

Arbuscular Mycorrhizal Fungi as a Plant Growth Stimulant in a Tomato and Onion Intercropping System
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Set Up and Crop Management
2.3. Root Colonization (%)
2.4. Root Morphology
2.5. Morpho-Agronomic Traits
2.6. Chlorophyll Contents
2.7. Leaf-Mineral Contents
2.8. Photosynthetic Activities
2.9. Statistical Analysis
3. Results
3.1. Root Colonization (%)
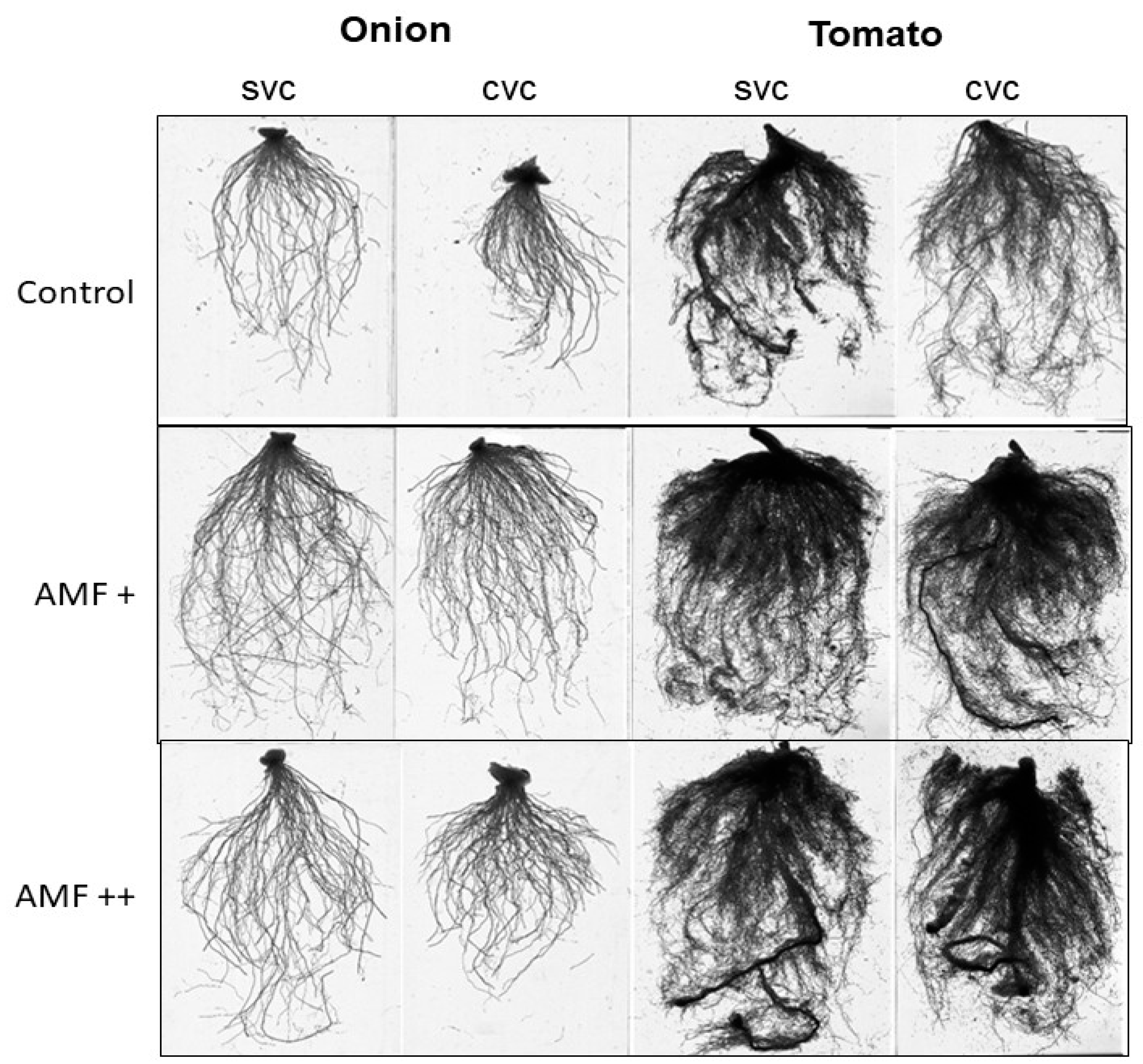
3.2. Effects on Root Morphology
3.3. Effect of AMF on Morpho-Agronomic Parameters
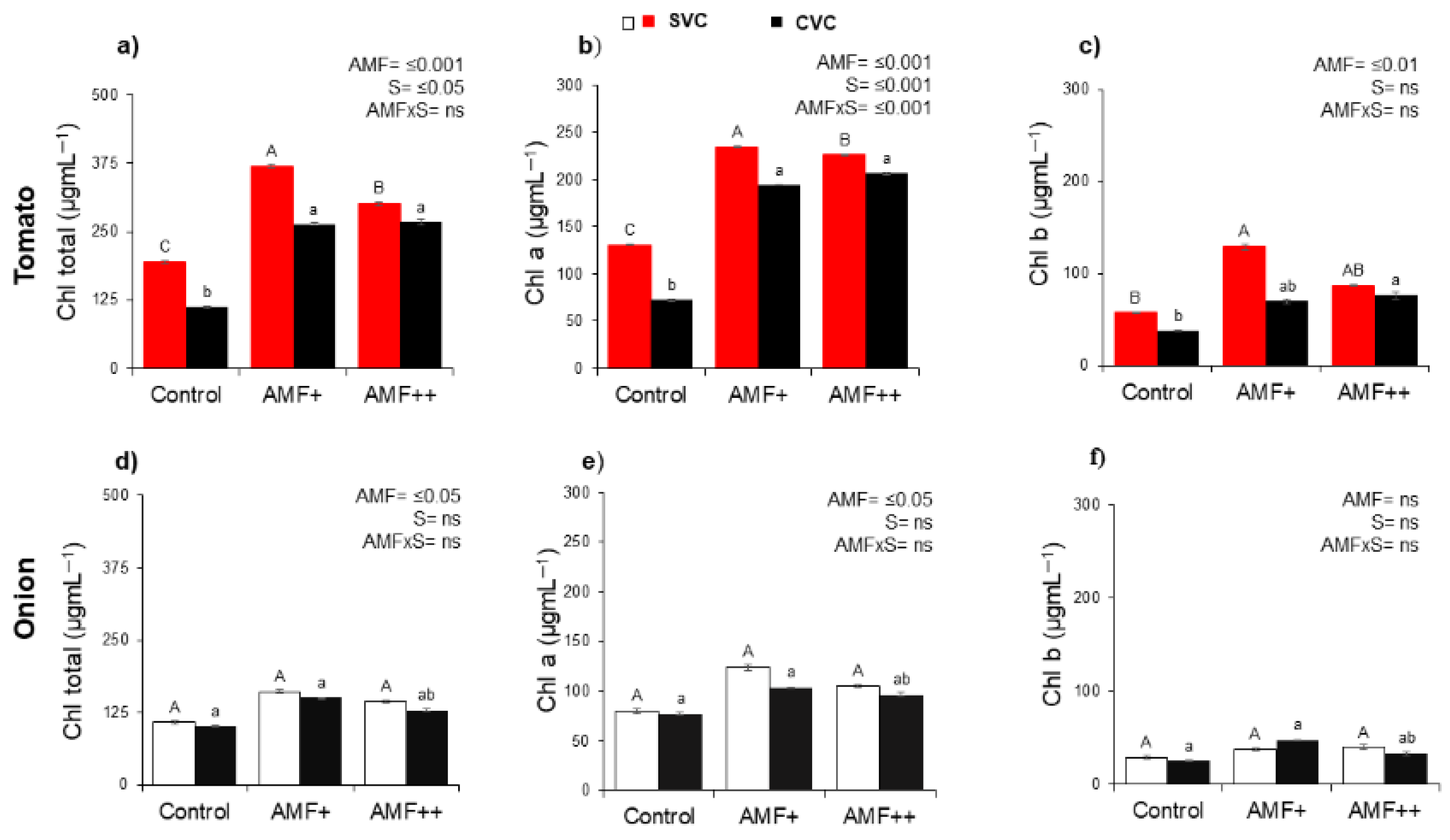
3.4. Chlorophyll Contents
3.5. Leaf Mineral Concentration
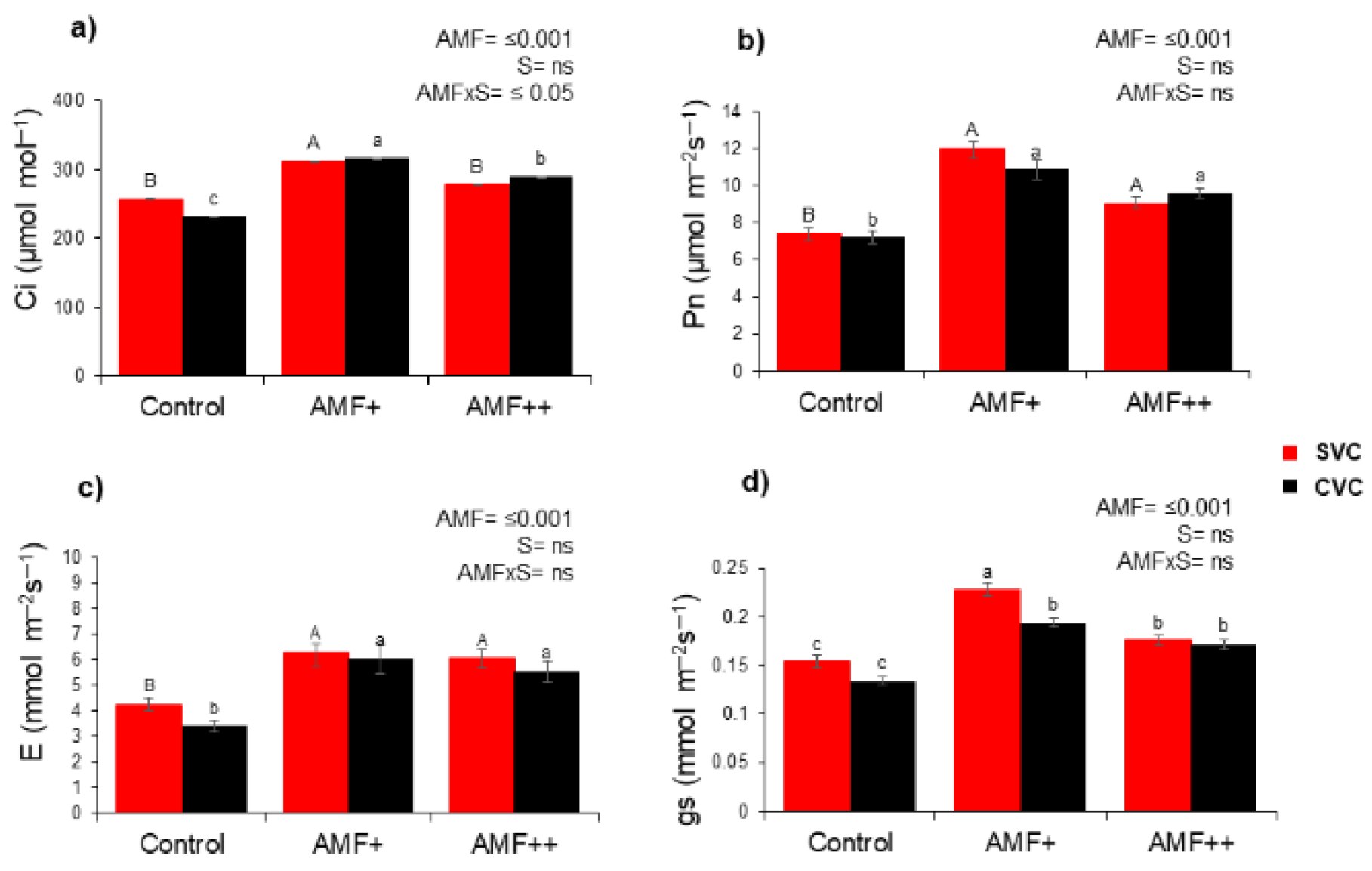
3.6. Photosynthetic Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Li, D.; Wan, S.; Wang, F.; Dou, W.; Xu, X.; Li, S.; Ma, R.; Qi, L. A long short-term memory-based model for greenhouse climate prediction. Int. J. Intell. Syst. 2022, 37, 135–151. [Google Scholar] [CrossRef]
- Mani, M. Pest Management in Horticultural Crops Under Protected Cultivation. In Trends in Horticultural Entomology; Springer: Berlin/Heidelberg, Germany, 2022; pp. 387–417. [Google Scholar]
- Cammarano, D.; Jamshidi, S.; Hoogenboom, G.; Ruane, A.C.; Niyogi, D.; Ronga, D. Processing tomato production is expected to decrease by 2050 due to the projected increase in temperature. Nat. Food 2022, 3, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Wako, F.-L.; Muleta, H.-D. The role of vermicompost application for tomato production: A review. J. Plant Nutr. 2022, 46, 129–144. [Google Scholar] [CrossRef]
- Montaño Méndez, I.E.; Valenzuela Patrón, I.N.; Villavicencio López, K.V. Competitiveness of the Mexican red tomato in the international market: Analysis 2003–2017. Rev. Mex. De Cienc. Agrícolas 2021, 12, 1185–1197. [Google Scholar]
- Przygocka-Cyna, K.; Barłóg, P.; Grzebisz, W.; Spiżewski, T. Onion (Allium cepa L.) Yield and Growth Dynamics Response to In-Season Patterns of Nitrogen and Sulfur Uptake. Agronomy 2020, 10, 1146. [Google Scholar] [CrossRef]
- Fabrice, D.; Yann, M.; Kémal, B. Onion (Allium Cepa) Production in Urban and Peri-Urban Areas: Financial Performance and Importance of This Activity for Market Gardeners in Southern Benin. Curr. Investig. Agric. Curr. Res. 2018, 3, 159. [Google Scholar]
- Thomas, B. Crop Development–Onion. In Edible Alliums: Botany, Production and Uses; CABI GB: Boston, MA, USA, 2022; pp. 51–67. [Google Scholar]
- Bhantana, P.; Rana, M.S.; Sun, X.-c.; Moussa, M.G.; Saleem, M.H.; Syaifudin, M.; Shah, A.; Poudel, A.; Pun, A.B.; Bhat, M.A. Arbuscular mycorrhizal fungi and its major role in plant growth, zinc nutrition, phosphorous regulation and phytoremediation. Symbiosis 2021, 84, 19–37. [Google Scholar] [CrossRef]
- Tringovska, I.; Yankova, V.; Markova, D.; Mihov, M. Effect of companion plants on tomato greenhouse production. Sci. Hortic. 2015, 186, 31–37. [Google Scholar] [CrossRef]
- Lithourgidis, A.; Dordas, C.; Damalas, C.A.; Vlachostergios, D. Annual intercrops: An alternative pathway for sustainable agriculture. Aust. J. Crop Sci. 2011, 5, 396–410. [Google Scholar]
- Tilman, D. Benefits of intensive agricultural intercropping. Nat. Plants 2020, 6, 604–605. [Google Scholar] [CrossRef] [PubMed]
- Basit, F.; Asghar, S.; Ahmed, T.; Ijaz, U.; Noman, M.; Hu, J.; Liang, X.; Guan, Y. Facile synthesis of nanomaterials as nanofertilizers: A novel way for sustainable crop production. Environ. Sci. Pollut. Res. 2022, 29, 51281–51297. [Google Scholar] [CrossRef] [PubMed]
- Bona, E.; Cantamessa, S.; Massa, N.; Manassero, P.; Marsano, F.; Copetta, A.; Lingua, G.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonads improve yield, quality and nutritional value of tomato: A field study. Mycorrhiza 2017, 27, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Naranjo-Ortiz, M.A.; Gabaldón, T. Fungal evolution: Diversity, taxonomy and phylogeny of the Fungi. Biol. Rev. 2019, 94, 2101–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Błaszkowski, J.; Czerniawska, B.; Wubet, T.; Schäfer, T.; Buscot, F.; Renker, C. Glomus irregulare, a new arbuscular mycorrhizal fungus in the Glomeromycota. Mycotaxon 2008, 106, 247–267. [Google Scholar]
- Stockinger, H.; Walker, C.; Schüßler, A. ‘Glomus intraradices DAOM197198’, a model fungus in arbuscular mycorrhiza research, is not Glomus intraradices. New Phytol. 2009, 183, 1176–1187. [Google Scholar] [CrossRef] [PubMed]
- Schüßler, A.; Walker, C. The Glomeromycota: A Species List with New Families and Genera; Amazon: Gloucester, UK, 2010. [Google Scholar]
- Roussis, I.; Beslemes, D.; Kosma, C.; Triantafyllidis, V.; Zotos, A.; Tigka, E.; Mavroeidis, A.; Karydogianni, S.; Kouneli, V.; Travlos, I. The Influence of Arbuscular Mycorrhizal Fungus Rhizophagus irregularis on the Growth and Quality of Processing Tomato (Lycopersicon esculentum Mill.) Seedlings. Sustainability 2022, 14, 9001. [Google Scholar] [CrossRef]
- Ye, Q.; Wang, H.; Li, H. Arbuscular Mycorrhizal Fungi Improve Growth, Photosynthetic Activity, and Chlorophyll Fluorescence of Vitis vinifera L. cv. Ecolly under Drought Stress. Agronomy 2022, 12, 1563. [Google Scholar] [CrossRef]
- Li, S.; Chi, S.; Lin, C.; Cai, C.; Yang, L.; Peng, K.; Huang, X.; Liu, J. Combination of biochar and AMF promotes phosphorus utilization by stimulating rhizosphere microbial co-occurrence networks and lipid metabolites of Phragmites. Sci. Total Environ. 2022, 845, 157339. [Google Scholar] [CrossRef] [PubMed]
- Sterrett, S.B. Composts as Horticultural Substrates for Vegetable Transplant Production. In Compost Utilization in Horticultural Cropping Systems; Lewis Publication: Boca Raton, FL, USA, 2001; pp. 227–240. [Google Scholar]
- Raviv, M.; Chen, Y.; Inbar, Y. Peat and peat substitutes as growth media for container-grown plants. In The Role of Organic Matter in Modern Agriculture; Springer: Berlin/Heidelberg, Germany, 1986; pp. 257–287. [Google Scholar]
- Herrera, F.; Castillo, J.; Chica, A.; Bellido, L.L. Use of municipal solid waste compost (MSWC) as a growing medium in the nursery production of tomato plants. Bioresour. Technol. 2008, 99, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Castillo, J.; Herrera, F.; López-Bellido, R.; López-Bellido, F.; López-Bellido, L.; Fernández, E. Municipal solid waste (MSW) compost as a tomato transplant medium. Compos. Sci. Util. 2004, 12, 86–92. [Google Scholar] [CrossRef]
- Burés, S. Sustratos; AbeBooks: Victoria, BC, Canada, 1997. [Google Scholar]
- Abad, M.; Noguera, P.; Puchades, R.; Maquieira, A.; Noguera, V. Physico-chemical and chemical properties of some coconut coir dusts for use as a peat substitute for containerised ornamental plants. Bioresour. Technol. 2002, 82, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Raviv, M.; Lieth, J.H.; Bar-Tal, A. Significance of soilless culture in agriculture. In Soil Culture; Academic Press: Cambridge, MA, USA, 2008; pp. 1–11. [Google Scholar]
- Shahid, A.; Hussain, R.; Riaz, A.; Younis, A.; Javaid, M.M.; Shah, S.Z.H.; Rashid, S.; Ali, S.; Haider, M.; Raza, A. Effect of different substrates on vegetative growth and quality of cast iron (Aspidistra elatior L.). Int. J. Biosci. 2017, 10, 297–308. [Google Scholar]
- Gamalero, E.; Trotta, A.; Massa, N.; Copetta, A.; Martinotti, M.G.; Berta, G. Impact of two fluorescent pseudomonads and an arbuscular mycorrhizal fungus on tomato plant growth, root architecture and P acquisition. Mycorrhiza 2004, 14, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Wolf, D.C.; Skipper, H.D. Soil sterilization. Methods Soil Anal.: Part 2 Microbiol. BioChem. Prop. 1994, 5, 41–51. [Google Scholar]
- Steiner, A.A. The universal nutrient solution. In Proceedings of the 6th International Congress on Soilless Culture, Lunteren, The Netherlands, 29 April–5 May 1984. [Google Scholar]
- Kormanik, P.; McGraw, A.; Schenck, N. Methods and principles of mycorrhizal research. Am. Phytopath. Soc. St Paul 1982, 134, 405. [Google Scholar]
- McGonigle, T.P.; Miller, M.H.; Evans, D.; Fairchild, G.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular—Arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Himmelbauer, M.L.; Loiskandl, A.W.; Kastanek, A.F. Estimating length, average diameter and surface area of roots using two different image analyses systems. Plant Soil 2004, 260, 111–120. [Google Scholar] [CrossRef]
- Bruuinsma, J. The quantitative analysis of chlorophylls a and b in plant extracts. Photochem. Photobiol. 1963, 2, 241–249. [Google Scholar] [CrossRef]
- Yang, Y.; Tang, M.; Sulpice, R.; Chen, H.; Tian, S.; Ban, Y. Arbuscular mycorrhizal fungi alter fractal dimension characteristics of Robinia pseudoacacia L. seedlings through regulating plant growth, leaf water status, photosynthesis, and nutrient concentration under drought stress. J. Plant Growth Regul. 2014, 33, 612–625. [Google Scholar] [CrossRef]
- Kalra, Y. Handbook of Reference Methods for Plant Analysis; CRC Press: Boca Raton, FL, USA, 1997. [Google Scholar]
- Hernández, A.; Castillo, H.; Ojeda, D.; Arras, A.; López, J.; Sánchez, E. Effect of vermicompost and compost on lettuce production. Chil. J. Agric. Res. 2010, 70, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Gong, X.; Zhang, X.; Zhang, W.; Sun, J.; Chen, B. Effects of arbuscular mycorrhizal fungi on photosynthesis, ion balance of tomato plants under saline-alkali soil condition. J. Plant Nutr. 2020, 43, 682–698. [Google Scholar] [CrossRef]
- Lanoue, J.; Leonardos, E.D.; Khosla, S.; Hao, X.; Grodzinski, B. Effect of elevated CO2 and spectral quality on whole plant gas exchange patterns in tomatoes. PLoS ONE 2018, 13, e0205861. [Google Scholar] [CrossRef] [Green Version]
- Gamalero, E.; Martinotti, M.; Trotta, A.; Lemanceau, P.; Berta, G. Morphogenetic modifications induced by Pseudomonas fluorescens A6RI and Glomus mosseae BEG12 in the root system of tomato differ according to plant growth conditions. New Phytol. 2002, 155, 293–300. [Google Scholar] [CrossRef]
- Hu, B.; Hu, S.; Vymazal, J.; Chen, Z. Application of arbuscular mycorrhizal fungi for pharmaceuticals and personal care productions removal in constructed wetlands with different substrate. J. Clean. Prod. 2022, 339, 130760. [Google Scholar] [CrossRef]
- Santoyo, G.; Kumar, A.; Aamir, M.; Sivakumar, U. Mitigation of Plant Abiotic Stress by Microorganisms: Applicability and Future Directions; Elsevier: Amsterdam, The Netherlands, 2022. [Google Scholar]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. BioChem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Gosling, P.; Mead, A.; Proctor, M.; Hammond, J.P.; Bending, G.D. Contrasting arbuscular mycorrhizal communities colonizing different host plants show a similar response to a soil phosphorus concentration gradient. New Phytol. 2013, 198, 546–556. [Google Scholar] [CrossRef]
- Sinclair, G.; Charest, C.; Dalpé, Y.; Khanizadeh, S. Influence of colonization by arbuscular mycorrhizal fungi on three strawberry cultivars under salty conditions. Agric. Food Sci. 2014, 23, 146–158. [Google Scholar] [CrossRef] [Green Version]
- González-González, M.F.; Ocampo-Alvarez, H.; Santacruz-Ruvalcaba, F.; Sánchez-Hernández, C.V.; Casarrubias-Castillo, K.; Becerril-Espinosa, A.; Castañeda-Nava, J.J.; Hernández-Herrera, R.M. Physiological, ecological, and biochemical implications in tomato plants of two plant biostimulants: Arbuscular mycorrhizal fungi and seaweed extract. Front. Plant Sci. 2020, 11, 999. [Google Scholar] [CrossRef]
- Fracasso, A.; Telò, L.; Lanfranco, L.; Bonfante, P.; Amaducci, S. Physiological beneficial effect of Rhizophagus intraradices inoculation on tomato plant yield under water deficit conditions. Agronomy 2020, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Cavagnaro, T.R.; Jackson, L.; Six, J.; Ferris, H.; Goyal, S.; Asami, D.; Scow, K. Arbuscular mycorrhizas, microbial communities, nutrient availability, and soil aggregates in organic tomato production. Plant Soil 2006, 282, 209–225. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Aroca, R. Modulation of Aquaporin Genes by the Arbuscular Mycorrhizal Symbiosis in Relation to Osmotic Stress Tolerance: Aquaporin in AM Plants Under Osmotic Stress. In Symbioses and Stress: Joint Ventures in Biology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 357–374. [Google Scholar]
- Rozpądek, P.; Rąpała-Kozik, M.; Wężowicz, K.; Grandin, A.; Karlsson, S.; Ważny, R.; Anielska, T.; Turnau, K. Arbuscular mycorrhiza improves yield and nutritional properties of onion (Allium cepa). Plant Physiol. Biochem. 2016, 107, 264–272. [Google Scholar] [CrossRef]
- Fussy, A.; Papenbrock, J. An overview of soil and soilless cultivation techniques—Chances, challenges and the neglected question of sustainability. Plants 2022, 11, 1153. [Google Scholar] [CrossRef] [PubMed]
- Busso, M.; Busso, M. Arbuscular mycorrhizal fungi and common mycorrhizal networks benefit plants through morphological, physiological and productive traits and soil quality. Lilloa 2022, 59, 301–317. [Google Scholar] [CrossRef]
- Bitterlich, M.; Sandmann, M.; Graefe, J. Arbuscular mycorrhiza alleviates restrictions to substrate water flow and delays transpiration limitation to stronger drought in tomato. Front. Plant Sci. 2018, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Salvioli, A.; Zouari, I.; Chalot, M.; Bonfante, P. The arbuscular mycorrhizal status has an impact on the transcriptome profile and amino acid composition of tomato fruit. BMC Plant Biol. 2012, 12, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajiboland, R.; Aliasgharzadeh, N.; Laiegh, S.F.; Poschenrieder, C. Colonization with arbuscular mycorrhizal fungi improves salinity tolerance of tomato (Solanum lycopersicum L.) plants. Plant Soil 2010, 331, 313–327. [Google Scholar] [CrossRef]
- Latef, A.A.H.A.; Chaoxing, H. Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Sci. Hortic. 2011, 127, 228–233. [Google Scholar] [CrossRef]
- Boyer, L.R.; Brain, P.; Xu, X.-M.; Jeffries, P. Inoculation of drought-stressed strawberry with a mixed inoculum of two arbuscular mycorrhizal fungi: Effects on population dynamics of fungal species in roots and consequential plant tolerance to water deficiency. Mycorrhiza 2015, 25, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Regvar, M.; Vogel-Mikuš, K. Arbuscular Mycorrhiza in Metal Hyperaccumulating Plants; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Jaiswal, D.K.; Krishna, R.; Chouhan, G.K.; de Araujo Pereira, A.P.; Ade, A.B.; Prakash, S.; Verma, S.K.; Prasad, R.; Yadav, J.; Verma, J.P. Bio-fortification of minerals in crops: Current scenario and future prospects for sustainable agriculture and human health. Plant Growth Regul. 2022, 98, 5–22. [Google Scholar] [CrossRef]
- Garcia, K.; Delaux, P.M.; Cope, K.R.; Ané, J.M. Molecular signals required for the establishment and maintenance of ectomycorrhizal symbioses. New Phytol. 2015, 208, 79–87. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Choudhary, A.K.; Pooniya, V.; Suri, V.K.; Singh, U. Soil factors associated with micronutrient acquisition in crops-biofortification perspective. In Biofortification of Food Crops; Springer: Berlin/Heidelberg, Germany, 2016; pp. 159–176. [Google Scholar]
- Tekaya, M.; Mechri, B.; Mbarki, N.; Cheheb, H.; Hammami, M.; Attia, F. Arbuscular mycorrhizal fungus Rhizophagus irregularis influences key physiological parameters of olive trees (Olea europaea L.) and mineral nutrient profile. Photosynthetica 2017, 55, 308–316. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Chanratana, M.; Kim, K.; Seshadri, S.; Sa, T. Impact of arbuscular mycorrhizal fungi on photosynthesis, water status, and gas exchange of plants under salt stress–A meta-analysis. Front. Plant Sci. 2019, 10, 457. [Google Scholar] [CrossRef] [PubMed]
- Gavito, M.E.; Jakobsen, I.; Mikkelsen, T.N.; Mora, F. Direct evidence for modulation of photosynthesis by an arbuscular mycorrhiza-induced carbon sink strength. New Phytol. 2019, 223, 896–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, D.P.; Read, D.J.; Scholes, J.D. Mycorrhizal sink strength influences whole plant carbon balance of Trifolium repens L. Plant Cell Environ. 1998, 21, 881–891. [Google Scholar] [CrossRef]
- Kaschuk, G.; Kuyper, T.W.; Leffelaar, P.A.; Hungria, M.; Giller, K.E. Are the rates of photosynthesis stimulated by the carbon sink strength of rhizobial and arbuscular mycorrhizal symbioses? Soil Biol. Bio-Chem. 2009, 41, 1233–1244. [Google Scholar] [CrossRef]
- Leventis, G.; Tsiknia, M.; Feka, M.; Ladikou, E.; Papadakis, I.; Chatzipavlidis, I.; Papadopoulou, K.; Ehaliotis, C. Arbuscular mycorrhizal fungi enhance growth of tomato under normal and drought conditions, via different water regulation mechanisms. Rhizosphere 2021, 19, 100394. [Google Scholar] [CrossRef]
- Othman, Y.; Bataineh, K.; Al-Ajlouni, M.; Alsmairat, N.; Ayad, J.; Shiyab, S.; Al-Qarallah, B.; St Hilaire, R. Soilless culture: Management of growing substrate, water, nutrient, salinity, microorganism and product quality. Fresenius Environ. Bull. 2019, 28, 3249–3260. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | pH | EC (dSm−1) | NO3− | NH4+ | PO43− | Fe2+ | Cu2+ | Zn2+ | Mn2+ |
|---|---|---|---|---|---|---|---|---|---|
| SVC | 7.4 | 0.2 | 5 | 3 | 0.70 | 0.46 | 0.1 | 0.03 | 0.03 |
| CVC | 7.6 | 1.2 | 3 | 5 | 13 | 0.73 | 0.2 | 0.2 | 1.3 |
| Treatments | RL (cm) | RD (cm) | RSA (cm2) | RV (cm3) | RF | RC | RT | |
|---|---|---|---|---|---|---|---|---|
| SVC | Control | 5132 ± 65 b | 0.66 ± 0.01 c | 993 ± 18 c | 30 ± 1.7 b | 34,463 ± 152 c | 3620 ± 93 b | 7049 ± 131 b |
| AMF+ | 5691 ± 73 a | 1.08 ± 0.04 a | 1440 ± 22 a | 44 ± 1.6 a | 43,923 ± 124 a | 4509 ± 93 a | 8578 ± 116 a | |
| AMF++ | 5483 ± 59 a | 0.83 ± 0.01 b | 1223 ± 33 b | 33 ± 1.7 b | 36,005 ± 109 b | 3752 ± 96 b | 7538 ± 98 b | |
| CVC | Control | 5013 ± 69 b | 0.81 ± 0.01 b | 1123 ± 14 c | 28 ± 2.1 b | 35,455 ± 199 c | 3635 ± 87 a | 7255 ± 92 a |
| AMF+ | 5467 ± 48 a | 1.10 ± 0.04 a | 1284 ± 12 a | 38 ± 1.4 a | 38,375 ± 178 b | 3945 ± 108 b | 7275 ± 135 a | |
| AMF++ | 5318 ± 40 a | 1.02 ± 0.03 a | 1207 ± 18 b | 33 ± 2.0 b | 40,404 ± 174 a | 3834 ± 131 a | 7420 ± 107 a | |
| AMF | <0.05 | <0.001 | <0.001 | <0.05 | <0.001 | <0.001 | <0.001 | |
| S | ns | <0.05 | <0.05 | ns | ns | <0.01 | <0.05 | |
| AMF × S | ns | ns | ns | ns | <0.001 | <0.001 | <0.05 |
| Treatments | RL (cm) | RD (cm) | RSA (cm2) | RV (cm3) | RF | RC | RT | |
|---|---|---|---|---|---|---|---|---|
| SVC | Control | 636 ± 1.2 b | 0.7 ± 0.09 c | 269 ± 1.17 c | 2.8 ± 0.03 b | 3173 ± 0.93 c | 925 ± 1.14 b | 111 ± 0.81 b |
| AMF+ | 1510 ± 1.2 a | 1.2 ± 0.05 a | 342 ± 0.98 a | 4.4 ± 0.04 a | 4438 ± 1.04 a | 1099 ± 0.77 a | 187 ± 1.12 a | |
| AMF++ | 1528 ± 1.5 a | 1.1 ± 0.05 b | 308 ± 1.28 b | 4.1 ± 0.05 a | 4249 ± 0.97 b | 1107 ± 0.78 a | 169 ± 1.04 a | |
| CVC | Control | 671 ± 1.0 c | 0.86 ± 0.04 b | 256 ± 1.14 a | 3.0 ± 0.05 a | 3325 ± 0.93 c | 980 ± 0.87 a | 120 ± 0.93 b |
| AMF+ | 1241 ± 1.7 b | 1.07 ± 0.06 a | 264 ± 1.96 a | 3.5 ± 0.05 a | 3966 ± 0.86 b | 997 ± 1.18 a | 159 ± 0.96 a | |
| AMF++ | 1316 ± 1.0 a | 1.1 ± 0.06 a | 276 ± 2.28 a | 3.5 ± 0.04 a | 4137 ± 0.85 a | 1022 ± 0.82 a | 141 ± 1.00 a | |
| AMF | <0.001 | <0.001 | <0.05 | <0.001 | <0.001 | <0.001 | <0.05 | |
| S | <0.001 | ns | <0.01 | <0.001 | <0.001 | <0.05 | <0.001 | |
| AMF × S | <0.001 | <0.001 | ns | <0.001 | <0.001 | <0.01 | <0.001 |
| Treatments | N (%) | P (%) | Ct (%) | Fe (ppm) | Cu (ppm) | Zn (ppm) | Mn (ppm) | |
|---|---|---|---|---|---|---|---|---|
| SVC | Control | 1.56 ± 0.07 b | 0.34 ± 0.08 b | 34 ± 0.67 a | 38 ± 1.21 c | 6 ± 0.45 c | 15 ± 2.07 b | 142 ± 0.20 b |
| AMF+ | 3.25 ± 0.07 a | 0.76 ± 0.05 a | 42 ± 0.33 a | 189 ± 0.34 a | 12 ± 0.26 b | 30 ± 0.35 a | 637 ± 1.01 a | |
| AMF++ | 2.71 ± 0.04 a | 0.51 ± 0.08 a | 42 ± 0.13 a | 129 ± 0.90 b | 16 ± 0.54 a | 21 ± 0.36 a | 362 ± 0.78 b | |
| CVC | Control | 1.93 ± 0.03 b | 0.47 ± 0.05 a | 32 ± 0.60 a | 56 ± 0.69 c | 8 ± 0.31 b | 11 ± 0.52 b | 93 ± 0.65 c |
| AMF+ | 3.06 ± 0.46 a | 0.68 ± 0.05 a | 39 ± 0.32 a | 163 ± 0.66 a | 19 ± 0.68 a | 35 ± 0.47 a | 124 ± 0.76 a | |
| AMF++ | 1.95 ± 0.11 b | 0.46 ± 0.04 a | 40 ± 0.14 a | 77 ± 0.78 b | 19 ± 0.43 a | 21 ± 0.31 a | 167 ± 0.55 b | |
| AMF | <0.001 | ns | ns | <0.001 | <0.001 | <0.001 | <0.001 | |
| S | <0.05 | ns | <0.001 | <0.001 | ns | <0.001 | <0.001 | |
| AMF × S | <0.01 | ns | ns | <0.001 | ns | <0.001 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shafiq, M.; Casas-Solís, J.; Neri-Luna, C.; Kiran, M.; Yasin, S.; González-Eguiarte, D.R.; Muñoz-Urias, A. Arbuscular Mycorrhizal Fungi as a Plant Growth Stimulant in a Tomato and Onion Intercropping System. Agronomy 2023, 13, 2003. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13082003
Shafiq M, Casas-Solís J, Neri-Luna C, Kiran M, Yasin S, González-Eguiarte DR, Muñoz-Urias A. Arbuscular Mycorrhizal Fungi as a Plant Growth Stimulant in a Tomato and Onion Intercropping System. Agronomy. 2023; 13(8):2003. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13082003
Chicago/Turabian StyleShafiq, Muhammad, Josefina Casas-Solís, Cecilia Neri-Luna, Munazza Kiran, Saba Yasin, Diego Raymundo González-Eguiarte, and Alejandro Muñoz-Urias. 2023. "Arbuscular Mycorrhizal Fungi as a Plant Growth Stimulant in a Tomato and Onion Intercropping System" Agronomy 13, no. 8: 2003. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13082003