
Genome-Wide Association Study for Fusarium Head Blight Resistance in Common Wheat from China

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Index of FHB Response
- 1 = less than 0.25;
- 2 = 0.25–less than 0.5;
- 3 = 0.5–less than 0.75;
- 4 = 0.75–1 or entire spike dead.
2.3. Genotyping and Filtering
2.4. Genome-Wide Association Study
2.5. Haplotype Analysis
2.6. Candidate Gene Identification
3. Results
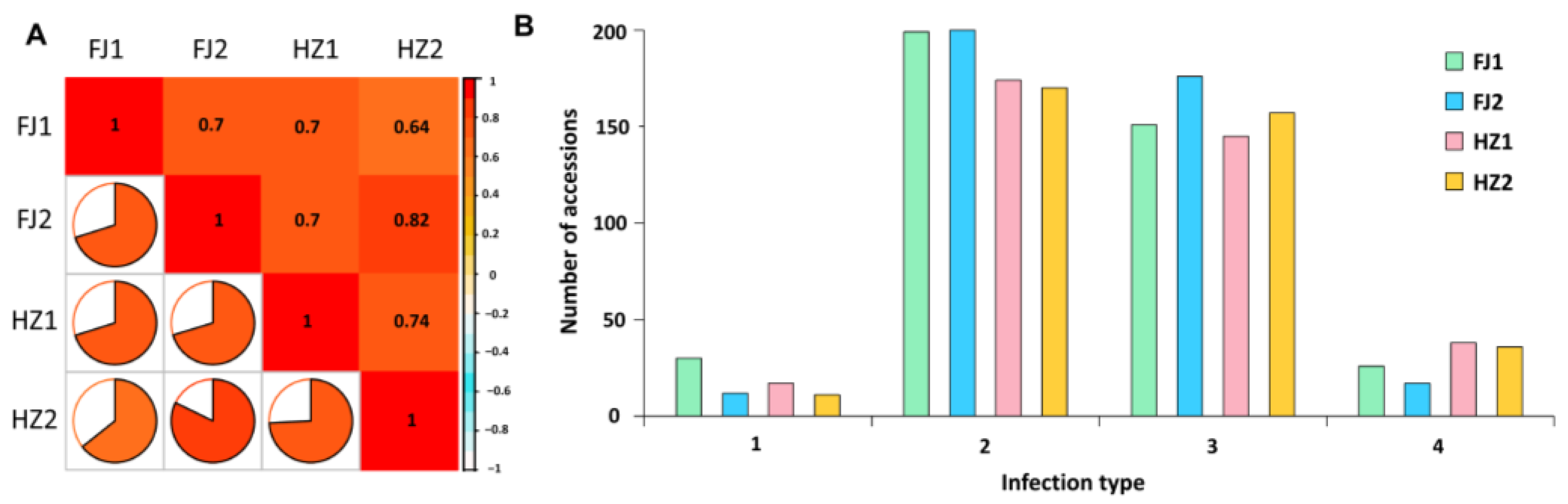
3.1. Distribution of FHB Responses in the Variety Panels
3.2. Genome-Wide Association Study of Wheat FHB Resistance
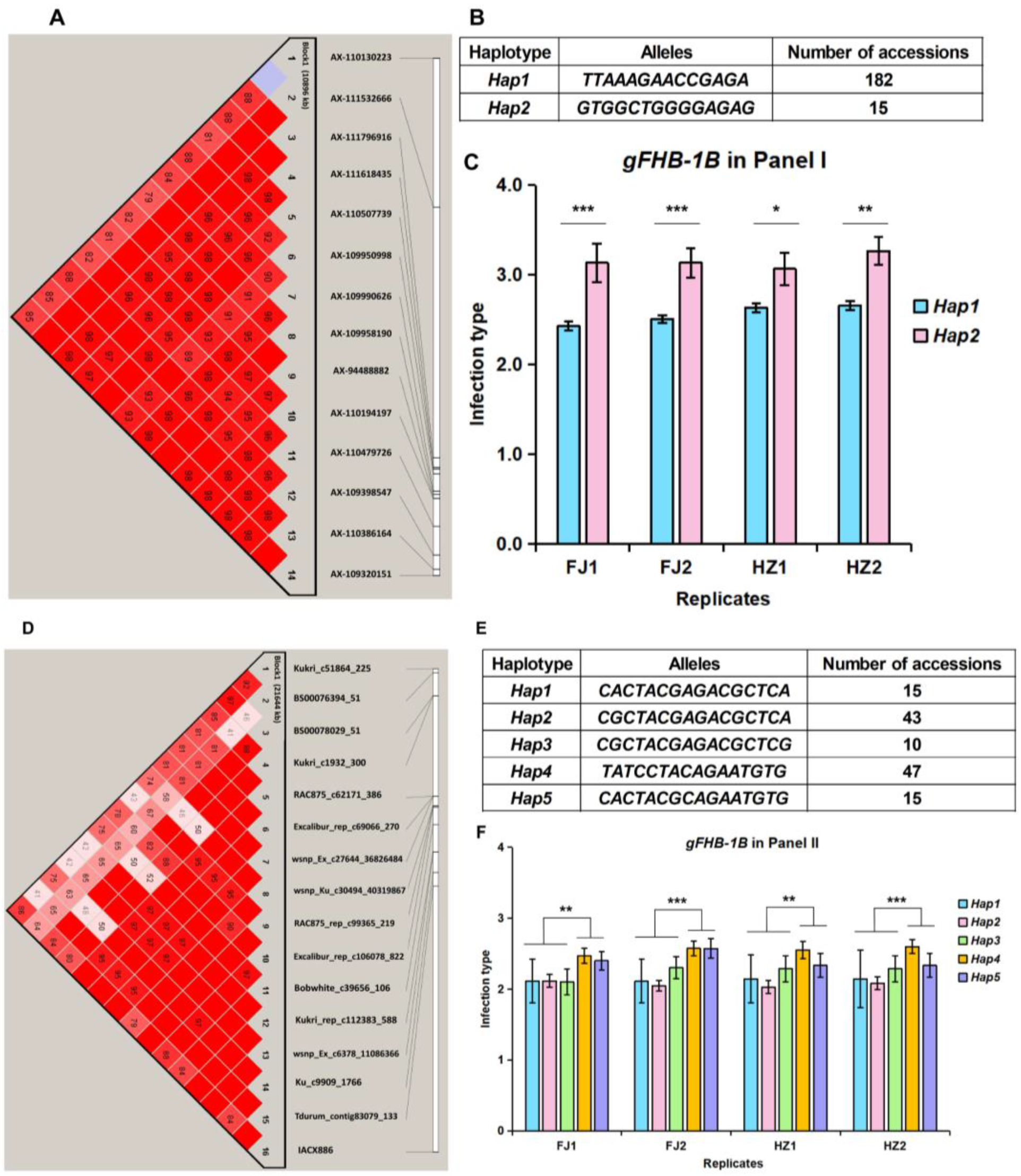
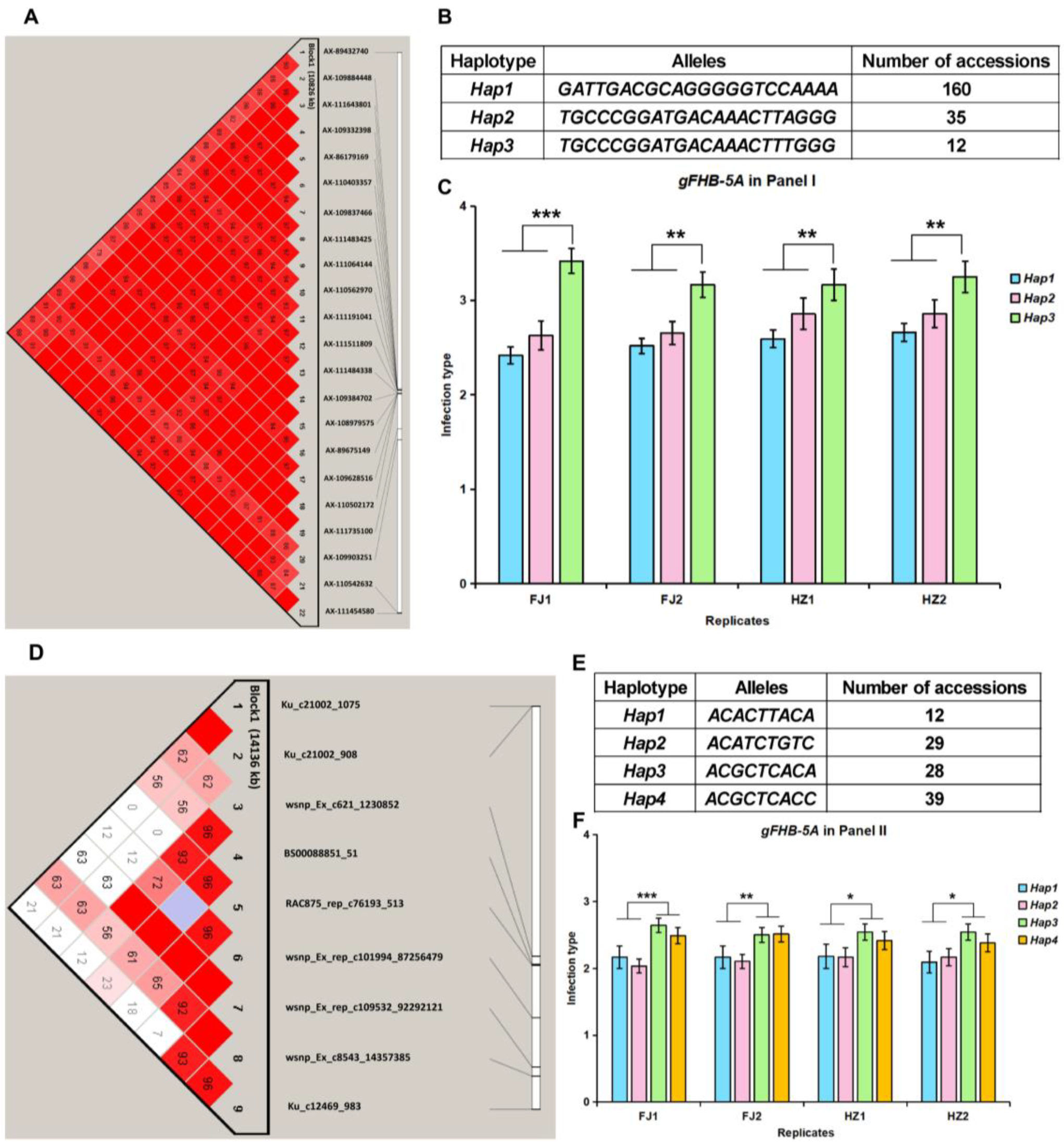
3.3. Haplotype Analysis of High-Confidence FHB Resistance Loci gFHB-1B and gFHB-5A
3.4. Prediction of Candidate Genes
3.5. Distribution of Resistance Alleles for gFHB-1B and gFHB-5A in the Huang-Huai Valley
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interests
References
- McMullen, M.; Bergstrom, G.; De-Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A unified effort to fight an enemy of wheat and barley: Fusarium head blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, S.H.; Zhang, Y.; Bie, T.D.; Gao, D.R. Damage of wheat Fusarium head blight epidemics and genetic improvement of wheat for scab resistance in China. Jiangsu J. Agric. Sci. 2012, 28, 938–942. [Google Scholar]
- Brown, N.A.; Urban, M.; van de Meene, A.M.; Hammond-Kosack, K.E. The infection biology of Fusarium graminearum: Defining the pathways of spikelet to spikelet colonisation in wheat ears. Fungal Biol. 2010, 114, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, B.; Mergoum, M.; Martinez-Espinoza, A.D.; Sapkota, S.; Pradhan, S.; Babar, M.A.; Bai, G.; Dong, Y.; Buck, J.W. Genetics of Fusarium head blight resistance in soft red winter wheat using a genome-wide association study. Plant Genome 2022, 15, e20222. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Hao, Y.; Mergoum, M.; Bai, G.; Humphreys, G.; Cloutier, S.; Xia, X.; He, Z. Breeding wheat for resistance to Fusarium head blight in the Global North: China, USA, and Canada. Crop J. 2019, 7, 730–738. [Google Scholar] [CrossRef]
- Buerstmayr, H.; Lemmens, M.; Hartl, L.; Doldi, L.; Steiner, B.; Stierschneider, M.; Ruckenbauer, P. Molecular mapping of QTLs for Fusarium head blight resistance in spring wheat. I. Resistance to fungal spread (Type II resistance). Theor. Appl. Genet. 2002, 104, 84–91. [Google Scholar] [CrossRef]
- Buerstmayr, H.; Steiner, B.; Hartl, L.; Griesser, M.; Angerer, N.; Lengauer, D.; Miedaner, T.; Schneider, B.; Lemmens, M. Molecular mapping of QTLs for Fusarium head blight resistance in spring wheat. II. Resistance to fungal penetration and spread. Theor. Appl. Genet. 2003, 107, 503–508. [Google Scholar] [CrossRef]
- Steiner, B.; Buerstmayr, M.; Michel, S.; Schweiger, W.; Lemmens, M.; Buerstmayr, H. Breeding strategies and advances in line selection for Fusarium head blight resistance in wheat. Trop. Plant Pathol. 2017, 42, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Cuthbert, P.A.; Somers, D.J.; Thomas, J.; Cloutier, S.; Brulé-Babel, A. Fine mapping Fhb1, a major gene controlling Fusarium head blight resistance in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2006, 112, 1465–1702. [Google Scholar] [CrossRef]
- Cuthbert, P.A.; Somers, D.J.; Brûlé-Babel, A. Mapping of Fhb2 on chromosome 6BS: A gene controlling Fusarium head blight field resistance in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 114, 429–437. [Google Scholar] [CrossRef]
- Xue, S.; Li, G.; Jia, H.; Xu, F.; Lin, F.; Tang, M.; Wang, Y.; An, X.; Xu, H.; Zhang, L.; et al. Fine mapping Fhb4, a major QTL conditioning resistance to Fusarium infection in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2010, 121, 147–156. [Google Scholar] [CrossRef]
- Xue, S.; Xu, F.; Tang, M.; Zhou, Y.; Li, G.; An, X.; Lin, F.; Xu, H.; Jia, H.; Zhang, L.; et al. Precise mapping Fhb5, a major QTL conditioning resistance to Fusarium infection in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 2011, 123, 1055–1063. [Google Scholar] [CrossRef]
- Cainong, J.C.; Bockus, W.W.; Feng, Y.; Chen, P.; Qi, L.; Sehgal, S.K.; Danilova, T.V.; Koo, D.H.; Friebe, B.; Gill, B.S. Chromosome engineering, mapping, and transferring of resistance to Fusarium head blight disease from Elymus tsukushiensis into wheat. Theor. Appl. Genet. 2015, 128, 1019–1027. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, X.; Hou, Y.; Cai, J.; Shen, X.; Zhou, T.; Xu, H.; Ohm, H.W.; Wang, H.; Li, A.; et al. High-density mapping of the major FHB resistance gene Fhb7 derived from Thinopyrum ponticum and its pyramiding with Fhb1 by marker-assisted selection. Theor. Appl. Genet. 2015, 128, 2301–2316. [Google Scholar] [CrossRef]
- Qi, L.; Pumphrey, M.; Friebe, B.; Chen, P.; Gill, B. Molecular cytogenetic characterization of alien introgressions with gene Fhb3 for resistance to Fusarium head blight disease of wheat. Theor. Appl. Genet. 2008, 117, 1155–1166. [Google Scholar] [CrossRef]
- Anderson, J.A.; Stack, R.W.; Liu, S.; Waldron, B.L.; Fjeld, A.D.; Coyne, C.; Moreno-Sevilla, B.; Fetch, J.M.; Song, Q.J.; Cregan, P.B.; et al. DNA markers for Fusarium head blight resistance QTLs in two wheat populations. Theor. Appl. Genet. 2001, 102, 1164–1168. [Google Scholar] [CrossRef]
- Bai, G.; Kolb, F.L.; Shaner, G.; Domier, L.L. Amplified fragment length polymorphism markers linked to a major quantitative trait locus controlling scab resistance in wheat. Phytopathology 1999, 89, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Lv, C.; Song, Y.; Gao, L.; Yao, Q.; Zhou, R.; Xu, R.; Jia, J. Integration of QTL detection and marker assisted selection for improving resistance to Fusarium head blight and important agronomic traits in wheat. Crop J. 2014, 2, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Schweiger, W.; Steiner, B.; Vautrin, S.; Nussbaumer, T.; Siegwart, G.; Zamini, M.; Jungreithmeier, F.; Gratl, V.; Lemmens, M.; Mayer, K.F.; et al. Suppressed recombination and unique candidate genes in the divergent haplotype encoding Fhb1, a major Fusarium head blight resistance locus in wheat. Theor. Appl. Genet. 2016, 129, 1607–1623. [Google Scholar] [CrossRef] [Green Version]
- Rawat, N.; Pumphrey, M.O.; Liu, S.; Zhang, X.; Tiwari, V.K.; Ando, K.; Trick, H.N.; Bockus, W.W.; Akhunov, E.; Anderson, J.A.; et al. Wheat fhb1 encodes a chimeric lectin with agglutinin domains and a pore-forming toxin-like domain conferring resistance to Fusarium head blight. Nat. Genet. 2016, 48, 1576–1580. [Google Scholar] [CrossRef]
- Li, G.; Zhou, J.; Jia, H.; Gao, Z.; Fan, M.; Luo, Y.; Zhao, P.; Xue, S.; Li, N.; Yuan, Y.; et al. Mutation of a histidine-rich calcium-binding-protein gene in wheat confers resistance to Fusarium head blight. Nat. Genet. 2019, 51, 1106–1112. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Bernardo, A.; Tian, B.; Chen, H.; Wang, S.; Ma, H.; Cai, S.; Liu, D.; Zhang, D.; Li, T.; et al. A deletion mutation in TaHRC confers Fhb1 resistance to Fusarium head blight in wheat. Nat. Genet. 2019, 51, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, S.; Ge, W.; Zhao, L.; Hou, B.; Wang, K.; Lyu, Z.; Chen, L.; Xu, S.; Guo, J.; et al. Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science 2020, 368, eaba5435. [Google Scholar] [CrossRef] [PubMed]
- International Wheat Genome Sequencing Consortium (IWGSC). Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using the high-density 90,000 SNP array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Zhang, J.; Che, Y.; Liu, W.; Lu, Y.; Yang, X.; Li, X.; Jia, J.; Liu, X.; Li, L. Construction of Agropyron Gaertn. genetic linkage maps using a wheat 660K SNP array reveals a homoeologous relationship with the wheat genome. Plant Biotechnol. J. 2018, 16, 818–827. [Google Scholar] [CrossRef] [Green Version]
- Allen, A.M.; Winfield, M.O.; Burridge, A.J.; Downie, R.C.; Benbow, H.R.; Barker, G.L.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A.; et al. Characterization of a wheat breeders array suitable for high-throughput SNP genotyping of global varieties of hexaploid bread wheat (Triticum aestivum). Plant Biotechnol. J. 2017, 15, 390–401. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; He, Z.; Rasheed, A.; Wen, W.; Yan, J.; Zhang, P.; Wan, Y.; Zhang, Y.; Xie, C.; Xia, X. Genome-wide association mapping of black point reaction in common wheat (Triticum aestivum L.). BMC Plant Biol. 2017, 17, 220. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Pan, Y.; Singh, P.K.; He, X.; Ren, Y.; Zhao, L.; Zhang, N.; Cheng, S.; Chen, F. Investigation and genome-wide association study for Fusarium crown rot resistance in Chinese common wheat. BMC Plant Biol. 2019, 19, 153. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Zhang, F.; Yan, X.; Zhang, X.; Dong, Z.; Cui, D.; Chen, F. Genome-wide association study for 13 agronomic traits reveals distribution of superior alleles in bread wheat from the Yellow and Huai Valley of China. Plant Biotechnol. J. 2017, 15, 953–969. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Zhang, F.; Zhao, C.; Lv, G.; Sun, C.; Pan, Y.; Guo, X.; Chen, F. Genome-wide association study of six quality traits reveals the association of the TaRPP13L1 gene with flour colour in Chinese bread wheat. Plant Biotechnol. J. 2019, 17, 2106–2122. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Hu, X.; Shi, T.; Yin, H.; Sun, D.; Hao, Y.; Xia, X.; Luo, J.; Fernie, A.R.; He, Z.; et al. Metabolite-based genome-wide association study enables dissection of the flavonoid decoration pathway of wheat kernels. Plant Biotechnol. J. 2020, 18, 1722–1735. [Google Scholar] [CrossRef]
- Li, T.; Luo, M.; Zhang, D.; Wu, D.; Li, L.; Bai, G. Effective marker alleles associated with type 2 resistance to Fusarium head blight infection in fields. Breed Sci. 2016, 66, 350–357. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, A.; Zhu, S.; Li, L.; He, X.; Sun, Z.; Li, T. Basal rachis internode injection: A novel inoculation method to evaluate wheat resistance to Fusarium head blight. Phytopathology 2021, 111, 1670–1674. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixedeffects models using lme4. J. Stat. Software 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. Plink: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Z. GAPIT Version 3: Boosting power and accuracy for genomic association and prediction. Genom. Proteom. Bioinform. 2021, 19, 629–640. [Google Scholar] [CrossRef]
- Li, M.X.; Yeung, J.M.; Cherny, S.S.; Sham, P.C. Evaluating the effective numbers of independent tests and significant p-value thresholds in commercial genotyping arrays and public imputation reference datasets. Hum. Genet. 2012, 131, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Wang, M.; Wu, J.; Guo, W.; Chen, Y.; Li, G.; Wang, Y.; Shi, W.; Xia, G.; Fu, D.; et al. WheatOmics: A platform combining multiple omics data to accelerate functional genomics studies in wheat. Mol. Plant. 2021, 14, 1965–1968. [Google Scholar] [CrossRef] [PubMed]
- Cowger, C.; Smith, J.; Boos, D.; Bradley, C.A.; Ransom, J.; Bergstrom, G.C. Managing a destructive, episodic crop disease: A national survey of wheat and barley growers’ experience with Fusarium head blight. Plant Dis. 2020, 104, 634–648. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Hua, C.; Li, L.; Sun, Z.; Yuan, M.; Bai, G.; Humphreys, G.; Li, T. Integration of meta-QTL discovery with omics: Towards a molecular breeding platform for improving wheat resistance to Fusarium head blight. Crop J. 2021, 9, 739–749. [Google Scholar] [CrossRef]
- Nannuru, V.K.R.; Windju, S.S.; Belova, T.; Dieseth, J.A.; Alsheikh, M.; Dong, Y.; McCartney, C.A.; Henriques, M.A.; Buerstmayr, H.; Michel, S.; et al. Genetic architecture of Fusarium head blight disease resistance and associated traits in Nordic spring wheat. Theor. Appl. Genet. 2022, 135, 2247–2263. [Google Scholar] [CrossRef]
- Carpenter, N.R.; Wright, E.; Malla, S.; Singh, L.; Sanford, D.V.; Clark, A.; Harrison, S.; Murphy, J.P.; Costa, J.; Chao, S.; et al. Identification and validation of Fusarium head blight resistance QTL in the U.S. soft red winter wheat cultivar ‘Jamestown’. Crop Sci. 2020, 60, 2919–2930. [Google Scholar] [CrossRef]
- Petersen, S.; Lyerly, J.H.; McKendry, A.L.; Islam, M.S.; BrownGuedira, G.; Cowger, C.; Dong, Y.; Murphy, J.P. Validation of Fusarium head blight resistance QTL in US winter wheat. Crop Science. 2017, 57, 1–12. [Google Scholar] [CrossRef]
- Miedaner, T.; Risser, P.; Paillard, S.; Schnurbusch, T.; Keller, B.; Hartl, L.; Holzapfel, J.; Korzun, V.; Ebmeyer, E.; Friedrich Utz, H. Broad-spectrum resistance loci for three quantitatively inherited diseases in two winter wheat populations. Mol. Breed. 2012, 29, 731–742. [Google Scholar] [CrossRef]
- Xu, Q.; Xu, F.; Qin, D.; Li, M.; Fedak, G.; Cao, W.; Yang, L.; Dong, J. Molecular mapping of QTLs conferring Fusarium head blight resistance in Chinese wheat cultivar Jingzhou 66. Plants 2020, 9, 1021. [Google Scholar] [CrossRef]
- Paillard, S.; Schnurbusch, T.; Tiwari, R.; Messmer, M.; Winzeler, M.; Keller, B.; Schachermayr, G. QTL analysis of resistance to Fusarium head blight in Swiss winter wheat (Triticum aestivum L.). Theor. Appl. Genet. 2004, 109, 323–332. [Google Scholar] [CrossRef]
- Yang, J.; Bai, G.; Shaner, G.E. Novel quantitative trait loci (QTL) for Fusarium head blight resistance in wheat cultivar Chokwang. Theor. Appl. Genet. 2005, 111, 1571–1579. [Google Scholar] [CrossRef]
- Sari, E.; Berraies, S.; Knox, R.E.; Singh, A.K.; Ruan, Y.; Cuthbert, R.D.; Pozniak, C.J.; Henriquez, M.A.; Kumar, S.; Burt, A.J.; et al. High density genetic mapping of Fusarium head blight resistance QTL in tetraploid wheat. PLoS ONE 2018, 13, e0204362. [Google Scholar] [CrossRef] [Green Version]
- Srinivasachary; Gosman, N.; Steed, A.; Faure, S.; Nicholson, P. Mapping of QTL associated with Fusarium head blight in spring wheat RL4137. Czech J. Genet. Plant Breed. 2008, 44, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Schmolke, M.; Zimmermann, G.; Buerstmayr, H.; Schweizer, G.; Miedaner, T.; Korzun, V.; Ebmeyer, E.; Hartl, L. Molecular mapping of Fusarium head blight resistance in the winter wheat population Dream/Lynx. Theor. Appl. Genet. 2005, 111, 747–756. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| QTL | Marker Interval | Physical Position (Mb) | Physical Distance (Mb) | No. of Significant SNPs | PVE (%) | Average PVE (%) | Panels | No. Replications |
|---|---|---|---|---|---|---|---|---|
| gFHB-1A.1 | tplb0025b13_2054~Kukri_c4900_2435 | 3.4~46.2 | 43 | 26 | 5.1~19.3 | 9.4 | I & II | 5 |
| gFHB-1A.2 | RAC875_rep_c120644_74~BS00009104_51 | 527.2~534.3 | 7.1 | 15 | 7.2~15.9 | 9.8 | I & II | 4 |
| gFHB-1A.3 | AX-110478077~AX-94942875 | 579.4~591.3 | 11.9 | 25 | 5.0~17.3 | 6.3 | I & II | 5 |
| gFHB-1B | Kukri_c51864_225~AX-109320151 | 582.9~607.9 | 25 | 32 | 5.9~16.2 | 8.6 | I & II | 5 |
| gFHB-2B | Tdurum_contig96648_102~BS00065136_51 | 694.1~704.0 | 9.9 | 27 | 7.3~18.2 | 12.5 | II | 4 |
| gFHB-3A.1 | Tdurum_contig76679_307~IAAV1155 | 1.3~14.9 | 13.5 | 57 | 8.1~19.7 | 13.8 | II | 4 |
| gFHB-3A.2 | AX-110433978~AX-109645516 | 539.7~570.8 | 31.1 | 100 | 5.0~9.7 | 8.2 | I | 4 |
| gFHB-3B | Kukri_c64588_274~AX-110404403 | 3.3~14.3 | 11 | 12 | 6.1~17.8 | 11.4 | I & II | 5 |
| gFHB-3D | Excalibur_c9485_707~Excalibur_c60530_113 | 0.8~9.7 | 9 | 21 | 7.9~19.7 | 14.3 | Panel II | 4 |
| gFHB-5A | AX-89432740~Ku_c12469_983 | 577.9~596.5 | 18.6 | 33 | 8.8~13.4 | 10.7 | I & II | 5 |
| gFHB-5D | AX-111388087~BobWhite_c31_2478 | 524.8~565.7 | 41 | 13 | 5.9~12.9 | 9.8 | I & II | 7 |
| gFHB-6B | BS00003897_51~Kukri_c38732_546 | 132.1~151.1 | 19 | 13 | 8.8~16.9 | 12.1 | II | 4 |
| gFHB-7A | BobWhite_c17904_259~Tdurum_contig30621_328 | 8.3~68.0 | 59.8 | 29 | 7.1~16.6 | 12.2 | II | 4 |
| gFHB-7B | AX-109345488~Tdurum_contig10980_1229 | 128.9~199.8 | 70.9 | 114 | 5.8~10.4 | 8.4 | I & II | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, C.; Chao, H.; Sun, X.; Suo, Y.; Chen, Z.; Li, Z.; Ma, L.; Li, J.; Ren, Y.; Hua, W.; et al. Genome-Wide Association Study for Fusarium Head Blight Resistance in Common Wheat from China. Agronomy 2023, 13, 1712. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13071712
Shi C, Chao H, Sun X, Suo Y, Chen Z, Li Z, Ma L, Li J, Ren Y, Hua W, et al. Genome-Wide Association Study for Fusarium Head Blight Resistance in Common Wheat from China. Agronomy. 2023; 13(7):1712. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13071712
Chicago/Turabian StyleShi, Chaonan, Huaiyu Chao, Xiaonan Sun, Yongqiang Suo, Zeyu Chen, Zhikang Li, Lin Ma, Jiaqi Li, Yan Ren, Wei Hua, and et al. 2023. "Genome-Wide Association Study for Fusarium Head Blight Resistance in Common Wheat from China" Agronomy 13, no. 7: 1712. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13071712