
Genome-Wide Association Analysis Reveals the Gene Loci of Yield Traits under Drought Stress at the Rice Reproductive Stage

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Field Experiment
2.2. Statistical Analysis of Phenotypic Data
2.3. Genotyping
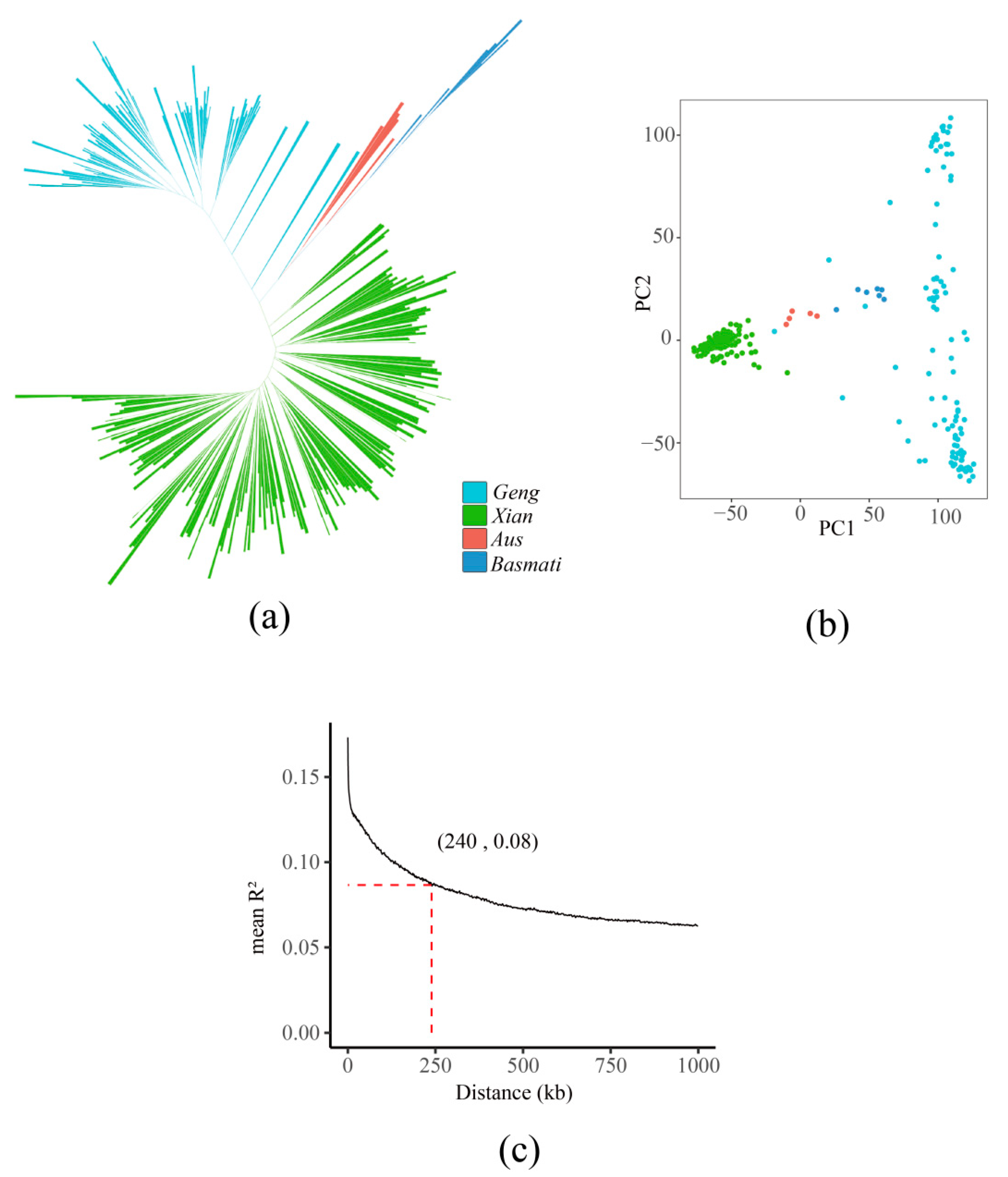
2.4. Population Genetic Analysis
2.5. Genome-Wide Association Study
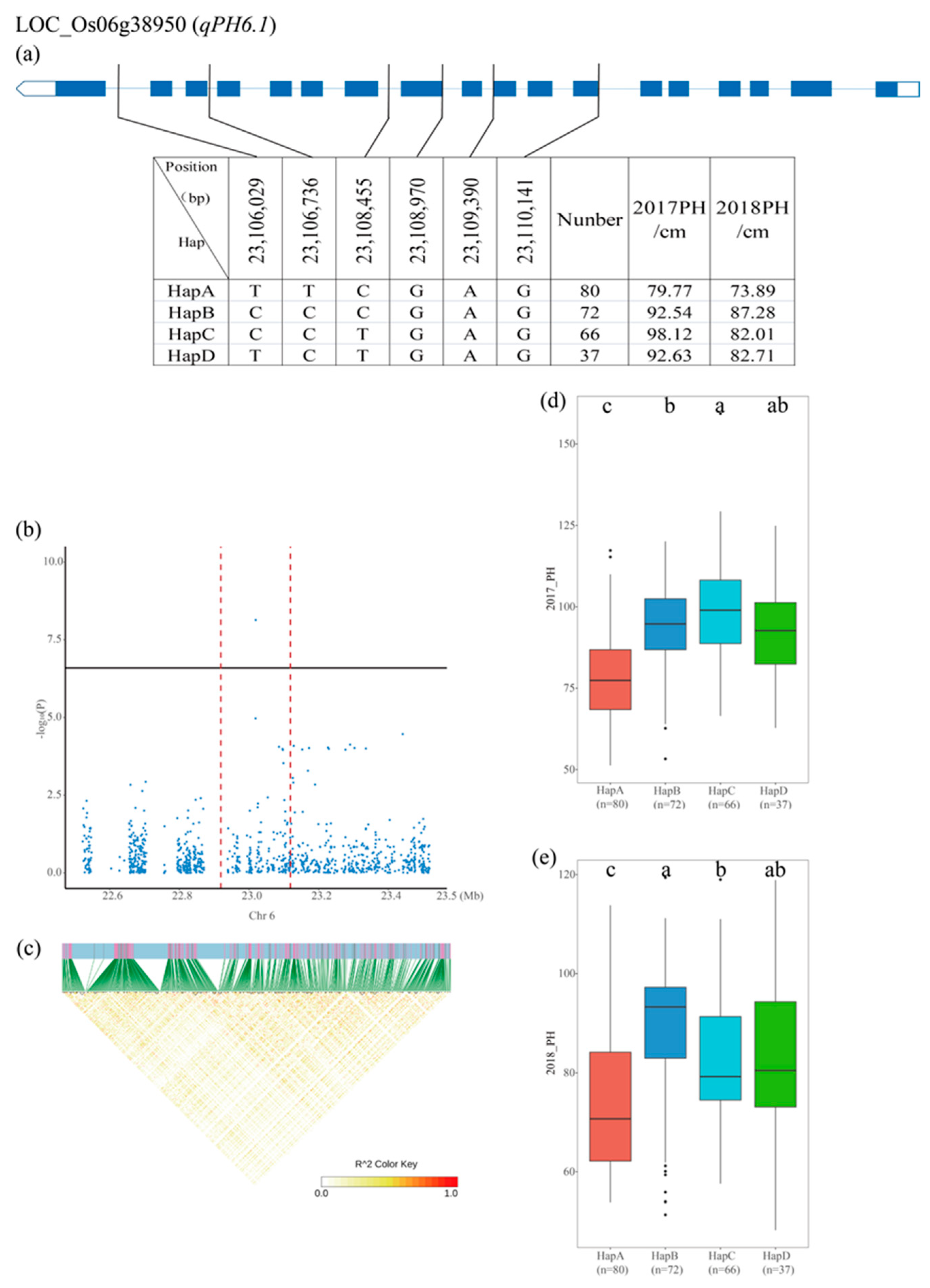
2.6. Identification of Candidate Genes and Haplotype Analysis
3. Results
3.1. Phenotypic Variations and Correlations
3.2. Phylogenetic and Population Structure Analysis
3.3. Genome-Wide Association Study of GYP, GNP, PNP, and PH
3.4. Candidate Gene Identification and Haplotype Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, X.; Xiong, H.; Jiang, C.; Zhang, D.; Yang, Z.; Huang, Y.; Zhu, W.; Ma, S.; Duan, J.; Wang, X.; et al. Natural variation of DROT1 confers drought adaptation in upland rice. Nat. Commun. 2022, 13, 4265. [Google Scholar] [CrossRef] [PubMed]
- Sheng-Chang, W.; Hai-Fu, T.U.; Dan, H.U.; Nai, W.U.; Xiang, C.; Li-Zhong, X. The exploitation of rice functional genes for abiotic stress. Chin. Bull. Life Sci. 2016, 28, 10. [Google Scholar]
- Teng, Z.; Chen, Y.; Meng, S.; Duan, M.; Zhang, J.; Ye, N. Environmental Stimuli: A Major Challenge during Grain Filling in Cereals. Int. J. Mol. Sci. 2023, 24, 2255. [Google Scholar] [CrossRef]
- Deng, X.; Singh, R.B.; Liu, J.; Güneralp, B. Physical and economic processes of water scarcity and water allocation for integrat-ed river basin management. Phys. Chem. Earth Parts A/B/C 2015, 79–82, 1. [Google Scholar]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Swamy, B.M.; Kumar, A. Genomics-based precision breeding approaches to improve drought tolerance in rice. Biotechnol. Adv. 2013, 31, 1308–1318. [Google Scholar] [CrossRef] [PubMed]
- Levitt, J. Responses of Plants to Environmental Stress, Volume 1: Chilling, Freezing, and High Temperature Stresses, 2nd ed.; Academic Press: Cambridge, MA, USA, 1980; Volume 38, p. 5. [Google Scholar]
- Costa, M.-C.D.; Farrant, J.M. Plant Resistance to Abiotic Stresses. Plants 2019, 8, 553. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-W.; Tian, H.-G.; Li, C.-G.; Chen, F.-Y.; Sun, Y.-X. Study on Drought Resistance of Rice. North Rice 2015, 45, 2. [Google Scholar]
- Yi, Y.; Wang, S.; Cheng, X.; Zhu, Q.; Li, Y.; Wang, F.; Zhang, M. Advances in Research on Drought Resistance in Rice. Agric. Biotechnol. 2022, 11, 26–33. [Google Scholar]
- Xu, M.; Jiang, X.; Zhou, G.; Chen, L. Relation between Rice (Oryza sativa L.) Drought resistance and Its Ability to Resist Water escaping, Proline and Sugar. Lifeence Res. 1998, 2, 113–117. [Google Scholar]
- Panda, D.; Mishra, S.S.; Behera, P.K. Drought Tolerance in Rice: Focus on Recent Mechanisms and Approaches. Rice Sci. 2021, 28, 119–132. [Google Scholar] [CrossRef]
- Mall, S.A.K. Biochemical Assimilates Accumulation and Remobilization at Reproduction Stage Drought Stress in Upland Rice (Oryza sativa L.). Environ. Ecol. 2017, 35, 2811–2818. [Google Scholar]
- Mohanty, S.K.; Sridhar, R. Physiology of rice tungro virus disease: Proline accumulation due to infection. Physiol. Plant. 1982, 56, 89–93. [Google Scholar] [CrossRef]
- Uga, Y.; Okuno, K.; Yano, M. Dro1, a major QTL involved in deep rooting of rice under upland field conditions. J. Exp. Bot. 2011, 62, 2485–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monir, M.M.; Jun, Z. Tools for quantitative trait locus mapping and genome-wide association study mapping: A review. J. Zhejiang Univ. Agric. Life Sci. 2014, 40, 379–386. [Google Scholar]
- Martin, N. Genetics of Cognitive Measures and Correlated Traits; School of Psychology, The University of Queensland: St Lucia, QLD, Australia, 2012. [Google Scholar]
- Kumari, M.; Muduli, L.; Meher, P.K.; Pradhan, S.K. Genome-Wide Association Study (GWAS) for Trait Analysis in Crops; Springer: New York, NY, USA, 2022; pp. 295–307. [Google Scholar] [CrossRef]
- Liu, Q.; Tang, Y.; Xiao, R.; Zhang, X.; Li, J. Progress on Molecular Research of Drought Resistance in Rice. Mol. Plant Breed. 2019, 17, 2841–2849. [Google Scholar]
- Bao-Feng, C.; Hong-Jian, L.I.; Meng-Ben, W.; Li-Min, L. Advance of Biomolecular Studies on Plant Drought Resistance Mecha-nisms. J. Shanxi Univ. (Nat. Sci. Ed.) 1999, 2, 400–405. [Google Scholar]
- Bernardo, R. Molecular Markers and Selection for Complex Traits in Plants: Learning from the Last 20 Years. Crop Sci. 2008, 48, 1649–1664. [Google Scholar] [CrossRef] [Green Version]
- Maisura; Chozin, M.A.; Lubis, I.; Junaedi, A.; Ehara, H. Some physiological character responses of rice under drought condi-tions in a paddy system. J. Int. Soc. Southeast Asian Agric. Sci. 2014, 20, 104–114. [Google Scholar]
- Luo, L.H.; Liu, G.H.; Xiao, Y.H.; Tang, W.B.; Chen, L.Y. Influences of High-Temperature Stress on the Fertility of Pollen, Spikelet and Grain-Weight in Rice. J. Hunan Agric. Univ. 2005, 31, 593. [Google Scholar]
- Pan, X.; Li, Y.; Zhang, H.; Liu, S.; Deng, H.; Li, X. Screening and Comparative Analysis of Drought Tolerance of Rice Varieties at the Reproductive Stage. Crop Sci. 2016, 57, 395–403. [Google Scholar] [CrossRef]
- Vivekanandan, P.Y. Association analysis in F1 and F2 generations of rice under reproductive stage drought stress. Electron. J. Plant Breed. 2010, 1, 890–898. [Google Scholar]
- Venuprasad, R.; Bool, M.E.; Quiatchon, L.; Cruz, M.T.S.; Amante, M.; Atlin, G.N. A large-effect QTL for rice grain yield under upland drought stress on chromosome 1. Mol. Breed. 2011, 30, 535–547. [Google Scholar] [CrossRef]
- Vikram, P.; Swamy, B.M.; Dixit, S.; Ahmed, H.U.; Cruz, M.T.S.; Singh, A.K.; Kumar, A. qDTY 1.1, a major QTL for rice grain yield under reproductive-stage drought stress with a consistent effect in multiple elite genetic backgrounds. BMC Genet. 2011, 12, 89. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Sandhu, N.; Singh, V.K.; Catolos, M.; Kumar, A. Genotyping-by-sequencing based QTL mapping for rice grain yield under reproductive stage drought stress tolerance. Sci. Rep. 2019, 9, 14326. [Google Scholar] [CrossRef] [Green Version]
- Yano, K.; Yamamoto, E.; Aya, K.; Takeuchi, H.; Lo, P.-C.; Hu, L.; Yamasaki, M.; Yoshida, S.; Kitano, H.; Hirano, K.; et al. Genome-wide association study using whole-genome sequencing rapidly identifies new genes influencing agronomic traits in rice. Nat. Genet. 2016, 48, 927–934. [Google Scholar] [CrossRef]
- Mansueto, L. SNP-Seek Database; International Rice Research Institute: Laguna, Philippines, 2015. [Google Scholar]
- Lin, Y.-L.; Wu, D.-H.; Wu, C.-C.; Huang, Y.-F. Explore the genetics of weedy traits using rice 3K database. Bot. Stud. 2021, 62, 2. [Google Scholar] [CrossRef]
- Qi, Y.; Wang, S.; Shen, C.; Zhang, S.; Chen, Y.; Xu, Y.; Liu, Y.; Wu, Y.; Jiang, D. OsARF12, a transcription activator on auxin response gene, regulates root elongation and affects iron accumulation in rice (Oryza sativa). New Phytol. 2011, 193, 109–120. [Google Scholar] [CrossRef]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast model-based estimation of ancestry in unrelated individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Bengough, A.G.; White, P.J. Preface. J. Exp. Bot. 2013, 64, 1179. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Dong, S.-S.; Xu, J.-Y.; He, W.-M.; Yang, T.-L. PopLDdecay: A fast and effective tool for linkage disequilibrium decay analysis based on variant call format files. Bioinformatics 2019, 35, 1786–1788. [Google Scholar] [CrossRef]
- Pantalião, G.F.; Narciso, M.; Guimarães, C.; Castro, A.; Colombari, J.M.; Breseghello, F.; Rodrigues, L.; Vianello, R.P.; Borba, T.O.; Brondani, C. Genome wide association study (GWAS) for grain yield in rice cultivated under water deficit. Genetica 2016, 144, 651–664. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.-X.; Yeung, J.M.Y.; Cherny, S.S.; Sham, P.C. Evaluating the effective numbers of independent tests and significant p-value thresholds in commercial genotyping arrays and public imputation reference datasets. Hum. Genet. 2011, 131, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Chen, H.; Qian, Y.; Liang, Z.; Zheng, G.; Xiang, J.; Feng, T.; Li, M.; Zeng, W.; Bao, Y.; et al. Genome-Wide Association Study of Rice Grain Shape and Chalkiness in a Worldwide Collection of Xian Accessions. Plants 2023, 12, 419. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Han, Y.; Liu, L.; Chen, Y.; Du, Y.; Zhang, J.; Sun, H.; Zhao, Q. qRT9, a quantitative trait locus controlling root thickness and root length in upland rice. J. Exp. Bot. 2015, 66, 2723–2732. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Huang, L.; Wang, M.; Cui, Y.; Xia, X. OsDSR-1, a calmodulin-like gene, improves drought tolerance through scavenging of reactive oxygen species in rice (Oryza sativa L.). Mol. Breed. 2017, 37, 75. [Google Scholar] [CrossRef]
- Gao, W.; Li, M.; Yang, S.; Gao, C.; Su, Y.; Zeng, X.; Jiao, Z.; Xu, W.; Zhang, M.; Xia, K. miR2105 and the kinase OsSAPK10 co-regulate OsbZIP86 to mediate drought-induced ABA biosynthesis in rice. Plant Physiol. 2022, 189, 889–905. [Google Scholar] [CrossRef] [PubMed]
- Dong, N.Q.; Sun, Y.; Guo, T.; Shi, C.L.; Lin, H.X. UDP-glucosyltransferase regulates grain size and abiotic stress tolerance associated with metabolic flux redirection in rice. Nat. Commun. 2020, 11, 2629. [Google Scholar] [CrossRef]
- Huang, L.; Wang, Y.; Wang, W.; Zhao, X.; Qin, Q.; Sun, F.; Hu, F.; Zhao, Y.; Li, Z.; Fu, B.; et al. Characterization of Transcription Factor Gene OsDRAP1 Conferring Drought Tolerance in Rice. Front. Plant Sci. 2018, 9, 94. [Google Scholar] [CrossRef] [Green Version]
- Ji, Q.; Zhang, L.S.; Wang, Y.F.; Wang, J. Genome-wide analysis of basic leucine zipper transcription factor families in Arabidopsis thaliana, Oryza sativa and Populus trichocarpa. J. Shanghai Univ. 2009, 13, 174–182. [Google Scholar] [CrossRef]
- Ni, E.; Deng, L.; Chen, H.; Lin, J.; Ruan, J.; Liu, Z.; Zhuang, C.; Zhou, H. OsCER1 regulates humidity-sensitive genic male sterility through very-long-chain (VLC) alkane metabolism of tryphine in rice. Funct. Plant Biol. 2021, 48, 461. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Han, S.; Zhou, X.; Zhao, C.; Guo, L.; Zhang, J.; Liu, F.; Huo, Q.; Zhao, W.; Guo, Z.; et al. Phosphorylation and ubiquitination of OsWRKY31 are integral to OsMKK10-2-mediated defense responses in rice. Plant Cell 2023, 35, 2391–2412. [Google Scholar] [CrossRef]
- Shi, F.; Dong, Y.; Wang, M.; Qiu, D. Transcriptomics analyses reveal that OsMIOX improves rice drought tolerance by regulating the expression of plant hormone and sugar related genes. Plant Biotechnol. Rep. 2020, 14, 339–349. [Google Scholar] [CrossRef]
- Arenhart, R.A.; De Lima, J.C.; Pedron, M.; Carvalho, F.E.L.; Da Silveira, J.A.G.; Rosa, S.B.; Caverzan, A.; Andrade, C.M.B.; Schünemann, M.; Margis, R.; et al. Involvement of ASR genes in aluminium tolerance mechanisms in rice. Plant Cell Environ. 2012, 36, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Carrari, F.; Cabrera-Ponce, J.L.; Vázquez-Rovere, C.; Herrera-Estrella, L.; Gudesblat, G.; Iusem, N.D. Analysis of an abscisic acid (ABA)-responsive gene promoter belonging to the Asr gene family from tomato in homologous and heterologous systems. Mol. Gen. Genet. MGG 1998, 258, 1–8. [Google Scholar] [CrossRef]
- Kumar, A.; Thomas, J.; Basu, S.; Ramegowda, H.V.; Pereira, A. Genome-Wide Association Study for Water Use Efficiency and Drought Resistance Traits in Rice. In Proceedings of the Plants & Animal Genome XXV Conference, San Diego, CA, USA, 14–18 January 2017. [Google Scholar]
- Xiang, J.; Zhang, C.; Wang, N.; Liang, Z.; Zheng, Z.; Liang, L.; Yuan, H.; Shi, Y. Genome-Wide Association Study Reveals Candidate Genes for Root-Related Traits in Rice. Curr. Issues Mol. Biol. 2022, 44, 4386–4405. [Google Scholar] [CrossRef]
- Byrt, C.S.; Millar, A.H.; Munns, R. Staple crops equipped for alkaline soils. Nat. Biotechnol. 2023, 41, 911–912. [Google Scholar] [CrossRef]
- Li, Y.; Han, S.; Sun, X.; Khan, N.U.; Zhong, Q.; Zhang, Z.; Zhang, H.; Ming, F.; Li, Z.; Li, J. Variations in OsSPL10 confer drought tolerance by directly regulating OsNAC2 expression and ROS production in rice. J. Integr. Plant Biol. 2023, 65, 918–933. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, R.; Huang, K.; Wei, Z.; Guo, B.; Huang, S.; Li, Z.; Jiang, W.; Wu, T.; Du, X. OsWRKY76 positively regulates drought stress via OsbHLH148-mediated jasmonate signaling in rice. Front. Plant Sci. 2023, 14, 1168723. [Google Scholar] [CrossRef]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive Analysis of NAC Family Genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef]
- Chen, Y.; Qi, H.; Yang, L.; Xu, L.; Wang, J.; Guo, J.; Zhang, L.; Tan, Y.; Pan, R.; Shu, Q.; et al. The OsbHLH002/OsICE1-OSH1 module orchestrates secondary cell wall formation in rice. Cell Rep. 2023, 42, 112702. [Google Scholar] [CrossRef]
- Zhang, C.; Li, M.; Rey, J.D.; Feng, T.; Lafitte, R.; Zheng, T.; Lv, Y.; Wu, F.; Fu, B.; Xu, J.; et al. Simultaneous improvement and genetic dissection of drought and submergence tolerances in rice (Oryza sativa L.) by selective introgression. Front. Plant Sci. 2023, 14, 1134450. [Google Scholar] [CrossRef]
- Man, J.; Gallagher, J.P.; Bartlett, M. Structural evolution drives diversification of the large LRR-RLK gene family. New Phytol. 2020, 226, 1492–1505. [Google Scholar] [CrossRef] [Green Version]
- Sreeman, S.M.; Vijayaraghavareddy, P.; Sreevathsa, R.; Rajendrareddy, S.; Arakesh, S.; Bharti, P.; Dharmappa, P.; Soolanayakanahally, R. Corrigendum: Introgression of Physiological Traits for a Comprehensive Improvement of Drought Adaptation in Crop Plants. Front. Chem. 2018, 6, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Gao, T.; Liu, C.; Mao, K.; Gong, X.; Li, C.; Ma, F. Fruit crops combating drought: Physiological responses and regulatory pathways. Plant Physiol. 2023, 192, 1768–1784. [Google Scholar] [CrossRef] [PubMed]
- Tax, F. Receptor-like Kinases in Plants: From Development to Defense; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Nodine, M.D.; Bryan, A.C.; Racolta, A.; Jerosky, K.V.; Tax, F.E. A few standing for many: Embryo receptor-like kinases. Trends Plant Sci. 2011, 16, 211–217. [Google Scholar] [CrossRef]
- Chen, X.; Ding, Y.; Yang, Y.; Song, C.; Wang, B.; Yang, S.; Guo, Y.; Gong, Z. Protein kinases in plant responses to drought, salt, and cold stress. J. Integr. Plant Biol. 2021, 63, 53–78. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.; Aalen, R.B.; Audenaert, D.; Beeckman, T.; Broadley, M.R.; Butenko, M.A.; Caño-Delgado, A.I.; de Vries, S.; Dresselhaus, T.; Felix, G.; et al. Tackling Drought Stress: Receptor-like Kinases Present New Approaches. Plant Cell 2012, 24, 2262–2278. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Song, L.; Xiao, Y.; Zhong, X.; Wang, J.; Xu, W.; Jiang, C.-Z. Overexpression of Myrothamnus flabellifolia MfWRKY41 confers drought and salinity tolerance by enhancing root system and antioxidation ability in Arabidopsis. Front. Plant Sci. 2022, 13, 967352. [Google Scholar] [CrossRef]
- Chu, X.; Wang, C.; Chen, X.; Lu, W.; Li, H.; Wang, X.; Hao, L.; Guo, X. Correction: The Cotton WRKY Gene GhWRKY41 Positively Regulates Salt and Drought Stress Tolerance in Transgenic Nicotiana benthamiana. PLoS ONE 2016, 11, e0157026. [Google Scholar] [CrossRef]
- Do, T.H.T.; Martinoia, E.; Lee, Y. Functions of ABC transporters in plant growth and development. Curr. Opin. Plant Biol. 2018, 41, 32–38. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Year | Mean ± SD | Max. | Mim. | CV (%) | H2 (%) |
|---|---|---|---|---|---|---|
| GYP (g) | 2017 | 15.60 ± 6.12 | 31.52 | 2.58 | 39.20% | 87.83% |
| 2018 | 17.81 ± 8.83 | 57.70 | 3.32 | 49.55% | ||
| GNP | 2017 | 118.06 ± 36.17 | 258.20 | 46.40 | 30.64% | 86.58% |
| 2018 | 98.90 ± 35.55 | 243.20 | 17.30 | 35.95% | ||
| PNP | 2017 | 9.85 ± 3.01 | 20.10 | 3.00 | 30.57% | 91.52% |
| 2018 | 9.80 ± 2.68 | 21.80 | 3.80 | 27.38% | ||
| PH (cm) | 2017 | 89.37 ± 16.53 | 159.35 | 51.30 | 18.49% | 95.41% |
| 2018 | 81.07 ± 15.64 | 119.40 | 48.20 | 19.29% |
| Trait | QTL | Chr | Lead SNP (bp) | p-Value | R2(%) | Known Genes/QTLs |
|---|---|---|---|---|---|---|
| GYP | qGYP3.1 | 3 | 27,772,039 | 1.18 × 10−7 | 11.53% | |
| qGYP4.1 | 4 | 1,152,913 | 8.14 × 10−9 | 11.74% | ||
| qGYP4.2 | 4 | 17,125,743 | 1.32 × 10−7 | 11.06% | ||
| qGYP9.1 | 9 | 17,087,568 | 1.55 × 10−7 | 10.72% | OsbHLH120 [40] | |
| qGYP10.1 | 10 | 5,284,683 | 1.35 × 10−7 | 9.81% | OsDSR-1 [41] | |
| qGYP12.1 | 12 | 7,352,766 | 1.54 × 10−7 | 10.74% | OsbZIP86 [42] | |
| GNP | qGNP1.1 | 1 | 16,207,018 | 1.0885 × 10−7 | 11.32% | |
| qGNP1.2 | 1 | 22,754,300 | 1.2248 × 10−7 | 9.84% | ||
| qGNP1.3 | 1 | 33,438,426 | 6.4749 × 10−8 | 11.54% | ||
| qGNP2.1 | 2 | 7,820,832 | 1.9861 × 10−7 | 10.82% | ||
| qGNP2.2 | 2 | 26,903,813 | 1.1258 × 10−7 | 11.86% | ||
| qGNP3.1 | 3 | 22,285,491 | 1.2046 × 10−7 | 9.65% | ||
| qGNP3.2 | 3 | 31,270,536 | 2.0749 × 10−7 | 10.65% | GSA1 [43] | |
| qGNP4.1 | 4 | 17,330,090 | 1.1495 × 10−7 | 11.30% | ||
| qGNP4.2 | 4 | 21,350,438 | 4.6361 × 10−8 | 12.09% | ||
| qGNP4.3 | 4 | 27,685,824 | 1.5817 × 10−7 | 11.00% | ||
| qGNP5.1 | 5 | 16,722,283 | 9.5616 × 10−8 | 11.19% | ||
| qGNP6.1 | 6 | 12,945,373 | 1.1852 × 10−7 | 9.72% | ||
| qGNP6.2 | 6 | 26,015,898 | 3.1678 × 10−8 | 13.06% | ||
| qGNP7.1 | 7 | 14,661,723 | 7.5111 × 10−8 | 11.76% | ||
| qGNP8.1 | 8 | 19,621,731 | 1.1988 × 10−7 | 9.68% | OsERF48 [44] | |
| qGNP9.1 | 9 | 17,087,568 | 6.4013 × 10−9 | 13.19% | OsbHLH120 [40] | |
| qGNP10.1 | 10 | 7,535,578 | 1.571 × 10−7 | 11.65% | ||
| qGNP11.1 | 11 | 2,559,169 | 1.0094 × 10−7 | 10.71% | OsZIP-2a [45] | |
| qGNP11.2 | 11 | 7,024,423 | 2.2725 × 10−8 | 12.24% | ||
| qGNP12.1 | 12 | 4,422,919 | 1.211 × 10−7 | 9.88% | ||
| PNP | qPNP2.1 | 2 | 24,673,219 | 1.71 × 10−8 | 11.24% | OsGL1-4 [46] |
| qPNP4.1 | 4 | 26,468,735 | 2.42 × 10−7 | 9.23% | OsRDCP1 [47] | |
| qPNP6.1 | 6 | 21,507,174 | 8.22 × 10−8 | 12.72% | OsMIOX [48] | |
| qPNP7.1 | 7 | 20,370,395 | 1.17 × 10−8 | 12.93% | ||
| qPNP8.1 | 8 | 6,254,665 | 1.83 × 10−8 | 11.13% | ||
| qPNP8.2 | 8 | 18,098,606 | 1.58 × 10−7 | 11.79% | ||
| qPNP9.1 | 9 | 6,714,468 | 1.58 × 10−8 | 17.73% | ||
| qPNP11.1 | 11 | 24,881,480 | 8.02 × 10−8 | 10.01% | ||
| qPNP11.2 | 11 | 27,691,415 | 1.79 × 10−7 | 12.04% | ||
| PH | qPH1.1 | 1 | 38,561,974 | 2.04E-13 | 20.49% | |
| qPH1.2 | 1 | 42,302,961 | 2.33 × 10−7 | 9.88% | Asr2 [49] | |
| qPH2.1 | 2 | 17,805,111 | 2.12 × 10−8 | 12.47% | ||
| qPH4.1 | 4 | 12,049,704 | 2.29 × 10−8 | 12.02% | ||
| qPH5.1 | 5 | 7,670,182 | 1.84 × 10−8 | 12.00% | ||
| qPH5.2 | 5 | 22,224,178 | 2.12 × 10−8 | 13.50% | ||
| qPH6.1 | 6 | 23,018,981 | 7.29 × 10−9 | 13.06% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, N.; Gao, Z.; Zhang, W.; Qian, Y.; Bai, D.; Zhao, X.; Bao, Y.; Zheng, Z.; Wang, X.; Li, J.; et al. Genome-Wide Association Analysis Reveals the Gene Loci of Yield Traits under Drought Stress at the Rice Reproductive Stage. Agronomy 2023, 13, 2096. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13082096
Wang N, Gao Z, Zhang W, Qian Y, Bai D, Zhao X, Bao Y, Zheng Z, Wang X, Li J, et al. Genome-Wide Association Analysis Reveals the Gene Loci of Yield Traits under Drought Stress at the Rice Reproductive Stage. Agronomy. 2023; 13(8):2096. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13082096
Chicago/Turabian StyleWang, Nansheng, Zhiyuan Gao, Wanyang Zhang, Yingzhi Qian, Di Bai, Xueyu Zhao, Yaling Bao, Zhenzhen Zheng, Xingmeng Wang, Jianfeng Li, and et al. 2023. "Genome-Wide Association Analysis Reveals the Gene Loci of Yield Traits under Drought Stress at the Rice Reproductive Stage" Agronomy 13, no. 8: 2096. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13082096