
Ecological Interaction in the Proliferation of Phytoparasitic Nematodes in Coffee var. Typica
 , , , , , , , , ,
, , , , , , , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Climatic Conditions
2.3. Soil Analysis
2.4. Soil and Root Sampling for Nematode Analysis
2.5. Soil and Root Nematode Analysis
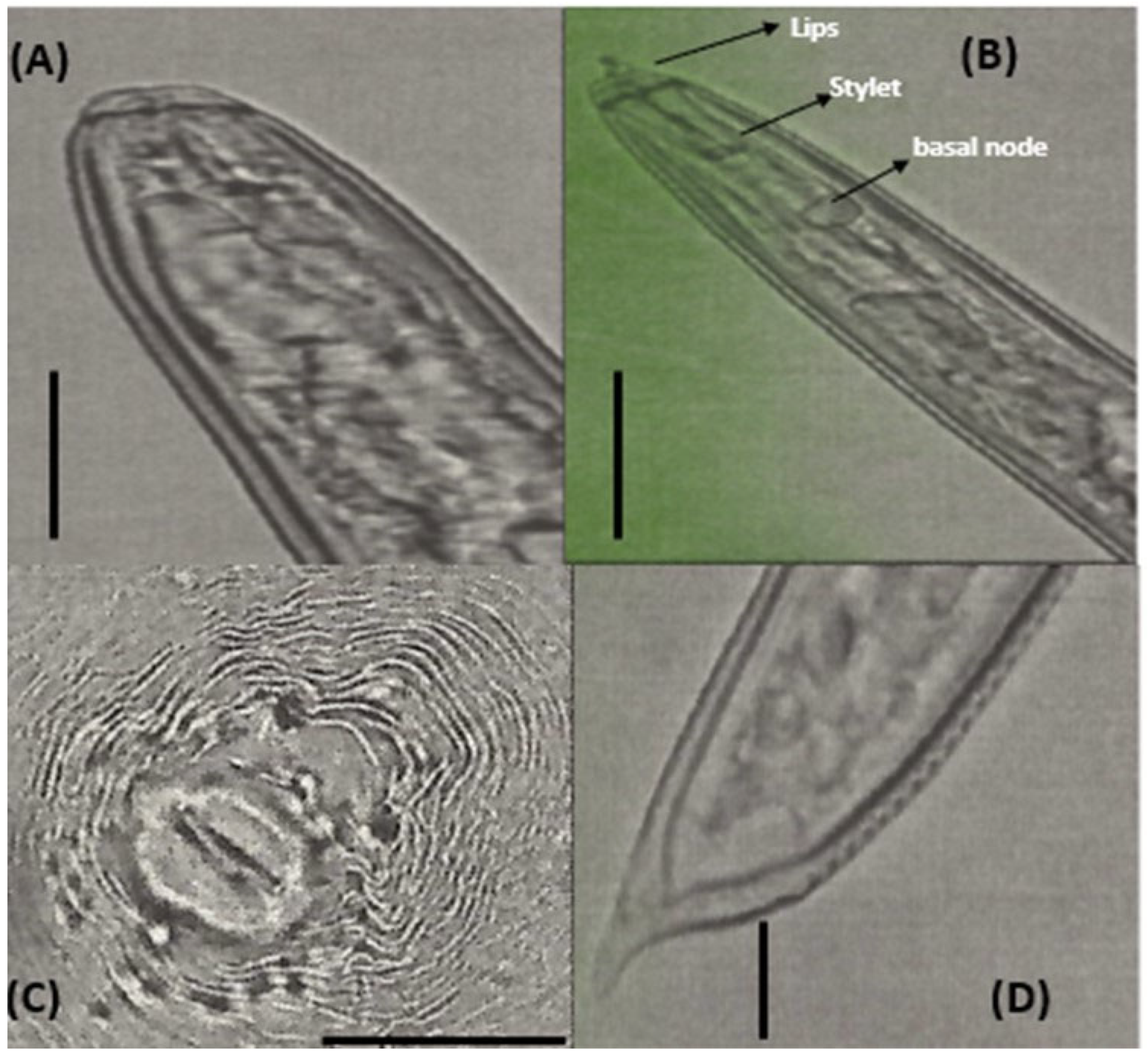
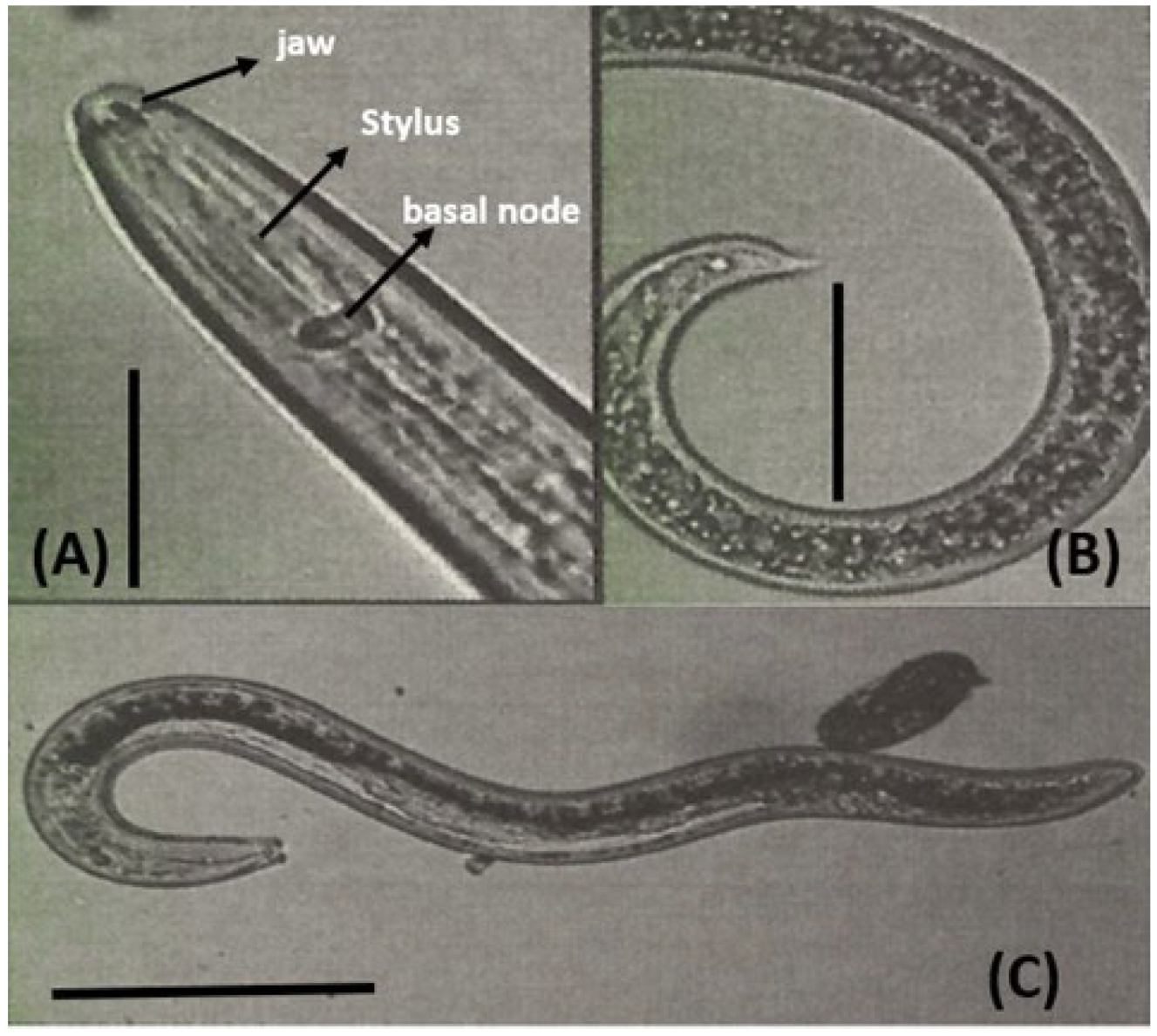
2.6. Identification of Phytoparasitic Nematodes
2.7. Statistical Analysis
3. Results
3.1. Characterization of the Environmental, Physical and Chemical Factors of the Collection Areas
3.2. Identification of Phytoparasitic Nematodes
3.3. Incidence of Phytoparasitic Nematodes in the Soil
3.4. Incidence of Phytoparasitic Nematodes on Roots
3.5. PCA Analysis
3.6. Pearson Correlation Analysis
3.7. Correlation Networks Established for the Different Nematodes Described
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Morales-Aranibar, L.; Yucra, F.E.Y.; Estrada, N.M.P.; Flores, P.Q.; Zevallos, R.N.M.; Zegarra, J.C.L.; Trujillo, U.P.; Aranibar, C.G.M.; Gonzales, H.H.S.; Aguilera, J.G.; et al. Production of New Biopesticides from Cymbopogon citratus for the Control of Coffee Rust (Hemileia vastatrix) under Laboratory and Field Conditions. Plants 2023, 12, 1166. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of The United Nations—FAO. Countries by Commodity. 2022. Available online: https://www.fao.org/faostat (accessed on 21 February 2021).
- Guevara-Sánchez, M.; Bernales del Águila, C.I.; Saavedra-Ramírez, J.; Owaki-López, J.J. Effect of altitude on coffee (Coffea arabica L.) quality: Comparison between mechanical and traditional drying. Sci. Agropecu. 2019, 10, 505–510. [Google Scholar] [CrossRef]
- MINAGRI. Síntesis Agroeconómica del Café; Dirección General de Políticas Agrarias: Lima, Peru, 2015. [Google Scholar]
- Mestanza, O.Q.; Oliva, M.L.; Obando, J.J. Hemileia vastatrix in coffee trees of Leoncio Prado and Huánuco provinces in the Huánuco region. Peru Coffee Cocoa Res. J. 2015, 3, 44–50. [Google Scholar]
- Bustillo, A.E. Nematodes and Coffee Diseases; ICAFE: San José, Costa Rica, 2003. [Google Scholar]
- Garambel-Acurio, S.I.; Lima-Medina, I.; Loza-Del Carpio, A.; Checahuari Jarata, S.E. Incidence of nematodes associated with the main coffee producing areas of the Puno region, Peru. Bioagro 2022, 34, 85–96. [Google Scholar] [CrossRef]
- Talavera, M. Guía de Control de Enfermedades y Plagas del Cafeto; INIAP: Lima, Peru, 2004. [Google Scholar]
- Coyne, P.; Nicol, J.; Claudius-Cole, B. Practical Plant Nematology: Field and Laboratory Guide. Secretariat; International Institute of Tropical Agriculture: Cotonou, Benin, 2008. [Google Scholar]
- Castillo, P.; Vovlas, N. Plant-parasitic nematode management in coffee: The role of microbiome engineering. Plant Soil 2017, 410, 401–418. [Google Scholar] [CrossRef]
- Inserra, R.N. Nematodes associated with coffee in Puerto Rico; Association for Plant Protection of Puerto Rico: San Juan, Puerto Rico, 1978; Volume 5, pp. 72–78. [Google Scholar]
- Sikora, R.A. Plant parasitic nematodes: A challenging pest to the coffee industry. J. Nematol. 2009, 41, 124–129. [Google Scholar]
- Ploeg, A.T.; Rodriguez, J. Plant-Parasitic Nematodes in Coffee. In Coffee Pests, Diseases and their Management; Zentmyer, J.A., Ed.; CAB International: Wallingford, UK, 2013; pp. 359–379. [Google Scholar]
- Decraemer, W.; Hunt, D.J. Structure and classification. In Plant Nematology; Perry, R.N., Moens, M., Eds.; CABI: Wallingford, UK, 2006; pp. 3–32. [Google Scholar] [CrossRef]
- Ferris, H.; Matute, M. Structural and functional succession in the nematode fauna of a soil food web. Appl. Soil Ecol. 2003, 23, 93–110. [Google Scholar] [CrossRef]
- Singh, R.; Yadav, D.K. Management of Plant-Parasitic Nematodes in Coffee: A Review. J. Plant Prot. Res. 2018, 58, 221–233. [Google Scholar]
- Yeates, G.W.; Bongers, T.; de Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding habits in soil nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Dababat, A.A.; Sikora, R.A.; Coyne, D. Nematode parasites of vegetables. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture; CABI: Wallingford, UK, 2010; pp. 319–392. [Google Scholar]
- Zasada, I.A.; Ferris, H.; Faghihi, J.; Hooks, C.R. Nematode communities as indicators of status and processes of soils. In Nematology in the 21st Century; Brill: Leiden, The Netherlands, 2010; pp. 409–436. [Google Scholar]
- Sikora, R.A.; Fernandez, E. Nematode parasites of coffee. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture; CABI: Wallingford, UK, 2005; pp. 459–485. [Google Scholar]
- Jaffee, B.A.; Pasternak, D.; Zilberman, D. Economic analysis of pest management strategies: The impact of nematodes on California strawberries. Crop Prot. 2014, 58, 33–39. [Google Scholar]
- Perry, R.N. Effect of population density of plant parasitic nematodes on plant growth. Nematology 2002, 4, 717–720. [Google Scholar]
- Vovlas, N.; Castillo, P. Pratylenchus in coffee: A review. Nematology 2011, 13, 547–560. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Rhoades, J.D. Soluble salts. In Methods of Soil Analysis: Part 2 Chemical and Microbiological Properties; ASA and SSSA: San Antonio, TX, USA, 1982; pp. 167–179. [Google Scholar]
- Canto, M. Manual de Nematología; Universidad Nacional Agraria La Molina: Lima, Perú, 2003. [Google Scholar]
- Durán Mora, J. Diagnosis, Population Dynamics, Biochemical and Molecular Characterization of Nodulating Nematodes (Meloidogyne spp.) in Tropical Crops in Costa Rica; Center for Research and Development of Sustainable Agriculture in the Humid Tropics (CIDASTH), Technological Institute of Costa Rica: Cartago, Costa Rica, 2012. [Google Scholar]
- Oliveira, C.; Santos, M.; Silva, L.H. Diagnosis of Phytomatodes, 1st ed.; Editorial Millenium: Sofia, Bulgaria, 2016. [Google Scholar]
- Rashid, F.; De Waele, D.; Coomans, A. Trichodoridae (Nematoda) of Brazil. Nematologica 1985, 31, 289–320. [Google Scholar]
- Mancini, G.; Moretti, F. The genus Helicotylenchus in Piedmont and in the Aosta Valley. Pt. 1. Redia 1976, 59, 225–228. [Google Scholar]
- Handoo, Z.A.; Golden, A.M. A key and diagnostic compendium to the species of the genus Trichodorus Cobb, 1913 (Nematoda: Trichodoridae). J. Nematol. 1990, 22, 176–194. [Google Scholar]
- Coomans, A. The taxonomy of the Dorylaimida (Nematoda) revisited. Fundam. Appl. Nematol. 1996, 19, 111–124. [Google Scholar]
- Giblin-Davis, R.M.; Williams, D.S.; Bekal, S. Rhabditida: Biology and systematics. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture; CABI Publishing: Wallingford, UK, 2001; pp. 55–96. [Google Scholar]
- Loof, P.A.A.; Coomans, A. The taxonomy of the nematode family Mononchidae. Wagening. Agric. Univ. Pap. 1989, 89, 1–120. [Google Scholar]
- Taylor, D.P.; Netscher, C. A taxonomic study of the genus Criconemella De Grisse & Loof, 1965 (Criconematidae: Tylenchida) with a discussion on the validity of certain species. Nematologica 1974, 20, 449–468. [Google Scholar]
- Bhering, L.L. Rbio: A Tool for Biometric and Statistical Analysis Using the R Platform. Crop. Breed. Appl. Biotechnol. 2017, 17, 187–190. [Google Scholar] [CrossRef] [Green Version]
- Machado, A.C.Z.; Kumar, R.; dos Reis Fatobene, B.J. Nematode problems in coffee and their sustainable management. In Nematode Diseases of Crops and Their Sustainable Management; Academic Press: Cambridge, MA, USA, 2023; pp. 641–651. [Google Scholar]
- Pereira, A.A.A.; Paiva, P.E.B.; Santos, C.M. Meloidogyne spp. in Coffea arabica L. cv. Red Catuaí IAC 144 in the state of Paraná, Brazil. Trop. Plant Pathol. 2019, 44, 331–333. [Google Scholar]
- Vieira, A.R.; Coelho, L.H.; Costa, L.M.; de Oliveira, A.F.; Campos, V.P. Occurrence and population density of root-knot nematodes in coffee fields in Brazil. Australas. Plant Pathol. 2015, 44, 257–264. [Google Scholar]
- Kushwaha, R.K.; Singh, A.K.; Rai, A.K.; Yadav, R.K. Diversity and prevalence of root-knot nematodes associated with coffee in India. J. Environ. Biol. 2020, 41, 609–616. [Google Scholar]
- Rajendran, G.; Ojiambo, P.S. Management of root-knot nematodes in coffee: A review of current research and future prospects. Plant Dis. 2021, 105, 1506–1519. [Google Scholar]
- Ribeiro, L.P.; Carneiro, R.G.; Guimarães, A.M.; Santos, J.M.; Gomes, A.C.M.M.M. Detection and molecular identification of Meloidogyne spp. in coffee fields in the state of Bahia, Brazil. Australas. Plant Dis. Notes 2019, 14, 29. [Google Scholar]
- Molina-Bravo, R.; Hernández-Uribe, J.P.; del Prado-Martínez, A.M. Response of Coffea arabica L. cv. Colombia to Meloidogyne incognita inoculation under shade and full sun conditions. Cienc. E Investig. Agrar. 2019, 46, 219–227. [Google Scholar]
- Gutiérrez-Gutiérrez, C.; Escuer, M.; Verdejo-Lucas, S. A review of the Dorylaimida (Nematoda) from Spain with description of six new species. Zootaxa 2020, 4729, 1–42. [Google Scholar]
- Li, X.; Huang, X.; Chen, X. Effects of soil physicochemical properties and microbial communities on the occurrence of plant-parasitic nematodes in vegetable fields. J. Nematol. 2019, 51, 1–10. [Google Scholar]
- Peng, H.; Chen, S.; Yang, J.; Li, L.; Liang, W. Relationships between plant-parasitic nematodes, soil properties and plant growth in Chinese ginger fields. J. Nematol. 2018, 50, 333–342. [Google Scholar]
- Rondanelli, G.M.; Longa, C.M.O.; Chitarra, W.S.; Rodrigues, F.A. Soil microbial biomass and enzymatic activity associated with coffee plants with different resistance to nematodes. Sci. Agric. 2020, 77, e20180031. [Google Scholar]
- Bernardo, J.T.; Aguilera, J.G.; Silva, R.B.; Medeiros, I.R.E.; Vian, R.; Niella, G.R.; Ulhoa, C.J. Isolamento on farm de Trichoderma: Uma ferramenta no controle de doenças de solo para os agricultores no Brasil. Revista Eletrônica Científica da UERGS 2019, 5, 263–270. [Google Scholar] [CrossRef]
- Aguilera, J.G.; Santos, J.C.S.S.; Santos, C.S.O.; Zuffo, A.M.; Vian, R.; Medeiros, I.R.E.; Barbosa, A.; Silva, R.B.; Bernardo, J.T. Otimização do método de produção massal de Trichoderma asperellum (Ascomycota: Hypocreaceae) em arroz. Acta Biológica Catarinense 2021, 7, 46–58. [Google Scholar] [CrossRef]
- Cruvinell, A.; Silva, T.M.; Rocha, M.A.M.; Azevedo, M.O.; Aguilera, J.G.; Bernardo, M.L.T.; Bernardo, J.T.; Almeida, J.E.M. Profitability of soybean production on Bom Jardim Lagoano farm with on-farm biological management. Res. Soc. Dev. 2022, 11, e135111436112. [Google Scholar] [CrossRef]
- Zhu, Q.; Zhao, L.; Li, X.; Li, F.; Li, X. Effects of different cropping patterns on soil nematode community structure in the Loess Plateau, China. J. Nematol. 2021, 53, 1–11. [Google Scholar]
- Kushwaha, S.; Choudhary, D.K.; Varma, A. Plant parasitic nematodes and their management in sustainable agriculture. In Sustainable Agriculture Reviews; Springer: Cham, Switzerland, 2020; pp. 205–233. [Google Scholar]
- Pereira, J.C.; Fernandes, C.D.; Martins, F.A.; Carneiro, R.M.D.G. Distribution of Meloidogyne spp. in coffee fields in Minas Gerais, Brazil. J. Nematol. 2019, 51, e2019-01. [Google Scholar]
- Vieira, P.; Powers, T.; Neves, M. Root-knot nematodes associated with coffee (Coffea arabica L.) in Brazil. Nematropica 2015, 4, 171–180. [Google Scholar]
- Rajendran, G.; Ojiambo, P.S. Phytoparasitic nematodes of coffee: A review of distribution, biology, and management. Ann. Appl. Biol. 2021, 179, 9–25. [Google Scholar]
- Ribeiro, S.G.; De Almeida, M.R.A.; Da Silva, M.C.M.; Correa, V.R. Nematode community in coffee plantations in Brazil: A comprehensive survey. J. Nematol. 2019, 51, e2019-08. [Google Scholar]
- Zhu, Y.; Xue, S.; Wang, X.; Li, Y.; Zhang, X.; Li, D. Nematode infection alters the phytohormone and amino acid metabolisms in coffee roots. J. Agric. Food Chem. 2021, 69, 3101–3109. [Google Scholar]
- Gutiérrez-Gutiérrez, C.; Ramírez-Suero, M.; Sánchez-Brito, C. Evaluation of fungi and nematodes in soils of coffee and orange crops in Mexico. Agrociencia 2020, 54, 557–570. [Google Scholar]
- Molina-Bravo, R.; Martínez-González, C.; García-Sánchez, E. Dorylaimida and Triplonchida nematodes from natural and agricultural soils in Mexico, with descriptions of four new species. J. Nematol. 2019, 51, e2019-10. [Google Scholar]
- Ploeg, A.T.; Rodriguez, M. Nematodes and their management in sustainable coffee production: A review. J. Nematol. 2013, 45, 1–13. [Google Scholar]
- Rivas, S.D.D.; Díaz, D.D.; Ávila, J.J. Effect of organic matter and tillage on soil nematode population. Agroproductividad 2017, 10, 69–74. [Google Scholar]
- Smith, J.; Johnson, A.; Brown, L. Effects of soil texture on plant-parasitic nematode populations. J. Agric. Sci. 2017, 145, 231–245. [Google Scholar]
- Jones, S.M.; Smith, R.L.; Johnson, K.W. Influence of soil texture on water and nutrient retention and its implications for plant-parasitic nematode infestation. Soil Sci. Soc. Am. J. 2014, 78, 456–467. [Google Scholar]
- Alfaro-Lucas, J.M.; Escuer, M.; Sorribas, F.J. Altitudinal variation in soil nematode communities in Mediterranean high mountain soils. J. Mt. Sci. 2018, 15, 35–46. [Google Scholar]
- Molinari, H.B.C.; Oliveira, C.M.G.; Ferraz, L.C.C.C.B.; Faria, M.V. Soil nematode community in Coffea arabica plantations with different plant densities and altitudes in Brazil. Nematology 2007, 9, 353–362. [Google Scholar] [CrossRef]
- Bolan, N.S.; Kunhikrishnan, A.; Thangarajan, R.; Kumpiene, J.; Park, J.; Makino, T.; Kirkham, M.B.; Scheckel, K. Remediation of heavy metal(loid)s contaminated soils - To mobilize or to immobilize? J. Hazard. Mater. 2011, 197, 443–460. [Google Scholar] [CrossRef]
- Schimel, J.; Bennett, J. Nitrogen mineralization: Challenges of a changing paradigm. Ecology 2004, 85, 591–602. [Google Scholar] [CrossRef]
- Ferraz, L.C.; Ferraz, S.; Flegg, J.J.M.; Molla, O.A.S. Distribution of root-knot nematodes (Meloidogyne spp.) and associated soil physical and chemical properties in banana fields in Brazil. Appl. Soil Ecol. 2008, 38, 90–99. [Google Scholar] [CrossRef]
- Ferris, H.; Zheng, L.; Walker, M.A. Density-dependent responses of the grape root nematode, Xiphinema index, to wine grape root systems. J. Nematol. 2012, 44, 36–43. [Google Scholar]
- Neher, D.A. Ecology of plant and free-living nematodes in natural and agricultural soil. Annu. Rev. Phytopathol. 2010, 48, 371–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bebber, D.P.; Ramotowski, M.A.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Melakeberhan, H.; Hobbs, S.L.A.; Smith, D.L. Effects of Meloidogyne incognita and Pratylenchus coffeae on the growth of coffee. J. Nematol. 2001, 33, 53–60. [Google Scholar]
- Bridge, J.; Starr, J. Nematode parasites of coffee and their management. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture; Waller, J.M., Briscoe, M.B.R., Evans, H.C., Eds.; CABI Publishing: Wallingford, UK, 2007; pp. 423–458. [Google Scholar]
- Eskes, A.B.; Engels, J.M.M. Nematode parasites of coffee, cacao, and tea. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture, 3rd ed.; Luc, M., Sikora, R.A., Bridge, J., Eds.; CABI Publishing: Wallingford, UK, 2017; pp. 435–474. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytonematodes | Identification of Key Phytonematodes |
|---|---|
| Meloidogyne spp. | Durán Mora [27] |
| Pratylenchus spp. | Oliveira, Santos and Silva [28] |
| Allotrichodorus spp. | Rashid, De Waele and Coomans [29] |
| Helicotylenchus spp. | Mancini & Moretti [30] |
| Trichodorus spp. | Handoo and Golden [31] |
| Dorylaimidos spp. | Coomans [32] |
| Rhabditidos spp. | Giblin-Davis, Williams and Bekal [33] |
| Mononchids spp. | Loof and Coomans [34] |
| Criconematidos spp. | Taylor and Netscher [35] |
| Region | Altitude (masl) | T * (°C) | Humidity (%) | Soil Texture | pH (KCl) | SOM (%) |
|---|---|---|---|---|---|---|
| Quebrada Honda | 1755 | 23.4 | 67 | Sandy loam | 5.15 | 4.12 |
| Huayanay High 01 | 1866 | 21.3 | 84.6 | Sandy loam | 5.47 | 7.53 |
| Stockade | 1676 | 25.8 | 80.9 | Sandy clay loam | 5.32 | 4.89 |
| Huayanay Center 01 | 1579 | 28.1 | 84.6 | Sandy clay loam | 5.18 | 3.22 |
| Huayanay | 1515 | 27.4 | 78.5 | Sandy clay loam | 5.25 | 3.35 |
| Huayanay High 02 | 1632 | 22.5 | 82.7 | Sandy loam | 5.51 | 6.81 |
| Aguilayoc | 1400 | 24.2 | 85.4 | Sandy clay loam | 5.42 | 6.11 |
| Huayanay Center 02 | 1642 | 28.3 | 84.4 | Sandy loam | 5.22 | 3.33 |
| Genre | Huayanay High 1 | Huayanay High 2 | Huayanay Center 1 | Huayanay Center 2 | Huayanay | Quebrada Honda | Aguilayoc | Stockade | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | % | No. | % | No. | % | No. | % | No. | % | No. | % | No. | % | No. | % | |
| Pratylenchus spp. | 20 | 11.1 | - | - | - | - | - | - | - | - | - | - | 10 | 2.3 | - | - |
| Trichodorides spp. | 40 | 22.3 | 20 | 5 | 20 | 7.6 | - | - | 30 | 11.1 | - | - | - | - | - | - |
| Dorylaimidae spp. | 100 | 55.5 | 140 | 35 | 100 | 38.4 | - | - | 60 | 22.2 | 80 | 30.8 | 80 | 18.2 | 60 | 42.8 |
| Rhabditidae spp. | 20 | 11.1 | 20 | 5 | - | - | 20 | 7.4 | 40 | 14.8 | - | - | - | - | 20 | 14.4 |
| Meloidogyne spp. | - | - | 160 | 40 | 120 | 46.4 | 200 | 74 | 120 | 44.4 | 120 | 46.2 | 340 | 77.3 | 60 | 42.8 |
| Mononchids spp. | - | - | 60 | 15 | - | - | 10 | 3.7 | - | - | - | - | - | - | - | - |
| Crichonematides spp. | - | - | - | - | 20 | 7.6 | - | - | - | - | 20 | 7.6 | - | - | - | - |
| Helicotylenchus spp. | - | - | - | - | - | - | 40 | 14.9 | 20 | 7.5 | 40 | 15.4 | 10 | 2.2 | - | - |
| TOTAL | 180 | 100 | 400 | 100 | 260 | 100 | 270 | 100 | 270 | 100 | 260 | 100 | 440 | 100 | 140 | 100 |
| Genre | Huayanay High 1 | Huayanay High 2 | Huayanay Center 1 | Huayanay Center 2 | Huayanay | Quebrada Honda | Aguilayoc | Stockade | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | % | No. | % | No. | % | No. | % | No. | % | No. | % | No. | % | No. | % | |
| Pratylenchus spp. | 24 | 34.3 | 14 | 5.9 | - | - | 20 | 15.6 | - | - | - | - | 2 | 0.9 | 8 | 3.9 |
| Rhabditidae spp. | 34 | 48.6 | 36 | 15.3 | 12 | 13.1 | 38 | 29.6 | 22 | 5.6 | - | - | - | - | 26 | 12.9 |
| Meloidogyne spp. | 12 | 17.1 | 186 | 78.8 | 80 | 86.9 | 70 | 54.8 | 368 | 94.4 | 96 | 96 | 230 | 99.1 | 168 | 83.2 |
| Helicotylenchus spp. | - | - | - | - | - | - | - | - | - | - | 4 | 4 | - | - | - | - |
| TOTAL | 70 | 100 | 236 | 100 | 92 | 100 | 128 | 100 | 390 | 100 | 100 | 100 | 232 | 100 | 202 | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morales-Aranibar, L.; Yucra, F.E.Y.; Estrada, N.M.P.; Salcedo, E.P.; Morales-Aranibar, C.; Gutiérrez, N.L.; Aguilar, O.M.; Chura, R.M.M.; Contreras, W.C.; Aguilera, J.G.; et al. Ecological Interaction in the Proliferation of Phytoparasitic Nematodes in Coffee var. Typica. Agronomy 2023, 13, 1653. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13061653
Morales-Aranibar L, Yucra FEY, Estrada NMP, Salcedo EP, Morales-Aranibar C, Gutiérrez NL, Aguilar OM, Chura RMM, Contreras WC, Aguilera JG, et al. Ecological Interaction in the Proliferation of Phytoparasitic Nematodes in Coffee var. Typica. Agronomy. 2023; 13(6):1653. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13061653
Chicago/Turabian StyleMorales-Aranibar, Luis, Francisca Elena Yucra Yucra, Nivia Marisol Pilares Estrada, Eliseo Pumacallahui Salcedo, Carlos Morales-Aranibar, Nataniel Linares Gutiérrez, Oscar Mamani Aguilar, Richar Marlon Mollinedo Chura, Wilberth Caviedes Contreras, Jorge González Aguilera, and et al. 2023. "Ecological Interaction in the Proliferation of Phytoparasitic Nematodes in Coffee var. Typica" Agronomy 13, no. 6: 1653. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13061653