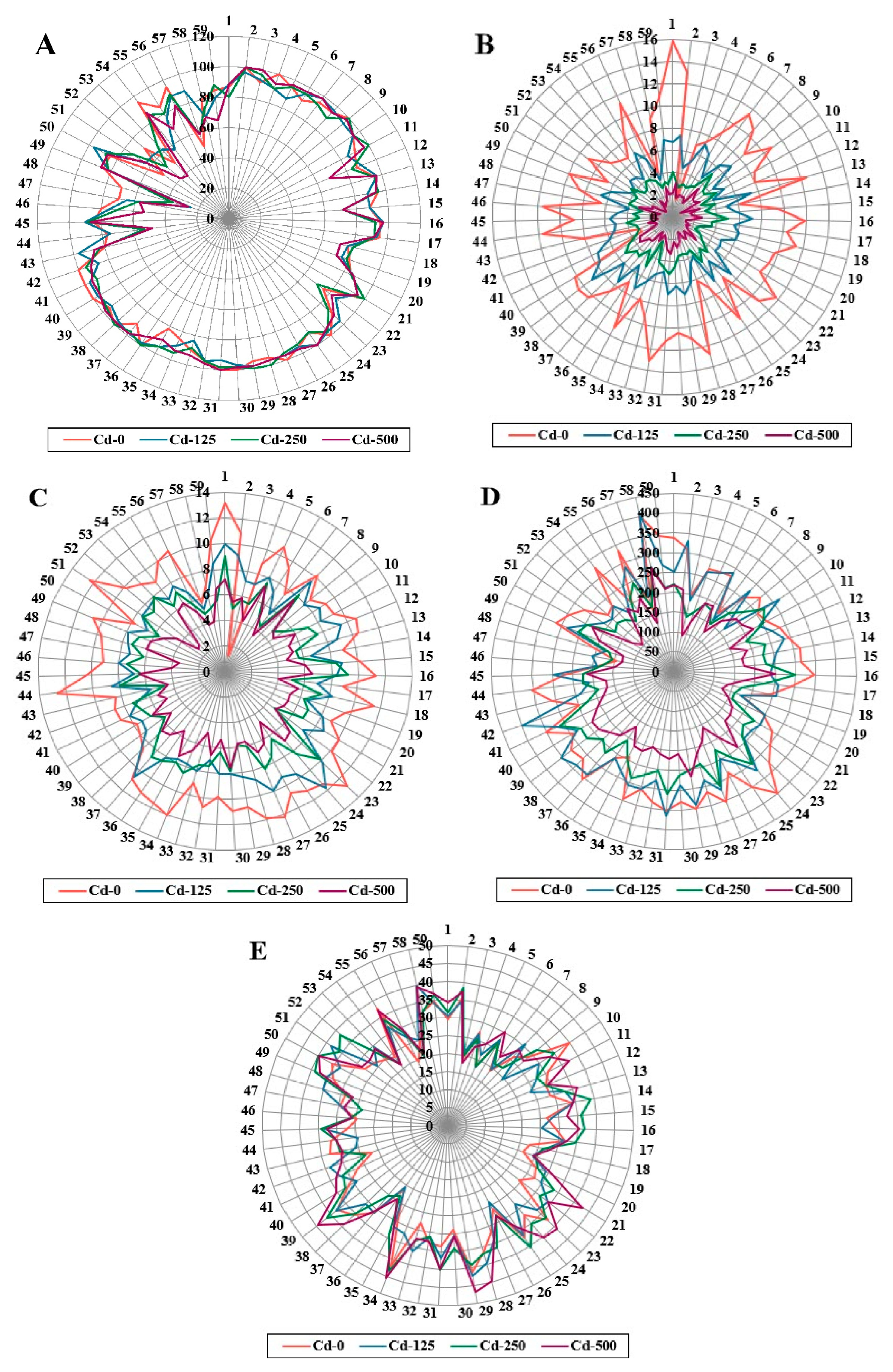
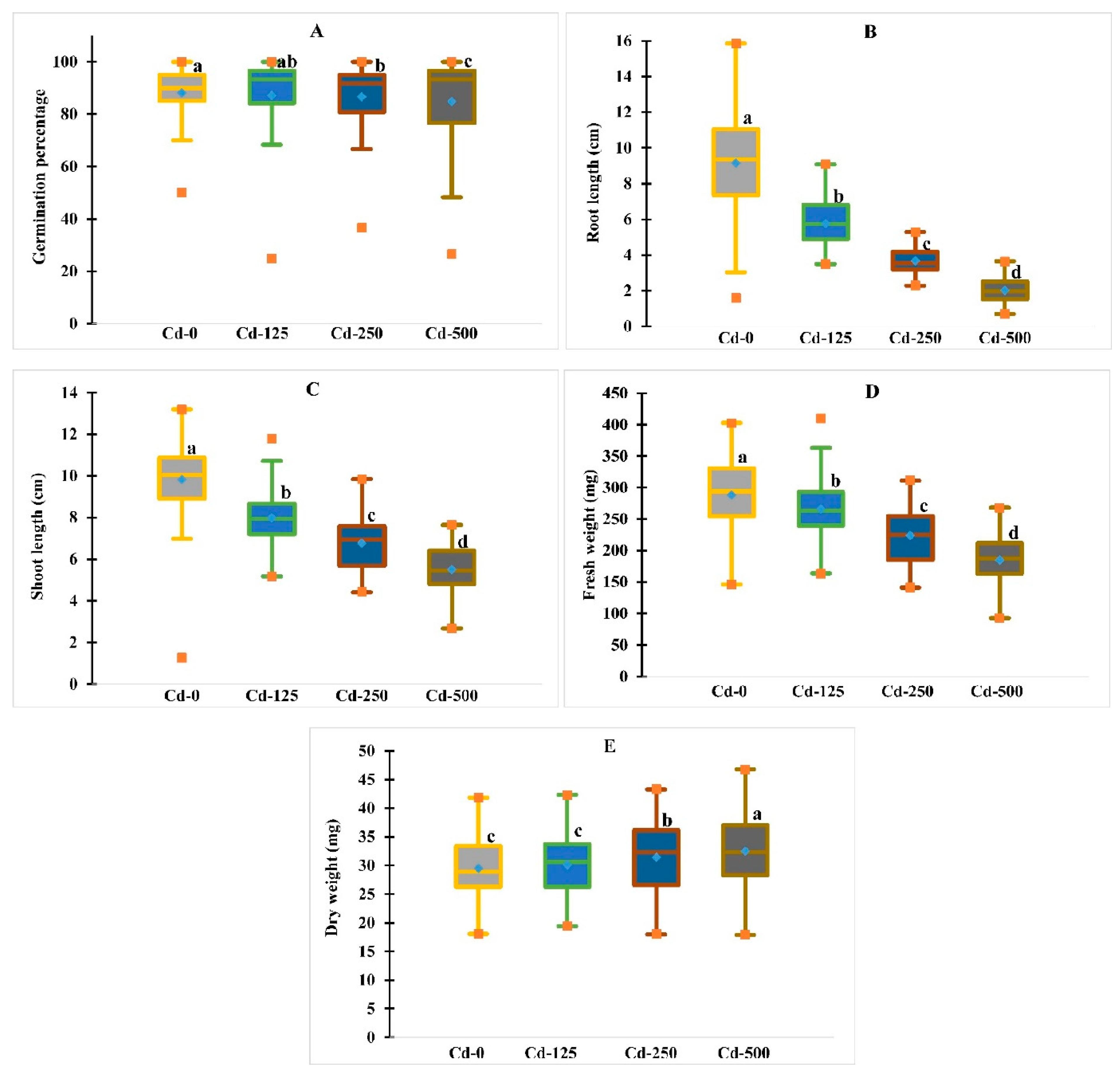
3.1. Performance of Morphological Traits under Cadmium Heavy Metal Stress Conditions
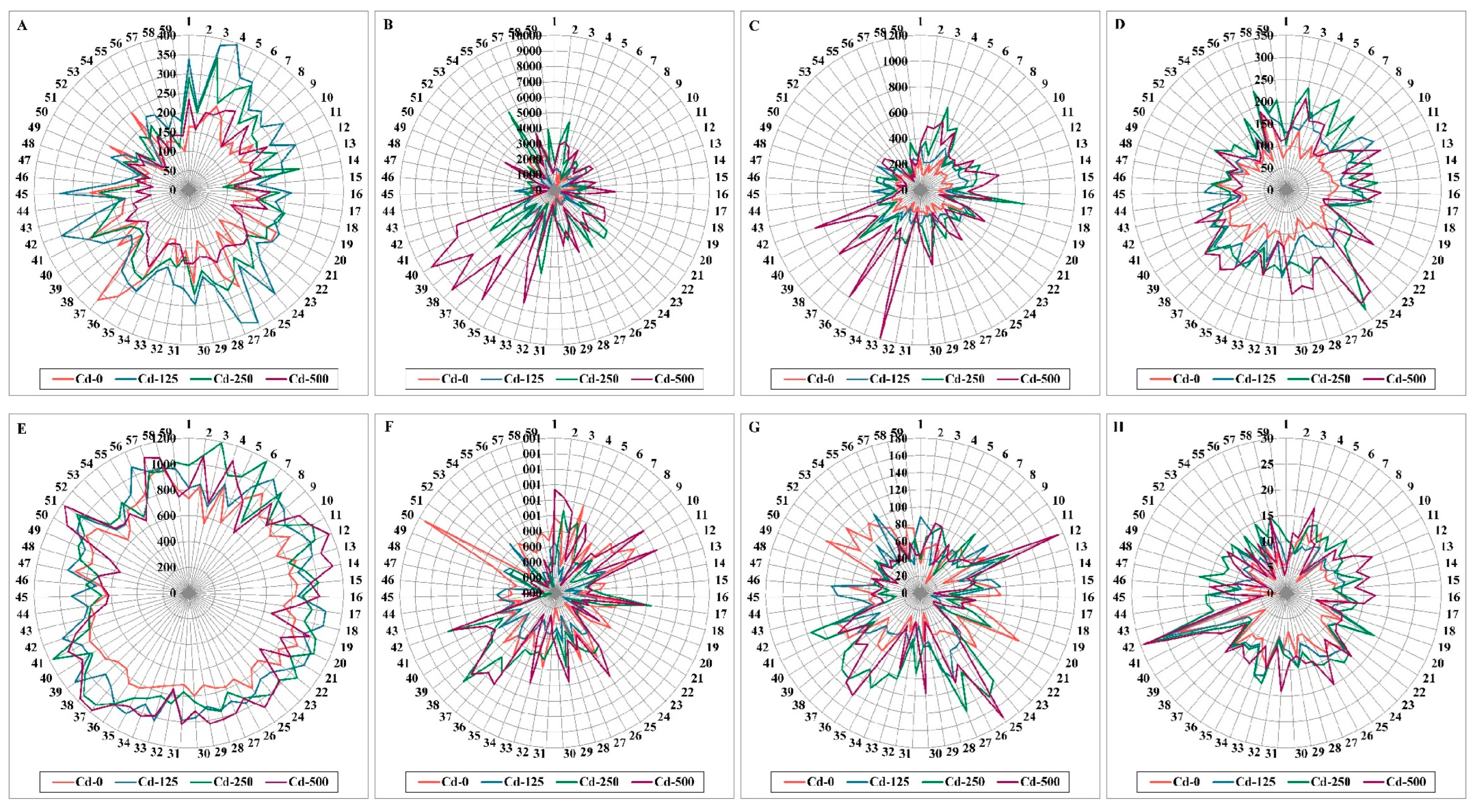
3.2. Physio-Biochemical Markers Assays
3.3. Ranking of Barley Accessions for Morphological Traits under Cadmium Stress Conditions
3.4. Relationship between Tolerance Degree and Physiochemical Traits
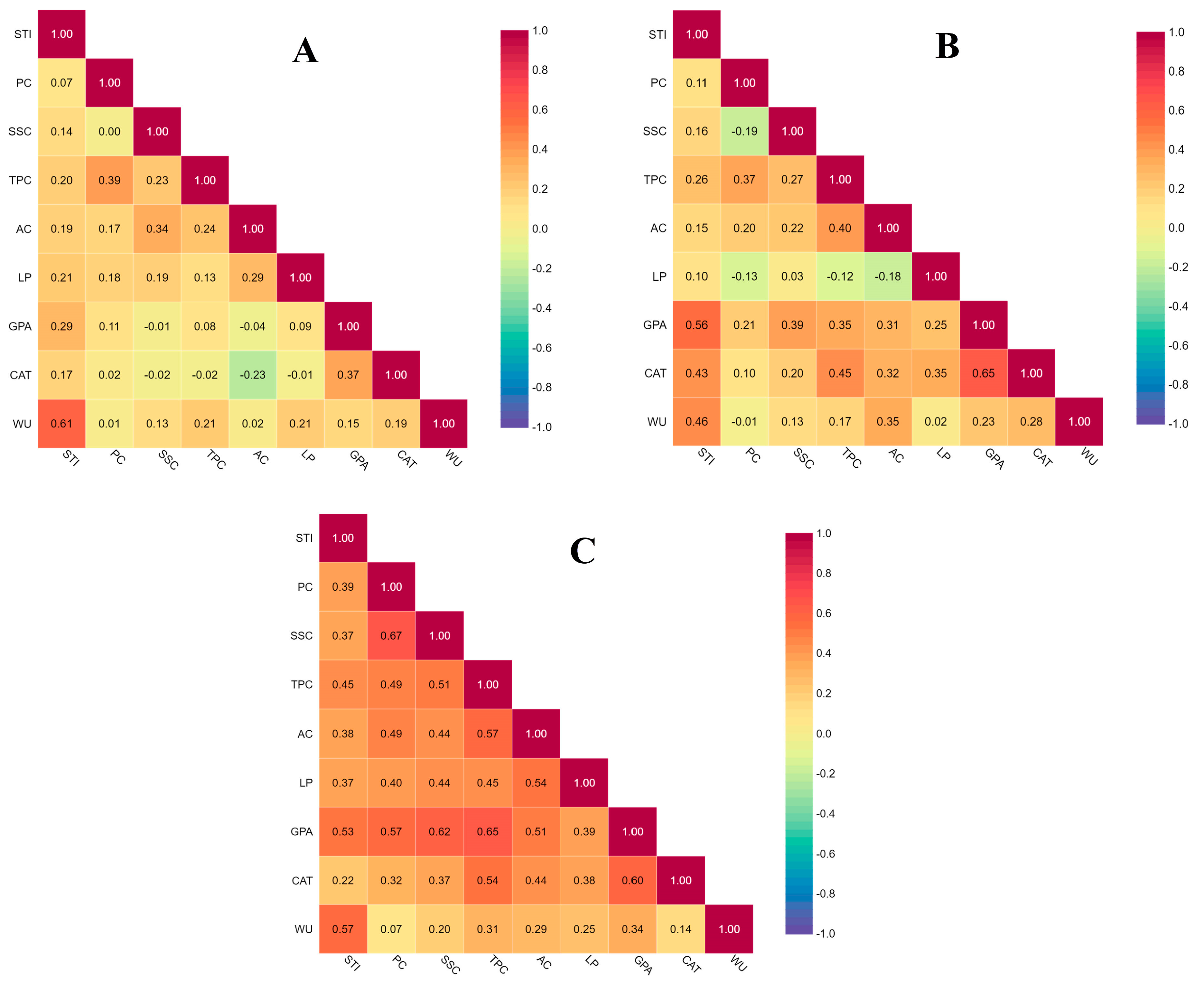
3.5. Correlations among Measured Traits under Three Different Concentrations of Cadmium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vareda, J.P.; Valente, A.J.M.; Durães, L. Assessment of heavy metal pollution from anthropogenic activities and remediation strategies: A review. J. Environ. Manag. 2019, 246, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Osmolovskaya, N.G.; Dung, V.V.; Kudryashova, Z.K.; Kuchaeva, L.N.; Popova, N.F. Effect of cadmium on distribution of potassium, calcium, magnesium, and oxalate accumulation in Amaranthus cruentus L. Plants. Russ. J. Plant Physiol. 2018, 65, 553–562. [Google Scholar] [CrossRef]
- Qin, S.; Liu, H.; Nie, Z.; Rengel, Z.; Gao, W.; Li, C.; Zhao, P. Toxicity of cadmium and its competition with mineral nutrients for uptake by plants: A review. Pedosphere 2020, 30, 168–180. [Google Scholar] [CrossRef]
- Islam, M.; Sandhi, A. Heavy Metal and Drought Stress in Plants: The Role of Microbes—A Review. Gesunde Pflanz. 2022, 1–14. [Google Scholar] [CrossRef]
- Berni, R.; Luyckx, M.; Xu, X.; Legay, S.; Sergeant, K.; Hausman, J.-F.; Lutts, S.; Cai, G.; Guerriero, G. Reactive oxygen species and heavy metal stress in plants: Impact on the cell wall and secondary metabolism. Environ. Exp. Bot. 2019, 161, 98–106. [Google Scholar] [CrossRef]
- Tan, Y.; Duan, Y.; Chi, Q.; Wang, R.; Yin, Y.; Cui, D.; Li, S.; Wang, A.; Ma, R.; Li, B.; et al. The role of reactive oxygen species in plant response to radiation. Int. J. Mol. Sci. 2023, 24, 3346. [Google Scholar] [CrossRef]
- Zandi, P.; Schnug, E. Reactive oxygen species, antioxidant responses and implications from a microbial modulation perspective. Biology 2022, 11, 155. [Google Scholar] [CrossRef]
- Berwal, M.K.; Haldhar, S.M.; Ram, C.; Shil, S.; Kumar, R.; Gora, J.S.; Singh, D.; Samadia, D.K.; Kumar, M.; Mekhemar, M. Calligonum polygonoides L. as novel source of bioactive compounds in hot arid regions: Evaluation of phytochemical composition and antioxidant activity. Plants 2021, 10, 1156. [Google Scholar] [CrossRef]
- Thakur, M.; Praveen, S.; Divte, P.R.; Mitra, R.; Kumar, M.; Gupta, C.K.; Kalidindi, U.; Bansal, R.; Roy, S.; Anand, A.; et al. Metal tolerance in plants: Molecular and physicochemical interface determines the “not so heavy effect” of heavy metals. Chemosphere 2022, 287, 131957. [Google Scholar] [CrossRef]
- Irato, P.; Santovito, G. Enzymatic and Non-Enzymatic Molecules with Antioxidant Function. Antioxidants 2021, 10, 579. [Google Scholar] [CrossRef]
- Delangiz, N.; Khoshru, B.; Asgari Lajayer, B.; Ghorbanpour, M.; Kazemalilou, S. Molecular Mechanisms of Heavy Metal Tolerance in Plants. In Cellular and Molecular Phytotoxicity of Heavy Metals; Springer: Cham, Switzerland, 2020; pp. 125–136. [Google Scholar]
- Seregin, I.V.; Kozhevnikova, A.D. Phytochelatins: Sulfur-Containing Metal(loid)-Chelating Ligands in Plants. Int. J. Mol. Sci. 2023, 24, 2430. [Google Scholar] [CrossRef] [PubMed]
- Kushwaha, A.; Rani, R.; Kumar, S.; Gautam, A. Heavy metal detoxification and tolerance mechanisms in plants: Implications for phytoremediation. Environ. Rev. 2016, 24, 39–51. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nozoye, T.; Nishizawa, N.K. Iron transport and its regulation in plants. Free Radic. Biol. Med. 2019, 133, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Wang, K.; He, X.; Yang, Z.; Wang, L. Characterization of physicochemical parameters and bioavailable heavy metals and their interactions with microbial community in arsenic-contaminated soils and sediments. Environ. Sci. Pollut. Res. 2022, 29, 49672–49683. [Google Scholar] [CrossRef]
- Guo, F.; Ding, C.; Zhou, Z.; Huang, G.; Wang, X. Effects of combined amendments on crop yield and cadmium uptake in two cadmium contaminated soils under rice-wheat rotation. Ecotoxicol. Environ. Saf. 2018, 148, 303–310. [Google Scholar] [CrossRef]
- Dawood, M.F.A.; Tahjib-Ul-Arif, M.; Sohag, A.A.M.; Abdel Latef, A.A.H. Fluoride mitigates aluminum-toxicity in barley: Morpho-physiological responses and biochemical mechanisms. BMC Plant Biol. 2022, 22, 287. [Google Scholar] [CrossRef]
- Pérez-García, L.-A.; Sáenz-Mata, J.; Fortis-Hernández, M.; Navarro-Muñoz, C.E.; Palacio-Rodríguez, R.; Preciado-Rangel, P. Plant-growth-promoting rhizobacteria improve germination and bioactive compounds in cucumber seedlings. Agronomy 2023, 13, 315. [Google Scholar] [CrossRef]
- Huybrechts, M.; Cuypers, A.; Deckers, J.; Iven, V.; Vandionant, S.; Jozefczak, M.; Hendrix, S. Cadmium and plant development: An agony from seed to seed. Int. J. Mol. Sci. 2019, 20, 3971. [Google Scholar] [CrossRef]
- Zulfiqar, U.; Jiang, W.; Xiukang, W.; Hussain, S.; Ahmad, M.; Maqsood, M.F.; Ali, N.; Ishfaq, M.; Kaleem, M.; Haider, F.U.; et al. Cadmium phytotoxicity, tolerance, and advanced remediation approaches in agricultural soils; A comprehensive review. Front. Plant Sci. 2022, 13, 773815. [Google Scholar] [CrossRef]
- Demecsová, L.; Zelinová, V.; Liptáková, Ľ.; Tamás, L. Mild cadmium stress induces auxin synthesis and accumulation, while severe cadmium stress causes its rapid depletion in barley root tip. Environ. Exp. Bot. 2020, 175, 104038. [Google Scholar] [CrossRef]
- Jawad Hassan, M.; Ali Raza, M.; Ur Rehman, S.; Ansar, M.; Gitari, H.; Khan, I.; Wajid, M.; Ahmed, M.; Abbas Shah, G.; Peng, Y.; et al. Effect of cadmium toxicity on growth, oxidative damage, antioxidant defense system and cadmium accumulation in two sorghum cultivars. Plants 2020, 9, 1575. [Google Scholar] [CrossRef] [PubMed]
- Lateef, D.; Mustafa, K.; Tahir, N. Screening of Iraqi barley accessions under PEG-induced drought conditions. All Life 2021, 14, 308–332. [Google Scholar] [CrossRef]
- Rasul, K.S.; Grundler, F.M.W.; Abdul-razzak Tahir, N. Genetic diversity and population structure assessment of Iraqi tomato accessions using fruit characteristics and molecular markers. Hortic. Environ. Biotechnol. 2022, 63, 523–538. [Google Scholar] [CrossRef]
- Tahir, N.A.; Ahmed, J.O.; Azeez, H.A.; Palani, W.R.M.; Omer, D.A. Phytochemical, antibacterial, antioxidant and phytotoxicity screening of the extracts collected from the fruit and root of wild Mt. Atlas mastic tree (Pistacia atlantica subsp. kurdica). Appl. Ecol. Environ. Res. 2019, 17, 4417–4429. [Google Scholar] [CrossRef]
- Tahir, N.A.; Rasul, K.S.; Lateef, D.D.; Grundler, F.M.W. Effects of Oak Leaf Extract, Biofertilizer, and Soil Containing Oak Leaf Powder on Tomato Growth and Biochemical Characteristics under Water Stress Conditions. Agriculture 2022, 12, 2082. [Google Scholar] [CrossRef]
- Ketata, H.Y.; Yau, S.K.; Nachit, M. Relative consistency performance across environments. In Proceedings of the International Symposium on Physiology and Breeding of Winter Cereals for stressed Mediterranean Environments, Montpellier, France, 3–6 July 1989; pp. 391–400. [Google Scholar]
- Pour-Aboughadareh, A.; Yousefian, M.; Moradkhani, H.; Moghaddam Vahed, M.; Poczai, P.; Siddique, K.H.M. iPASTIC: An online toolkit to estimate plant abiotic stress indices. Appl. Plant Sci. 2019, 7, e11278. [Google Scholar] [CrossRef]
- Verheijen, M.; Tong, W.; Shi, L.; Gant, T.W.; Seligman, B.; Caiment, F. Towards the development of an omics data analysis framework. Regul. Toxicol. Pharmacol. 2020, 112, 104621. [Google Scholar] [CrossRef]
- Bianchi, L.; Sframeli, M.; Vantaggiato, L.; Vita, G.L.; Ciranni, A.; Polito, F.; Oteri, R.; Gitto, E.; Di Giuseppe, F.; Angelucci, S.; et al. Nusinersen Modulates Proteomics Profiles of Cerebrospinal Fluid in Spinal Muscular Atrophy Type 1 Patients. Int. J. Mol. Sci. 2021, 22, 4329. [Google Scholar] [CrossRef]
- El Rasafi, T.; Oukarroum, A.; Haddioui, A.; Song, H.; Kwon, E.E.; Bolan, N.; Tack, F.M.G.; Sebastian, A.; Prasad, M.N.V.; Rinklebe, J. Cadmium stress in plants: A critical review of the effects, mechanisms, and tolerance strategies. Crit. Rev. Environ. Sci. Technol. 2022, 52, 675–726. [Google Scholar] [CrossRef]
- Haider, F.U.; Liqun, C.; Coulter, J.A.; Cheema, S.A.; Wu, J.; Zhang, R.; Wenjun, M.; Farooq, M. Cadmium toxicity in plants: Impacts and remediation strategies. Ecotoxicol. Environ. Saf. 2021, 211, 111887. [Google Scholar] [CrossRef]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The chemistry of reactive oxygen species (ROS) revisited: Outlining their role in biological macromolecules (DNA, lipids and proteins) and induced pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Dayem, A.A.; Hossain, M.K.; Lee, S.B.; Kim, K.; Saha, S.K.; Yang, G.-M.; Choi, H.Y.; Cho, S.-G. The Role of Reactive Oxygen Species (ROS) in the Biological Activities of Metallic Nanoparticles. Int. J. Mol. Sci. 2017, 18, 120. [Google Scholar] [CrossRef] [PubMed]
- Dalla Vecchia, F.; La Rocca, N.; Moro, I.; de Faveri, S.; Andreoli, C.; Rascio, N. Morphogenetic, ultrastructural and physiological damages suffered by submerged leaves of Elodea canadensis exposed to cadmium. Plant Sci. 2005, 168, 329–338. [Google Scholar] [CrossRef]
- Abbas, T.; Rizwan, M.; Ali, S.; Adrees, M.; Zia-Ur-Rehman, M.; Qayyum, M.F.; Ok, Y.S.; Murtaza, G. Effect of biochar on alleviation of cadmium toxicity in wheat (Triticum aestivum L.) grown on Cd-contaminated saline soil. Environ. Sci. Pollut. Res. 2018, 25, 25668–25680. [Google Scholar] [CrossRef] [PubMed]
- Abeed, A.H.A.; Mahdy, R.E.; Alshehri, D.; Hammami, I.; Eissa, M.A.; Abdel Latef, A.A.H.; Mahmoud, G.A.-E. Induction of resilience strategies against biochemical deteriorations prompted by severe cadmium stress in sunflower plant when Trichoderma and bacterial inoculation were used as biofertilizers. Front. Plant Sci. 2022, 13, 1004173. [Google Scholar] [CrossRef] [PubMed]
- Badawy, I.H.; Hmed, A.A.; Sofy, M.R.; Al-Mokadem, A.Z. Alleviation of cadmium and nickel toxicity and phyto-stimulation of tomato plant L. by Endophytic Micrococcus luteus and Enterobacter cloacae. Plants 2022, 11, 2018. [Google Scholar] [CrossRef]
- Carvalho, M.E.; Piotto, F.A.; Franco, M.R.; Borges, K.L.; Gaziola, S.A.; Castro, P.R.; Azevedo, R.A. Cadmium toxicity degree on tomato development is associated with disbalances in B and Mn status at early stages of plant exposure. Ecotoxicology 2018, 27, 1293–1302. [Google Scholar] [CrossRef]
- Carvalho, M.E.; Piotto, F.A.; Gaziola, S.A.; Jacomino, A.P.; Jozefczak, M.; Cuypers, A.; Azevedo, R.A. New insights about cadmium impacts on tomato: Plant acclimation, nutritional changes, fruit quality and yield. Food Energy Secur. 2018, 7, e00131. [Google Scholar] [CrossRef]
- Naciri, R.; Lahrir, M.; Benadis, C.; Chtouki, M.; Oukarroum, A. Interactive effect of potassium and cadmium on growth, root morphology and chlorophyll a fluorescence in tomato plant. Sci. Rep. 2021, 11, 5384. [Google Scholar] [CrossRef]
- Dad, K.; Nawaz, M.; Hassan, R.; Javed, K.; Shaheen, A.; Zhao, F.; Imran, M.; Shah, S.T.H.; Anwar, M.F.; Aurangzaib, M. Impact of biochar on the growth and physiology of tomato grown in the cadmium contaminated soil. Pak. J. Agric. Sci. 2021, 34, 454–462. [Google Scholar] [CrossRef]
- Abeed, A.H.A.; Salama, F.M. Attenuating Effect of an Extract of Cd-Hyperaccumulator Solanum nigrum on the Growth and Physio-chemical Changes of Datura innoxia Under Cd Stress. J. Soil Sci. Plant Nutr. 2022, 22, 4868–4882. [Google Scholar] [CrossRef]
- Cong, L.; Yu, L.; Jing, T.; Yanshu, Z.; Jinjuan, F. Changes in sucrose metabolism in maize varieties with different cadmium sensitivities under cadmium stress. PLoS ONE 2020, 15, e0243835. [Google Scholar]
- Yuan, H.M.; Huang, X. Inhibition of root meristem growth by cadmium involves nitric oxide-mediated repression of auxin accumulation and signalling in Arabidopsis. Plant Cell Environ. 2016, 39, 120–135. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Habib, M.; Kakavand, S.N.; Zahid, Z.; Zahra, N.; Sharif, R.; Hasanuzzaman, M. Phytoremediation of cadmium: Physiological, biochemical, and molecular mechanisms. Biology 2020, 9, 177. [Google Scholar] [CrossRef]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 52, 39. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Raihan, M.R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of Reactive Oxygen Species and Antioxidant Defense in Plants under Salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef]
- Balali-Mood, M.; Naseri, K.; Tahergorabi, Z.; Khazdair, M.R.; Sadeghi, M. Toxic mechanisms of five heavy metals: Mercury, lead, chromium, cadmium, and arsenic. Front. Pharmacol. 2021, 12, 643972. [Google Scholar] [CrossRef]
- Kaur, H.; Nazir, F.; Hussain, S.J.; Kaur, R.; Rajurkar, A.B.; Kumari, S.; Siddiqui, M.H.; Mahajan, M.; Khatoon, S.; Khan, M.I.R. Gibberellic acid alleviates cadmium-induced seed germination inhibition through modulation of carbohydrate metabolism and antioxidant capacity in mung bean seedlings. Sustainability 2023, 15, 3790. [Google Scholar] [CrossRef]
- Nouri, M.; El Rasafi, T.; Haddioui, A. Responses of two barley subspecies to In vitro- induced heavy metal stress: Seeds germination, seedlings growth and cytotoxicity assay. Agriculture 2019, 65, 107–118. [Google Scholar] [CrossRef]
- Ali, J.; Ali, F.; Ahmad, I.; Rafique, M.; Munis, M.F.H.; Hassan, S.W.; Sultan, T.; Iftikhar, M.; Chaudhary, H.J. Mechanistic elucidation of germination potential and growth of Sesbania sesban seedlings with Bacillus anthracis PM21 under heavy metals stress: An in vitro study. Ecotoxicol. Environ. Saf. 2021, 208, 111769. [Google Scholar] [CrossRef]
- Spormann, S.; Nadais, P.; Sousa, F.; Pinto, M.; Martins, M.; Sousa, B.; Fidalgo, F.; Soares, C. Accumulation of proline in plants under contaminated soils-Are We on the same page? Antioxidants 2023, 12, 666. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J.; Cano, A.; Reiter, R.J. Melatonin and Carbohydrate Metabolism in Plant Cells. Plants 2021, 10, 1917. [Google Scholar] [CrossRef] [PubMed]
- Jeandet, P.; Formela-Luboińska, M.; Labudda, M.; Morkunas, I. The Role of Sugars in Plant Responses to Stress and Their Regulatory Function during Development. Int. J. Mol. Sci. 2022, 23, 5161. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, Q.; Lu, H.; Li, J.; Yang, D.; Liu, J.; Yan, C. Phenolic metabolism and related heavy metal tolerance mechanism in Kandelia Obovata under Cd and Zn stress. Ecotoxicol. Environ. Saf. 2019, 169, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Chao, Y.-Y.; Chen, C.-Y.; Huang, W.-D.; Kao, C.H. Salicylic acid-mediated hydrogen peroxide accumulation and protection against Cd toxicity in rice leaves. Plant Soil 2010, 329, 327–337. [Google Scholar] [CrossRef]
- Jiang, S.; Weng, B.; Liu, T.; Su, Y.; Liu, J.; Lu, H.; Yan, C. Response of phenolic metabolism to cadmium and phenanthrene and its influence on pollutant translocations in the mangrove plant Aegiceras corniculatum (L.) Blanco (Ac). Ecotoxicol. Environ. Saf. 2017, 141, 290–297. [Google Scholar] [CrossRef]
- Luo, J.-S.; Zhang, Z. Mechanisms of cadmium phytoremediation and detoxification in plants. Crop. J. 2021, 9, 521–529. [Google Scholar] [CrossRef]
- Galić, V.; Mlinarić, S.; Marelja, M.; Zdunić, Z.; Brkić, A.; Mazur, M.; Begović, L.; Šimić, D. Contrasting water withholding responses of young maize plants reveal link between lipid peroxidation and osmotic regulation corroborated by genetic analysis. Front. Plant Sci. 2022, 13, 2099. [Google Scholar] [CrossRef]
- Zhao, H.; Guan, J.; Liang, Q.; Zhang, X.; Hu, H.; Zhang, J. Effects of cadmium stress on growth and physiological characteristics of sassafras seedlings. Sci. Rep. 2021, 11, 9913. [Google Scholar] [CrossRef]
- Waheed, A.; Haxim, Y.; Islam, W.; Ahmad, M.; Ali, S.; Wen, X.; Khan, K.A.; Ghramh, H.A.; Zhang, Z.; Zhang, D. Impact of cadmium stress on growth and physio-biochemical attributes of Eruca sativa Mill. Plants 2022, 11, 2981. [Google Scholar] [CrossRef]
- Kintlová, M.; Blavet, N.; Cegan, R.; Hobza, R. Transcriptome of barley under three different heavy metal stress reaction. Genomics Data 2017, 13, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Kalai, T.; Khamassi, K.; Da Teixeira Silva, J.A.; Gouia, H.; Bettaieb Ben-Kaab, L. Cadmium and copper stress affect seedling growth and enzymatic activities in germinating barley seeds. Arch. Agron. Soil Sci. 2014, 60, 765–783. [Google Scholar] [CrossRef]
- Kintlová, M.; Vrána, J.; Hobza, R.; Blavet, N.; Hudzieczek, V. Transcriptome response to cadmium exposure in barley (Hordeum vulgare L.). Front. Plant Sci. 2021, 12, 629089. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Code | ASR | STI | Rank | Accession Code | ASR | STI | Rank | Accession Code | ASR | STI | Rank |
|---|---|---|---|---|---|---|---|---|---|---|---|
| AC29 | 10.09 | 1.16 | 1 | AC26 | 22.91 | 1.06 | 21 | AC14 | 37.00 | 0.96 | 41 |
| AC24 | 10.64 | 1.17 | 2 | AC7 | 23.82 | 1.07 | 22 | AC53 | 37.73 | 0.43 | 42 |
| AC25 | 10.73 | 1.14 | 3 | AC21 | 24.36 | 1.13 | 23 | AC11 | 38.18 | 0.92 | 43 |
| AC37 | 11.82 | 1.11 | 4 | AC50 | 24.45 | 0.93 | 24 | AC56 | 39.00 | 1.02 | 44 |
| AC23 | 11.91 | 1.13 | 5 | AC10 | 24.91 | 1.08 | 25 | AC51 | 39.09 | 0.47 | 45 |
| AC13 | 13.00 | 1.16 | 6 | AC43 | 25.27 | 0.91 | 26 | AC42 | 39.27 | 0.97 | 46 |
| AC8 | 14.64 | 1.13 | 7 | AC31 | 26.00 | 1.14 | 27 | AC49 | 42.73 | 0.88 | 47 |
| AC16 | 14.64 | 1.15 | 8 | AC34 | 27.27 | 0.95 | 28 | AC33 | 42.82 | 0.91 | 48 |
| AC2 | 15.45 | 1.22 | 9 | AC3 | 28.18 | 0.86 | 29 | AC18 | 43.36 | 0.79 | 49 |
| AC28 | 16.00 | 1.16 | 10 | AC12 | 28.45 | 0.95 | 30 | AC59 | 43.73 | 0.88 | 50 |
| AC36 | 16.36 | 1.15 | 11 | AC17 | 29.00 | 1.05 | 31 | AC55 | 48.55 | 0.82 | 51 |
| AC41 | 16.36 | 1.09 | 12 | AC32 | 29.09 | 1.11 | 32 | AC44 | 50.18 | 0.78 | 52 |
| AC40 | 17.27 | 1.15 | 13 | AC45 | 29.45 | 0.93 | 33 | AC19 | 50.73 | 0.75 | 53 |
| AC38 | 17.64 | 1.15 | 14 | AC15 | 30.00 | 0.97 | 34 | AC46 | 51.27 | 0.77 | 54 |
| AC30 | 17.91 | 1.17 | 15 | AC1 | 31.09 | 1.06 | 35 | AC47 | 51.64 | 0.57 | 55 |
| AC27 | 18.09 | 1.15 | 16 | AC22 | 33.27 | 0.82 | 36 | AC54 | 53.55 | 0.65 | 56 |
| AC39 | 19.73 | 1.09 | 17 | AC20 | 33.91 | 0.90 | 37 | AC52 | 53.82 | 0.58 | 57 |
| AC35 | 20.82 | 1.00 | 18 | AC57 | 34.09 | 0.50 | 38 | AC58 | 54.09 | 0.66 | 58 |
| AC6 | 21.45 | 1.02 | 19 | AC5 | 35.73 | 1.00 | 39 | AC48 | 58.55 | 0.30 | 59 |
| AC9 | 22.18 | 1.12 | 20 | AC4 | 36.73 | 1.03 | 40 |
| Accession Code | ASR | STI | Rank | Accession Code | ASR | STI | Rank | Accession Code | ASR | STI | Rank |
|---|---|---|---|---|---|---|---|---|---|---|---|
| AC36 | 7.18 | 1.15 | 1 | AC26 | 22.45 | 1.02 | 21 | AC53 | 36.55 | 0.45 | 41 |
| AC38 | 9.45 | 1.15 | 2 | AC16 | 23.00 | 1.09 | 22 | AC20 | 37.00 | 0.85 | 42 |
| AC11 | 12.00 | 1.05 | 3 | AC14 | 23.55 | 0.99 | 23 | AC4 | 37.00 | 0.99 | 43 |
| AC30 | 12.27 | 1.17 | 4 | AC6 | 23.82 | 0.97 | 24 | AC57 | 38.36 | 0.34 | 44 |
| AC29 | 12.45 | 1.11 | 5 | AC3 | 24.45 | 0.86 | 25 | AC18 | 38.73 | 0.79 | 45 |
| AC21 | 12.91 | 1.14 | 6 | AC10 | 25.18 | 1.04 | 26 | AC12 | 39.91 | 0.86 | 46 |
| AC37 | 12.91 | 1.05 | 7 | AC13 | 26.36 | 1.07 | 27 | AC54 | 40.91 | 0.91 | 47 |
| AC28 | 13.45 | 1.14 | 8 | AC8 | 27.45 | 1.04 | 28 | AC1 | 41.91 | 0.94 | 48 |
| AC31 | 13.91 | 1.16 | 9 | AC41 | 28.00 | 0.99 | 29 | AC49 | 43.36 | 0.85 | 49 |
| AC24 | 14.55 | 1.12 | 10 | AC33 | 29.82 | 0.94 | 30 | AC55 | 44.00 | 0.82 | 50 |
| AC39 | 14.55 | 1.07 | 11 | AC50 | 30.36 | 0.84 | 31 | AC19 | 46.45 | 0.75 | 51 |
| AC9 | 15.09 | 1.10 | 12 | AC45 | 30.64 | 0.88 | 32 | AC15 | 49.64 | 0.79 | 52 |
| AC42 | 19.27 | 1.03 | 13 | AC25 | 31.27 | 0.99 | 33 | AC58 | 50.73 | 0.68 | 53 |
| AC32 | 19.82 | 1.11 | 14 | AC7 | 31.82 | 1.00 | 34 | AC46 | 51.45 | 0.71 | 54 |
| AC35 | 19.91 | 0.98 | 15 | AC23 | 32.18 | 1.00 | 35 | AC22 | 52.09 | 0.66 | 55 |
| AC2 | 20.18 | 1.17 | 16 | AC51 | 34.09 | 0.53 | 36 | AC44 | 54.82 | 0.58 | 56 |
| AC27 | 20.18 | 1.10 | 17 | AC59 | 34.18 | 0.90 | 37 | AC47 | 54.82 | 0.50 | 57 |
| AC5 | 20.55 | 1.04 | 18 | AC17 | 34.27 | 1.00 | 38 | AC52 | 56.64 | 0.47 | 58 |
| AC34 | 22.00 | 0.97 | 19 | AC43 | 34.82 | 0.77 | 39 | AC48 | 58.27 | 0.37 | 59 |
| AC40 | 22.18 | 1.09 | 20 | AC56 | 34.82 | 1.01 | 40 |
| Accession Code | ASR | STI | Rank | Accession Code | ASR | STI | Rank | Accession Code | ASR | STI | Rank |
|---|---|---|---|---|---|---|---|---|---|---|---|
| AC31 | 10.91 | 1.14 | 1 | AC13 | 21.36 | 1.07 | 21 | AC17 | 34.64 | 0.97 | 41 |
| AC38 | 11.00 | 1.11 | 2 | AC7 | 21.55 | 1.02 | 22 | AC22 | 35.00 | 0.74 | 42 |
| AC29 | 12.36 | 1.09 | 3 | AC34 | 22.73 | 0.93 | 23 | AC51 | 36.00 | 0.44 | 43 |
| AC37 | 12.73 | 1.02 | 4 | AC24 | 23.55 | 1.07 | 24 | AC57 | 36.27 | 0.33 | 44 |
| AC39 | 13.18 | 1.06 | 5 | AC41 | 25.18 | 0.99 | 25 | AC54 | 37.73 | 0.91 | 45 |
| AC30 | 13.36 | 1.14 | 6 | AC35 | 25.55 | 0.91 | 26 | AC56 | 40.91 | 0.89 | 46 |
| AC25 | 13.73 | 1.05 | 7 | AC33 | 25.73 | 0.95 | 27 | AC15 | 45.91 | 0.75 | 47 |
| AC2 | 14.45 | 1.17 | 8 | AC23 | 26.55 | 1.00 | 28 | AC18 | 46.73 | 0.68 | 48 |
| AC8 | 14.64 | 1.07 | 9 | AC21 | 26.64 | 1.05 | 29 | AC19 | 48.00 | 0.67 | 49 |
| AC32 | 14.91 | 1.10 | 10 | AC10 | 26.91 | 1.02 | 30 | AC55 | 48.55 | 0.68 | 50 |
| AC5 | 15.73 | 1.04 | 11 | AC14 | 28.91 | 0.96 | 31 | AC12 | 49.09 | 0.68 | 51 |
| AC16 | 16.00 | 1.09 | 12 | AC36 | 29.36 | 1.03 | 32 | AC58 | 49.64 | 0.63 | 52 |
| AC9 | 16.64 | 1.08 | 13 | AC43 | 30.45 | 0.78 | 33 | AC59 | 50.00 | 0.68 | 53 |
| AC11 | 17.36 | 0.99 | 14 | AC50 | 30.91 | 0.81 | 34 | AC47 | 53.18 | 0.46 | 54 |
| AC26 | 17.55 | 1.03 | 15 | AC4 | 31.36 | 1.00 | 35 | AC46 | 53.45 | 0.52 | 55 |
| AC27 | 19.73 | 1.08 | 16 | AC45 | 31.82 | 0.86 | 36 | AC44 | 54.45 | 0.48 | 56 |
| AC28 | 19.91 | 1.09 | 17 | AC42 | 32.91 | 0.96 | 37 | AC53 | 56.64 | 0.24 | 57 |
| AC3 | 20.00 | 0.88 | 18 | AC49 | 34.09 | 0.87 | 38 | AC52 | 57.00 | 0.33 | 58 |
| AC40 | 20.00 | 1.07 | 19 | AC20 | 34.18 | 0.84 | 39 | AC48 | 58.18 | 0.28 | 59 |
| AC6 | 20.27 | 0.98 | 20 | AC1 | 34.45 | 0.98 | 40 |
| Accession Code | ASR | STI | Rank | Accession Code | ASR | STI | Rank | Accession Code | ASR | STI | Rank |
|---|---|---|---|---|---|---|---|---|---|---|---|
| AC29 | 10.09 | 1.12 | 1 | AC6 | 21.91 | 0.99 | 21 | AC1 | 35.82 | 0.99 | 41 |
| AC38 | 10.09 | 1.14 | 2 | AC41 | 22.09 | 1.02 | 22 | AC57 | 36.27 | 0.39 | 42 |
| AC37 | 11.09 | 1.06 | 3 | AC11 | 22.45 | 0.99 | 23 | AC56 | 39.27 | 0.97 | 43 |
| AC24 | 13.27 | 1.12 | 4 | AC21 | 22.55 | 1.11 | 24 | AC49 | 39.82 | 0.87 | 44 |
| AC39 | 14.64 | 1.07 | 5 | AC32 | 22.55 | 1.10 | 25 | AC53 | 40.00 | 0.37 | 45 |
| AC30 | 15.36 | 1.16 | 6 | AC3 | 23.36 | 0.87 | 26 | AC22 | 42.36 | 0.74 | 46 |
| AC25 | 15.55 | 1.06 | 7 | AC5 | 24.00 | 1.03 | 27 | AC12 | 43.82 | 0.83 | 47 |
| AC28 | 16.18 | 1.13 | 8 | AC34 | 24.73 | 0.95 | 28 | AC15 | 45.45 | 0.83 | 48 |
| AC31 | 16.36 | 1.15 | 9 | AC7 | 25.45 | 1.03 | 29 | AC59 | 46.00 | 0.82 | 49 |
| AC36 | 16.45 | 1.11 | 10 | AC10 | 25.55 | 1.05 | 30 | AC18 | 46.09 | 0.75 | 50 |
| AC2 | 16.64 | 1.19 | 11 | AC50 | 28.09 | 0.86 | 31 | AC54 | 47.73 | 0.82 | 51 |
| AC16 | 16.73 | 1.11 | 12 | AC45 | 29.27 | 0.89 | 32 | AC55 | 47.91 | 0.77 | 52 |
| AC8 | 17.55 | 1.08 | 13 | AC14 | 29.82 | 0.97 | 33 | AC19 | 48.91 | 0.73 | 53 |
| AC9 | 17.73 | 1.10 | 14 | AC43 | 30.27 | 0.82 | 34 | AC58 | 52.09 | 0.66 | 54 |
| AC27 | 19.18 | 1.11 | 15 | AC33 | 31.91 | 0.93 | 35 | AC46 | 52.73 | 0.67 | 55 |
| AC40 | 19.64 | 1.10 | 16 | AC42 | 32.27 | 0.98 | 36 | AC47 | 54.27 | 0.51 | 56 |
| AC26 | 20.27 | 1.04 | 17 | AC20 | 32.82 | 0.86 | 37 | AC44 | 54.45 | 0.61 | 57 |
| AC23 | 20.64 | 1.04 | 18 | AC17 | 33.09 | 1.01 | 38 | AC52 | 56.64 | 0.46 | 58 |
| AC13 | 20.91 | 1.10 | 19 | AC4 | 34.00 | 1.01 | 39 | AC48 | 58.55 | 0.32 | 59 |
| AC35 | 21.55 | 0.96 | 20 | AC51 | 35.73 | 0.48 | 40 |
| Cd-125 | |||||
| Features | p-Value | Significant | High Tolerance | Moderate Tolerance | Low Tolerance |
| WU | 0.00 | Yes | 273.39 a ± 22.54 | 229.81 b ± 16.67 | 129.67 c ± 11.23 |
| LP | 0.50 | No | 11.61 a ± 2.89 | 10.25 a ± 3.78 | 9.42 a ± 2.64 |
| GPA | 0.50 | No | 0.25 a ± 0.01 | 0.22 a ± 0.01 | 0.18 a ± 0.01 |
| AC | 0.61 | No | 946.52 a ± 49.09 | 912.26 a ± 42.56 | 888.43 a ± 38.82 |
| CAT | 0.61 | No | 64.94 a ± 11.42 | 55.74 a ± 16.38 | 55.11 a ± 17.41 |
| SSC | 0.78 | No | 275.63 a ± 28.21 | 277.90 a ± 32.30 | 240.78 a ± 38.19 |
| TPC | 0.78 | No | 151.90 a ± 34.17 | 146.29 a ± 29.51 | 139.81 a ± 27.89 |
| PC | 0.85 | No | 1410.00 a ± 167.42 | 1325.00 a ± 122.45 | 1274.00 a ± 156.73 |
| Cd-250 | |||||
| Features | p-Value | Significant | High Tolerance | Moderate Tolerance | Low Tolerance |
| GPA | 0.00 | Yes | 0.38 a ± 0.005 | 0.20 b ± 0.002 | 0.12 c ± 0.006 |
| WU | 0.00 | Yes | 229.18 a ± 12.21 | 191.28 b ± 18.93 | 151.43 c ± 17.28 |
| CAT | 0.00 | Yes | 83.56 a ± 9.12 | 56.23 b ± 8.56 | 47.12 b ± 9.08 |
| TPC | 0.02 | Yes | 184.80 a ± 23.02 | 156.15 b ± 24.11 | 148.25 b ± 13.26 |
| SSC | 0.20 | No | 367.77 a ± 37.04 | 305.40 a ± 49.03 | 286.97 a ± 29.19 |
| AC | 0.24 | No | 950.15 a ± 56.11 | 921.59 a ± 42.22 | 856.87 a ± 36.12 |
| PC | 0.63 | No | 2530.00 a ± 112.08 | 2437.00 a ± 118.17 | 2022.00 a ± 119.23 |
| LP | 0.66 | No | 13.55 a ± 0.92 | 13.67 a ± 1.10 | 12.17 a ± 1.21 |
| Cd-500 | |||||
| Features | p-Value | Significant | High Tolerance | Moderate Tolerance | Low Tolerance |
| GPA | 0.00 | Yes | 0.44 a ± 0.003 | 0.21 b ± 0.001 | 0.11 c ± 0.001 |
| WU | 0.00 | Yes | 181.08 a ± 13.12 | 155.79 b ± 21.17 | 115.35 c ± 16.03 |
| PC | 0.00 | Yes | 3711.00 a ± 24.29 | 2130.00 b ± 19.83 | 1388.00 c ± 27.07 |
| TPC | 0.00 | Yes | 183.47 a ± 9.76 | 138.30 b ± 7.72 | 119.77 b ± 8.86 |
| SSC | 0.00 | Yes | 473.20 a ± 23.26 | 270.21 b ± 19.19 | 238.06 b ± 17.05 |
| AC | 0.00 | Yes | 975.98 a ± 16.77 | 843.01 b ± 17.03 | 795.59 b ± 11.52 |
| LP | 0.00 | Yes | 13.91 a ± 1.07 | 10.66 b ± 1.15 | 8.63 b ± 1.17 |
| CAT | 0.08 | No | 73.12 a ± 6.78 | 56.13 a ± 7.29 | 49.03 a ± 4.89 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tahir, N.A.-r.; Lateef, D.D.; Mustafa, K.M.; Rasul, K.S.; Khurshid, F.F. Determination of Physiochemical Characteristics Associated with Various Degrees of Cadmium Tolerance in Barley Accessions. Agronomy 2023, 13, 1502. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13061502
Tahir NA-r, Lateef DD, Mustafa KM, Rasul KS, Khurshid FF. Determination of Physiochemical Characteristics Associated with Various Degrees of Cadmium Tolerance in Barley Accessions. Agronomy. 2023; 13(6):1502. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13061502
Chicago/Turabian StyleTahir, Nawroz Abdul-razzak, Djshwar Dhahir Lateef, Kamil Mahmud Mustafa, Kamaran Salh Rasul, and Fawzy Faidhullah Khurshid. 2023. "Determination of Physiochemical Characteristics Associated with Various Degrees of Cadmium Tolerance in Barley Accessions" Agronomy 13, no. 6: 1502. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13061502