
The Key Physiological and Biochemical Traits Underlying Common Bean (Phaseolus vulgaris L.) Response to Iron Deficiency, and Related Interrelationships


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Growth Conditions
2.2. Spad Index
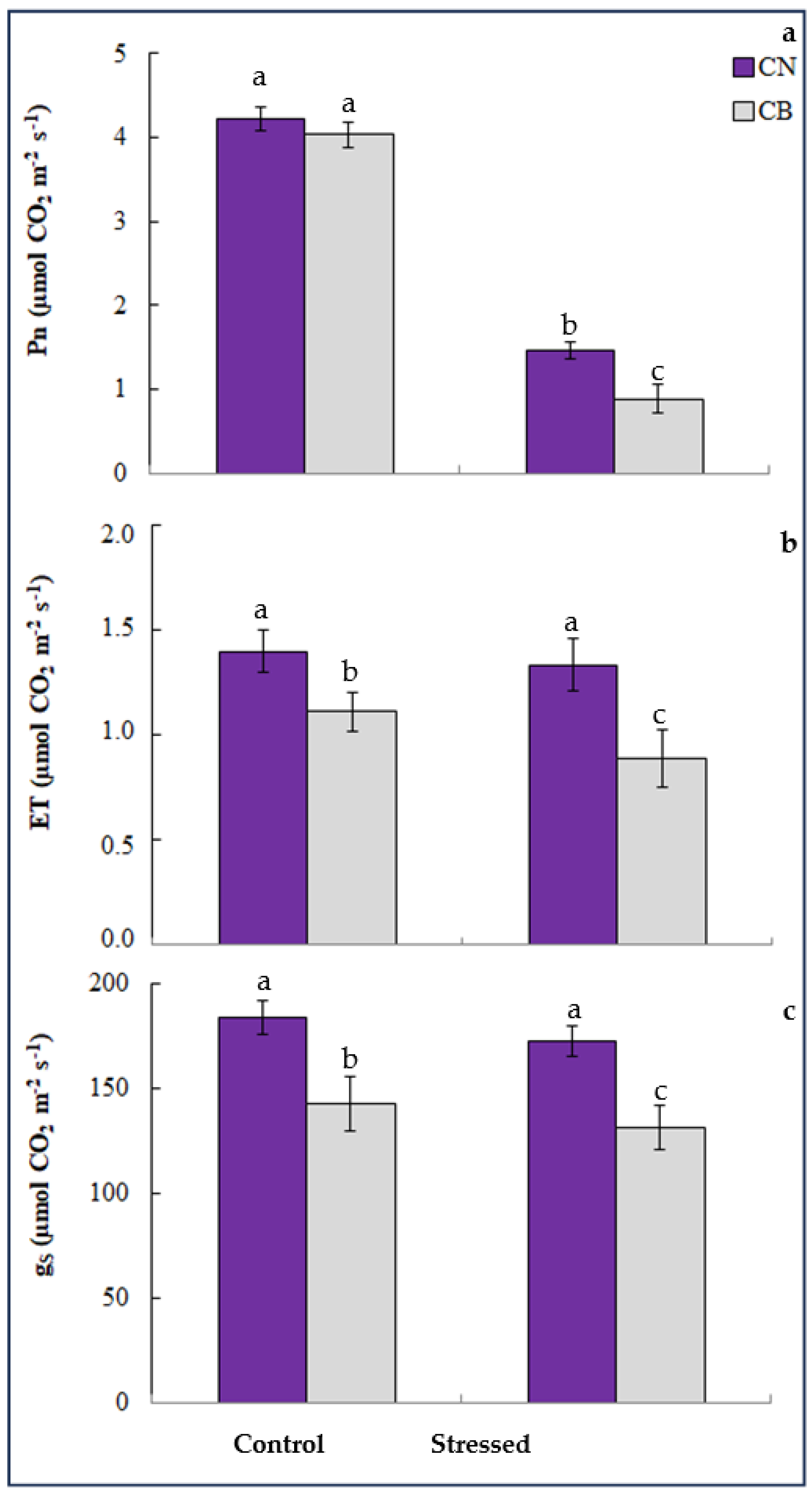
2.3. Net Photosynthesis and Gas Exchange
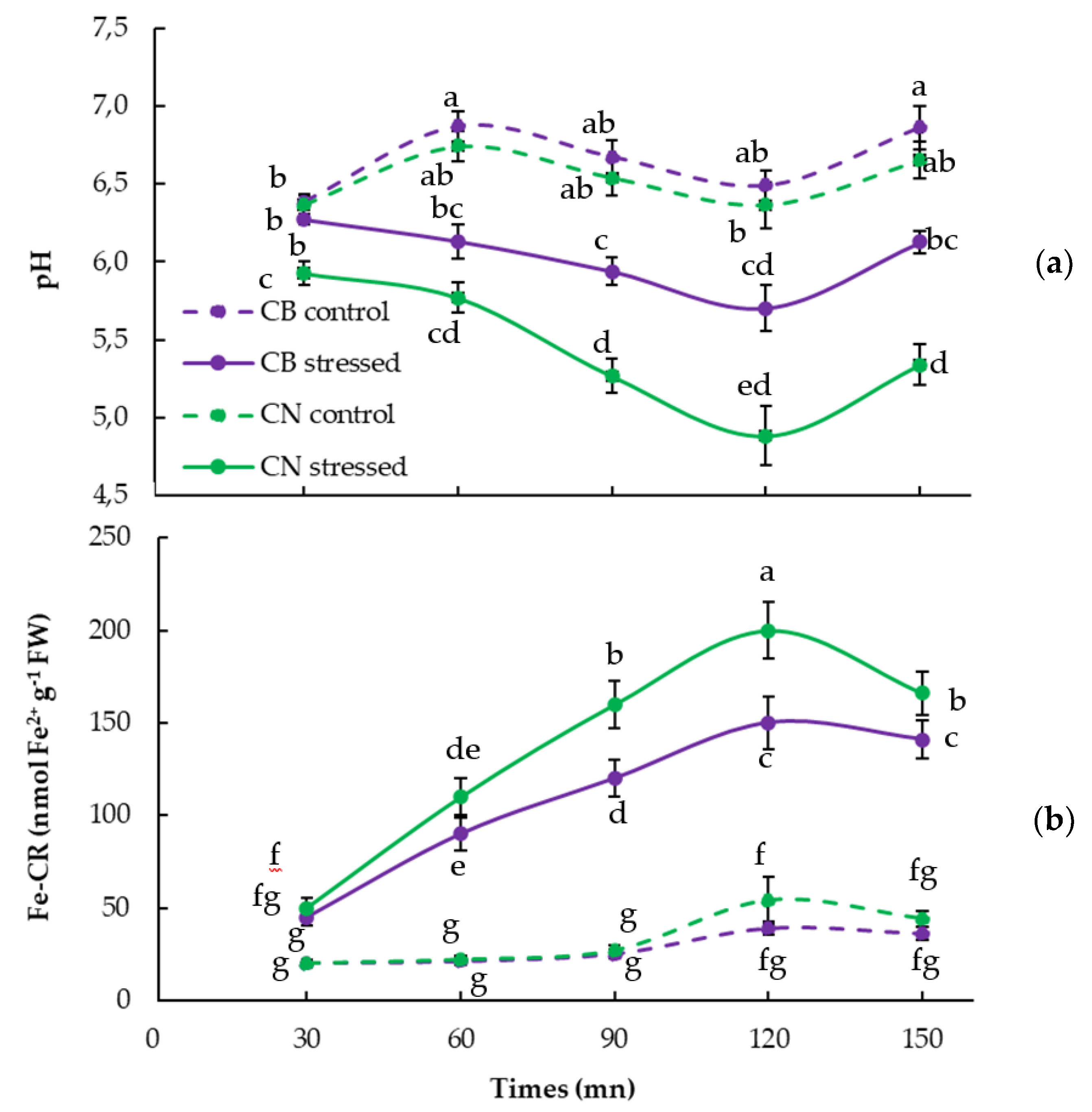
2.4. Acidification Capacity (H-ATPase Activity)
2.5. Fe Chelates Reductase Activity (Fe-CR)
2.6. Determination of Photosynthetic Pigments
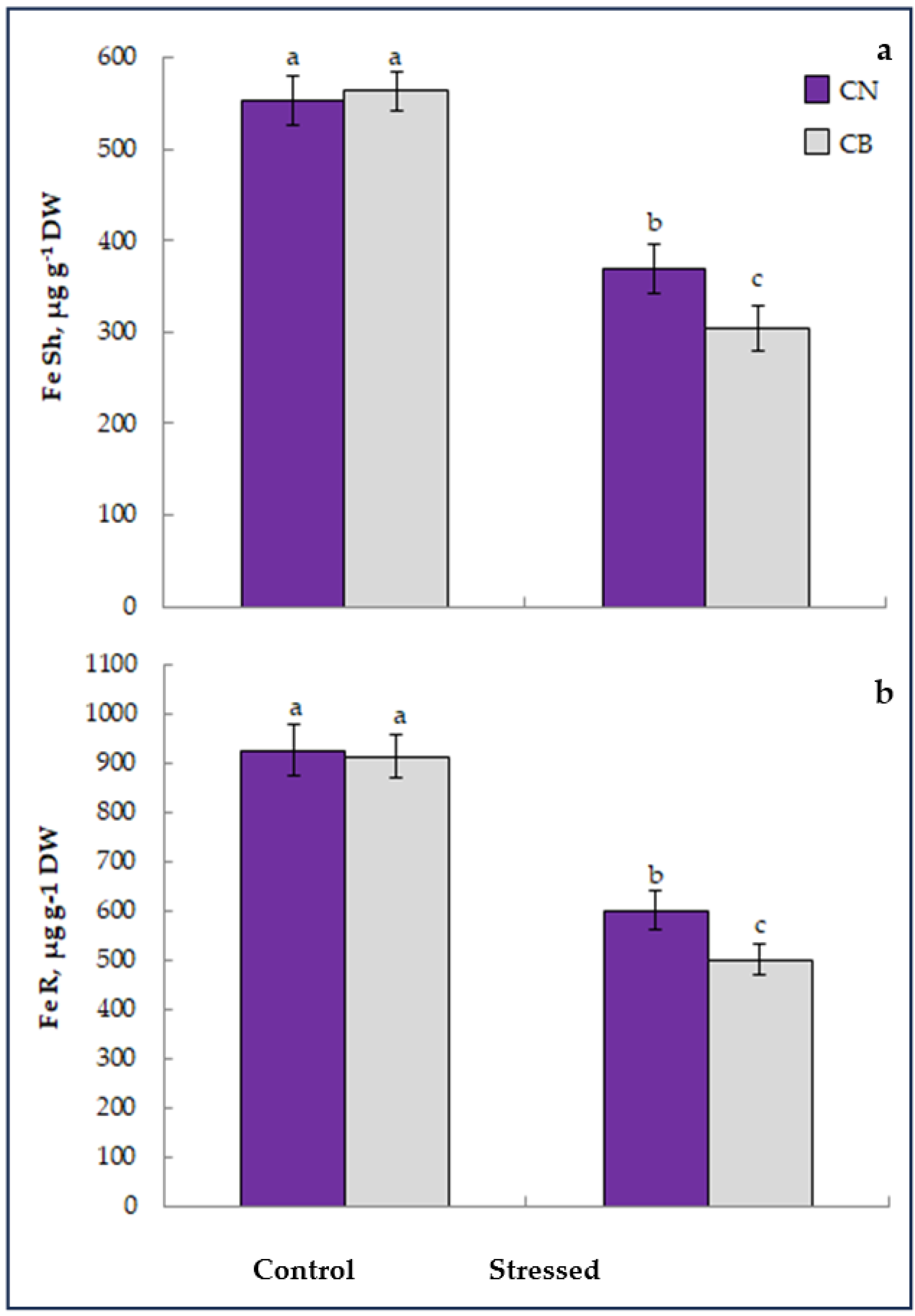
2.7. Determination of Active Iron
2.8. Calculation and Analyzed Parameters
2.9. Statistical Analysis
3. Results
3.1. Photosynthetic Pigments, Gas Exchange Parameters, and Plant Growth
3.2. Iron Nutrition and Distribution
3.3. Rhizosphere Acidification (H-ATPase) and Fe-Chelate Reductase (Fe-CR) Activities
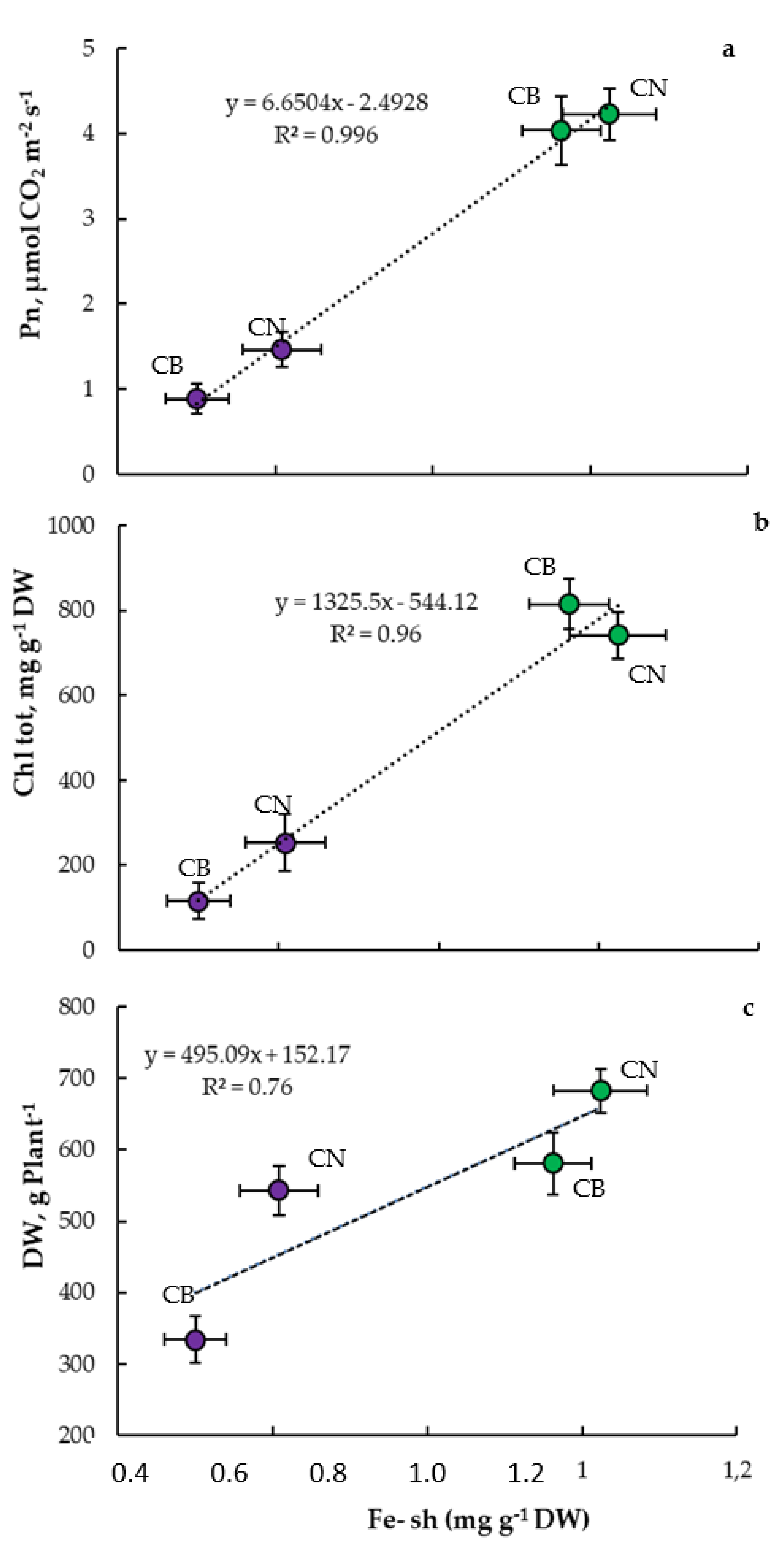
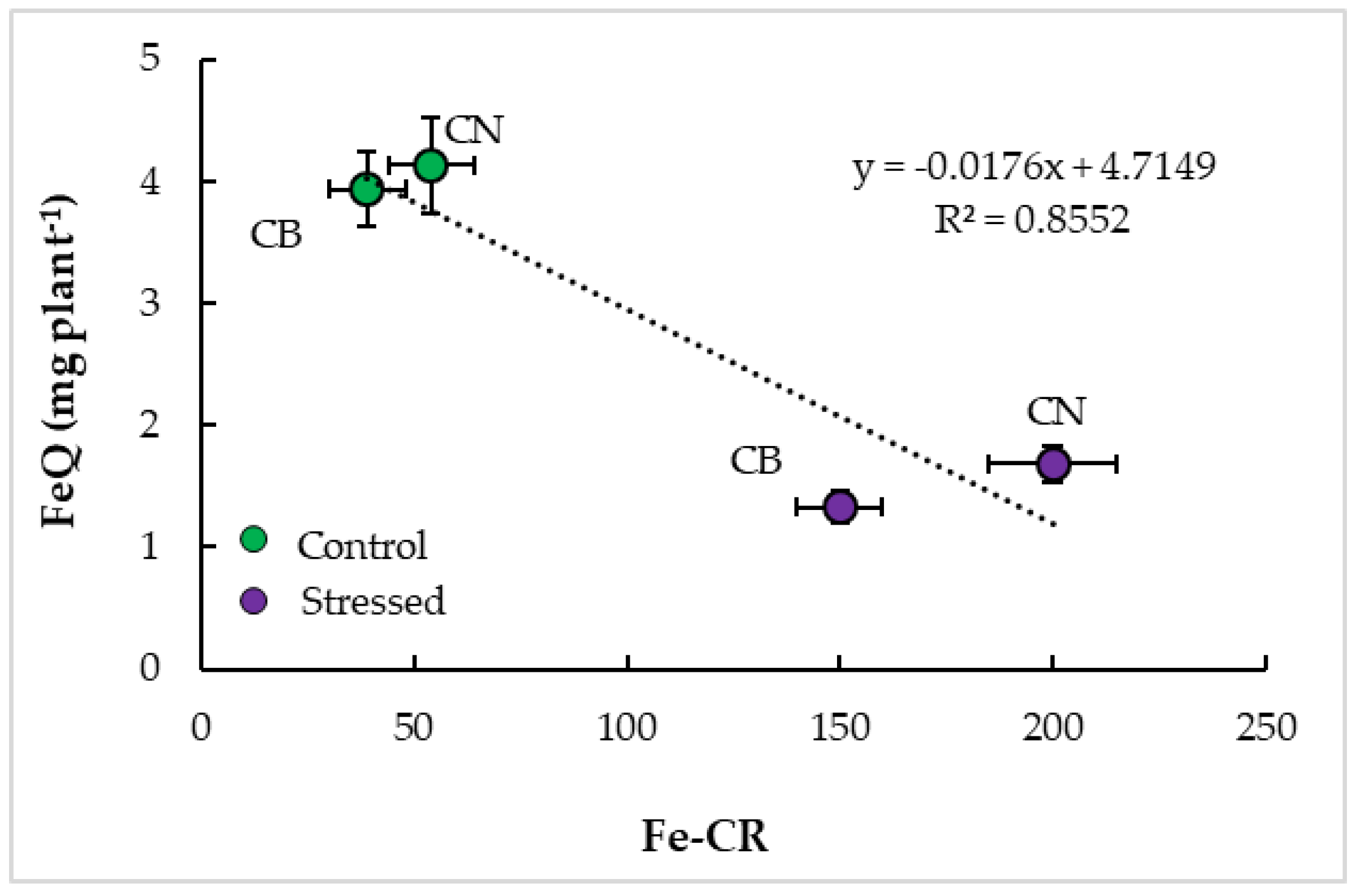
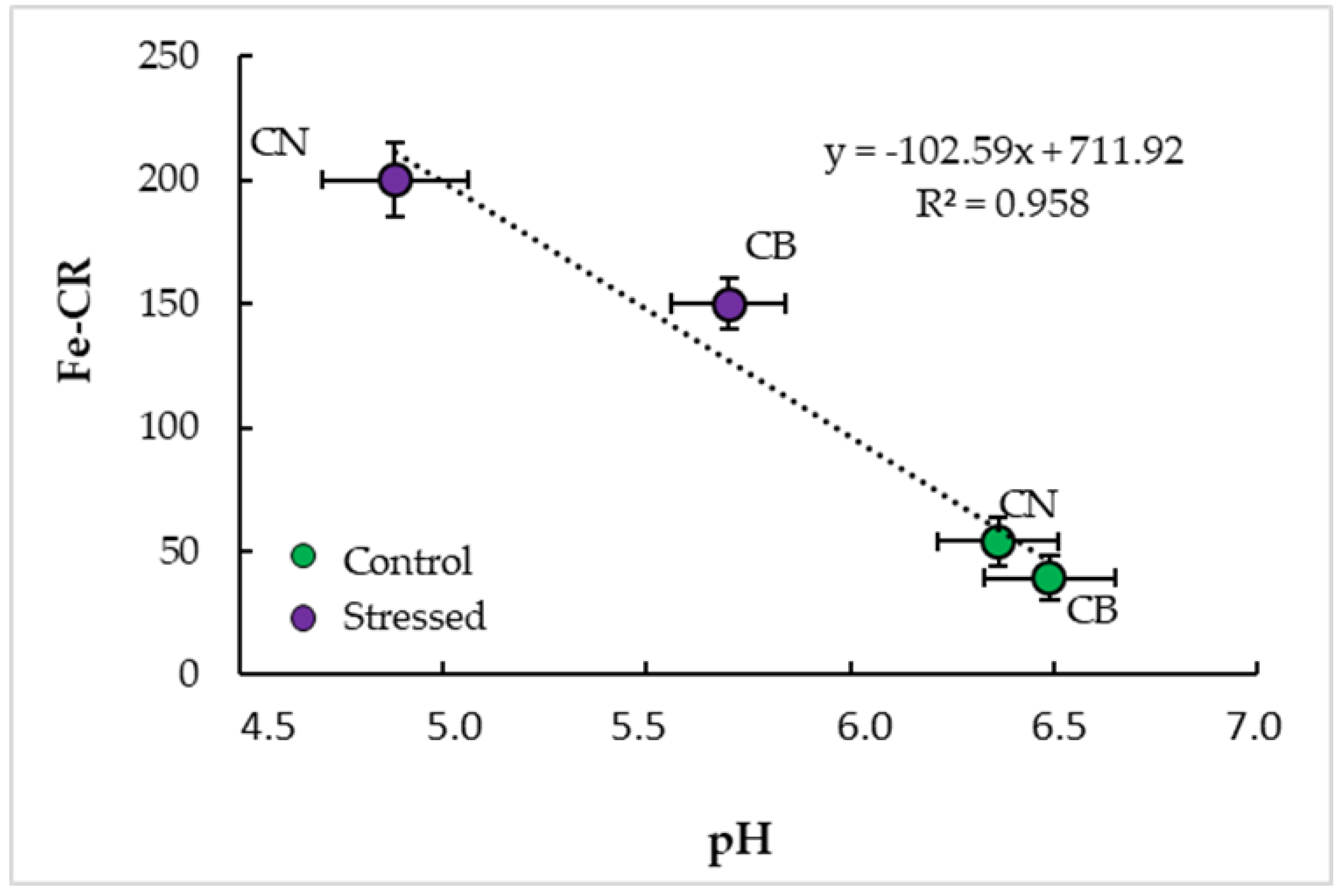
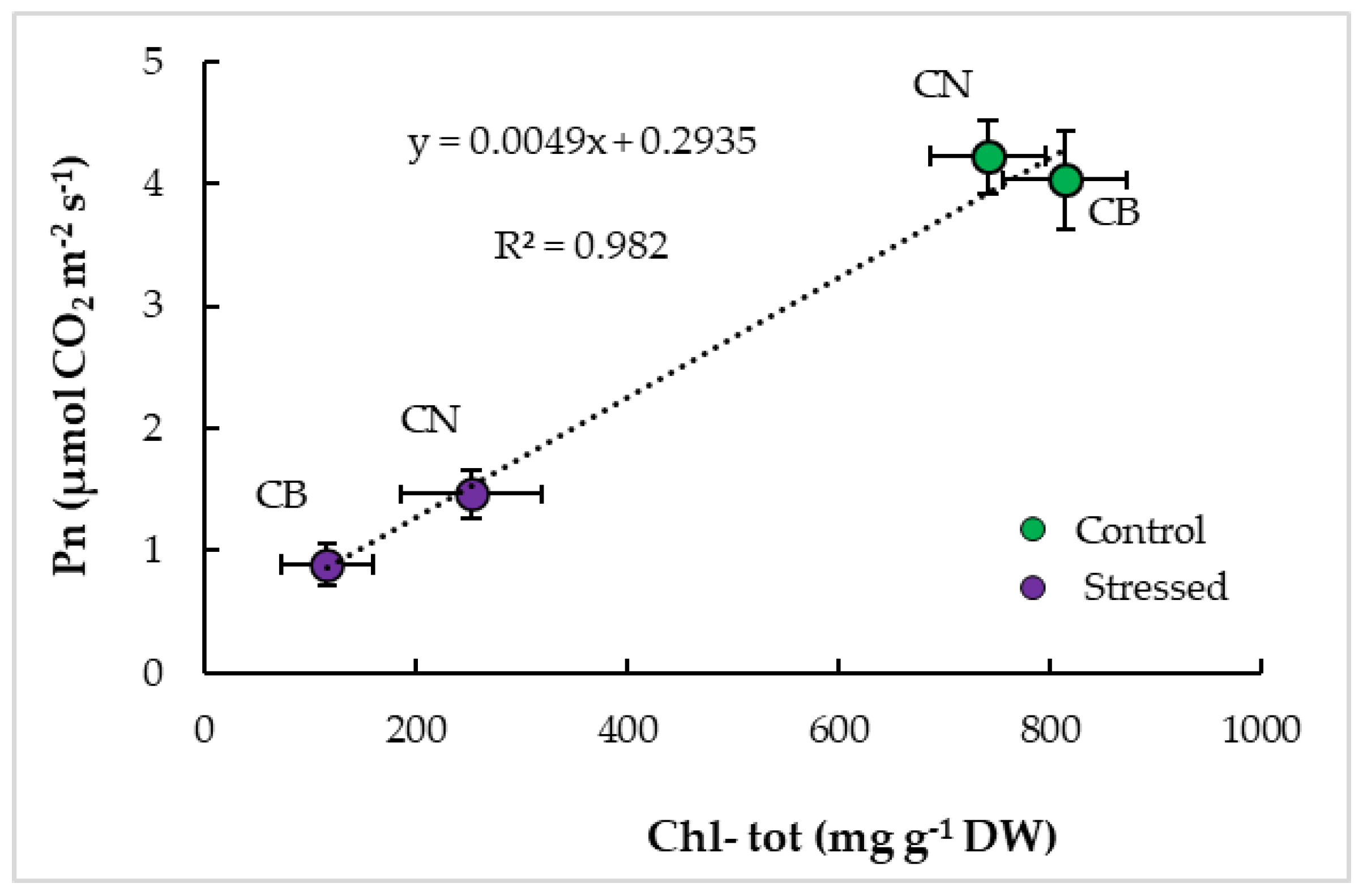
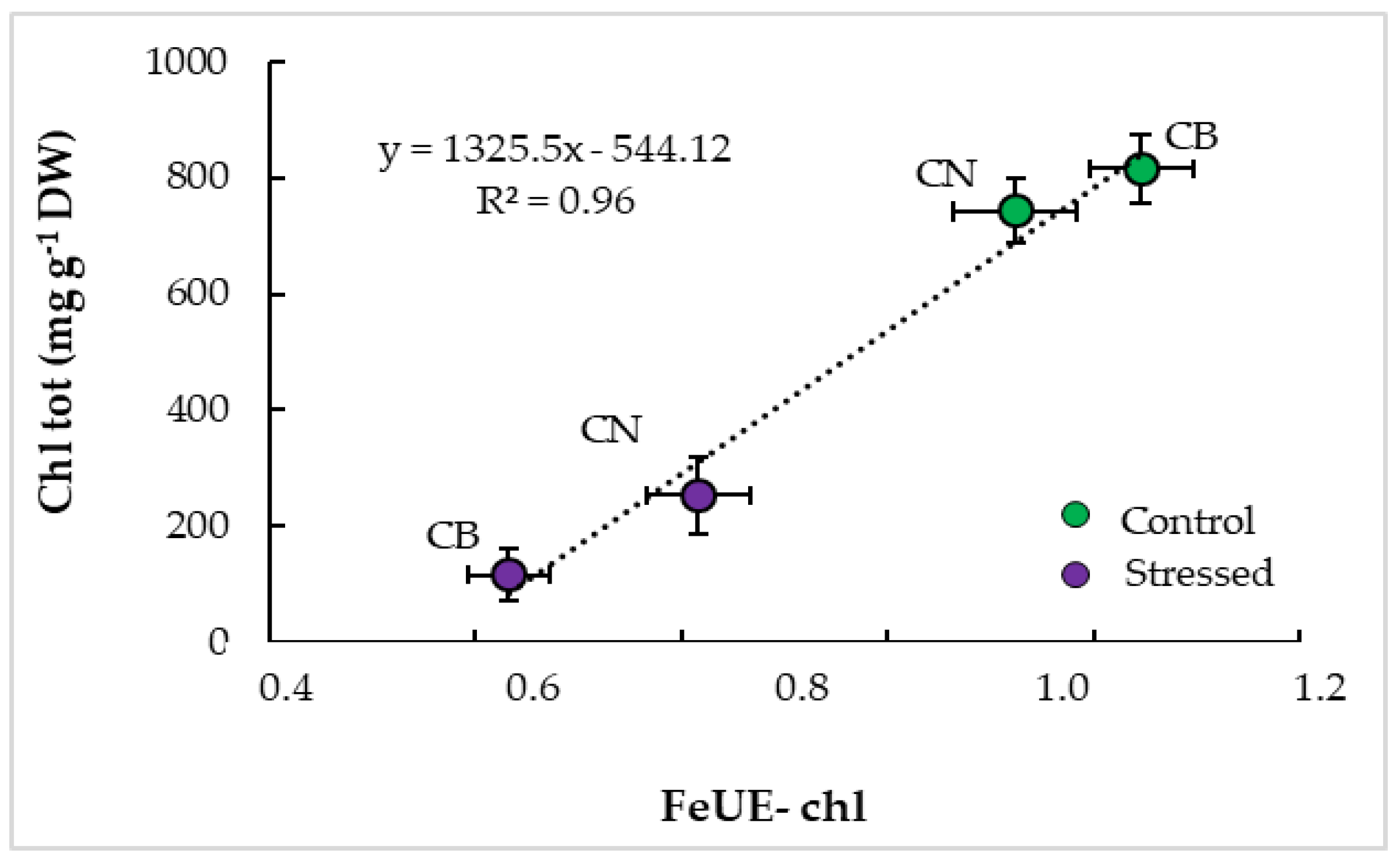
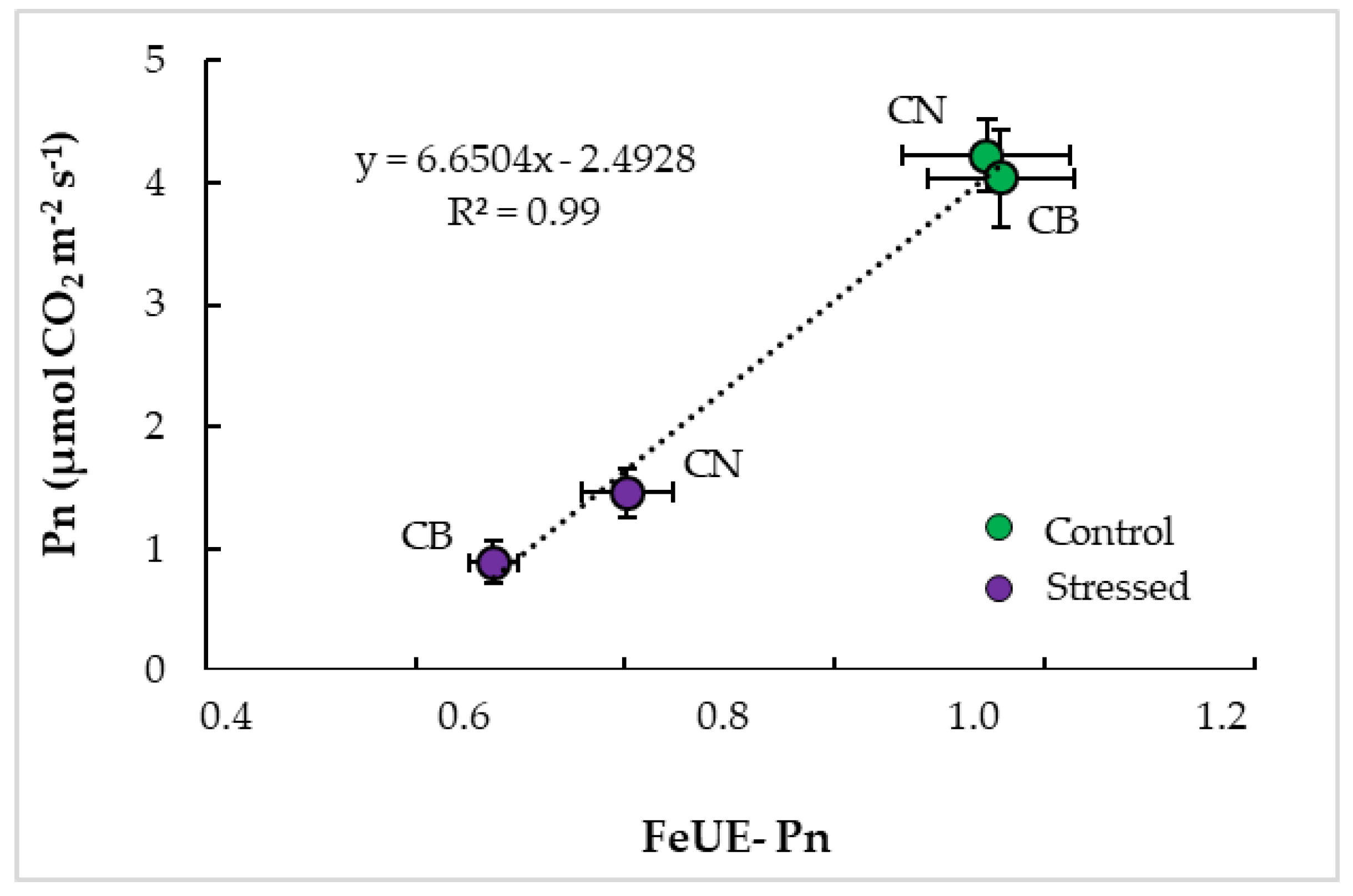
3.4. Interrelationships and Physiological Indicators
4. Discussion
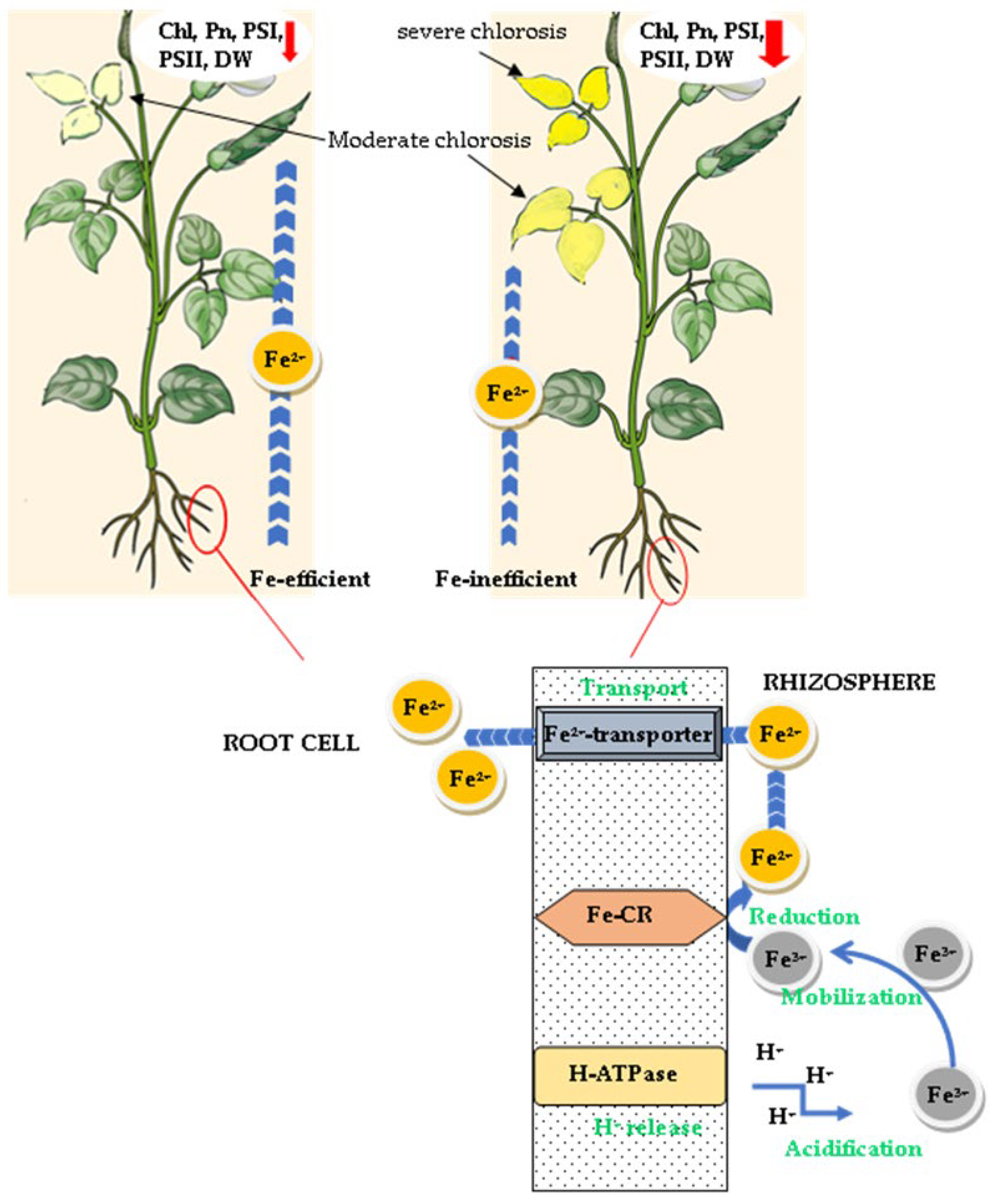
4.1. Plant Aspect and Physiological Response to Iron Deficiency
4.2. Iron Remobilization and Use Efficiency
4.3. Parameter Interrelationships and Tolerance Indicators
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lindsay, W.L. Chemical Equilibria in Soils; The Blackburn Press: Caldwell, NJ, USA, 2001; ISBN 1930665113/978-1930665118. [Google Scholar]
- Khan, S.; Akhtar, N.; Rehman, S.-U.; Jamil, M. Iron Oxide Nanoparticles: Plant Response, Interaction, Phytotoxicity and Defense Mechanisms. In Nanomaterials and Nanocomposites Exposures to Plants; Husen, A., Ed.; Springer: Berlin/Heidelberg, Germany, 2023; pp. 227–245. [Google Scholar] [CrossRef]
- Valipour, M.; Baninasab, B.; Khoshgoftarmanesh, A.-H.; Gholami, M. Oxidative stress and antioxidant responses to direct and bicarbonate induced iron deficiency in two quince rootstocks. Sci. Hort. 2020, 261, 108933. [Google Scholar] [CrossRef]
- Hantzis, L.J.; Kroh, G.-E.; Jahn, C.-E.; Cantrell, M.; Peers, G.; Pilon, M.; Et Ravet, K. Un programme d’économie de fer pendant la carence cible des protéines Fe spécifiques. Physiol. Vég. 2018, 176, 596–610. [Google Scholar] [CrossRef]
- Wang, J.; Pantopoulos, K. Regulation of cellular iron metabolism. Biochem. J. 2011, 434, 365–381. [Google Scholar] [CrossRef] [PubMed]
- Hochmuth, G. Iron (Fe) Nutrition of Plants; This Document Is SL353, One of a Series of the Soil and Water Science Department, Florida Cooperative Extension Service; Institute of Food and Agricultural Sciences, University of Florida: Gainesville, FL, USA, 2011; Available online: https://edis.ifas.ufl.edu (accessed on 13 August 2023).
- Wu, J.; Zhao, Y.; Qi, H.; Zhao, X.; Yang, T.; Du, Y.; Zhang, H.; Wei, Z. Identifying the key factors that affect the formation of humic substance during different materials composting. Biores. Technol. 2017, 244, 1193–1196. [Google Scholar] [CrossRef] [PubMed]
- Krouma, A. The Physiological Mechanisms Underlying N2-Fixing Common Bean (Phaseolus vulgaris L.) Tolerance to Iron Deficiency. Phyton Int. J. Exp. Bot. 2023, 92, 2133–2150. [Google Scholar] [CrossRef]
- Valentinuzzi, F.; Pii, Y.; Carlo, P.; Roberto, T.; Fontanella, M.C.; Beone, G.M.; Astolfi, S.; Mimmo, T.; Cesco, S. Root-shoot- root Fe translocation in cucumber plants grown in a heterogeneous Fe provision. Plant Sci. 2020, 293, 110431. [Google Scholar] [CrossRef] [PubMed]
- Krouma, A. Differential response of pea (Pisum sativum L.) genotypes to iron deficiency in relation to the growth, rhizosphere acidification and ferric chelate reductase activities. Aust. J. Crop Sci. 2021, 15, 925–932. [Google Scholar] [CrossRef]
- En-Jung, H.; Brian, M.-W. Alkaline stress and iron deficiency regulate iron uptake and riboflavin synthesis gene expression differently in root and leaf tissue: Implications for iron deficiency chlorosis. J. Exp. Bot. 2016, 67, 5671–5685. [Google Scholar] [CrossRef]
- Ceballos-Laita, L.; Takahashi, D.; Uemura, M.; Abadía, J.; López-Millán, A.-L.; Rodríguez-Celma, J. Effects of Fe and Mn Deficiencies on the Root Protein Profiles of Tomato (Solanum lycopersicum) Using Two-Dimensional Electrophoresis and Label-Free Shotgun Analyses. Int. J. Mol. Sci. 2022, 23, 3719. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.-L.; Clode, P.-L.; Lambers, H. Effects of pH and bicarbonate on the nutrient status and growth of three Lupinus species. Plant Soil 2020, 447, 9–28. [Google Scholar] [CrossRef]
- Brumbarova, T.; Bauer, P.; Ivanov, R. Molecular mechanisms governing Arabidopsis iron uptake. Trends Plant Sci. 2015, 20, 124–133. [Google Scholar] [CrossRef]
- Slatni, T.; Krouma, A.; Gouia, H.; Abdelly, C. Importance of ferric-chelate reductase activity and acidification capacity in root nodules of N2-fixing common bean (Phaseolus vulgaris L.) subjected to iron deficiency. Symbiosis 2009, 47, 35–42. [Google Scholar] [CrossRef]
- Merry, R.; Dobbels, A.A.; Sadol, W.; Naeve, S.; Stupar, R.-M.; Lorenz, A.J. Iron deficiency in soybean. Crop Sci. 2022, 62, 136–152. [Google Scholar] [CrossRef]
- Li, Q.; Chen, L.; Yang, A. The molecular mechanisms underlying iron deficiency responses in rice. Int. J. Mol. Sci. 2020, 21, 43. [Google Scholar] [CrossRef]
- Valentinuzzi, F.; Venuti, S.; Pii, Y.; Marroni, F.; Cesco, S.; Hartmann, F.; Mimmo, T.; Morgante, M.; Pinton, R.; Tomasi, N.; et al. Common and specific responses to iron and phosphorus deficiencies in roots of apple tree (Malus × domestica). Plant Mol. Biol. 2019, 101, 129–148. [Google Scholar] [CrossRef]
- Zamboni, A.; Zanin, L.; Tomasi, N.; Pezzotti, M.; Pinton, R.; Varanini, Z.; Cesco, S. Genome-wide microarray analysis of tomato roots showed defined responses to iron deficiency. BMC Genom. 2012, 13, 101. [Google Scholar] [CrossRef] [PubMed]
- Celletti, S.; Paolacci, A.-R.; Mimmo, T.; Pii, Y.; Cesco, S.; Ciaffi, M.; Astolfi, S. The effect of excess Sulphate supply on iron accumulation in three graminaceous plants at the early vegetative phase. Environ. Exp. Bot. 2016, 128, 31–38. [Google Scholar] [CrossRef]
- Kosegarten, H.; Hoffmann, B.; Rroco, E.; Grolig, F.; Glusenkamp, K.-H.; Mengel, K. Apoplastic pH and Fe-III reduction in young sunflower Helianthus annuus roots. Physiol. Plant. 2004, 122, 95–106. [Google Scholar] [CrossRef]
- Karimi, H.-R.; Tari, F.-E. Effects of NaHCO3 on photosynthetic characteristics, and iron and sodium transfer in pomegranate. J. Plant Nutr. 2017, 401, 1–22. [Google Scholar] [CrossRef]
- Briat, J.-F.; Fobis-Loisy, I.; Grignon, N.; Lobreaux, S.; Pascal, N.; Savino, G.; Thoiron, S.; Wiren, N.; Wuytswinkel, O. Cellular and molecular aspects of iron metabolism in plants. Biol. Cell 1995, 84, 69–81. [Google Scholar] [CrossRef]
- James, M.-C.; Janneke, B.; Jorge, R.-C. Iron homeostasis in plants—A brief overview. Metallomics 2017, 9, 813–823. [Google Scholar] [CrossRef]
- Salhi, K.; Hajlaoui, H.; Krouma, A. Genotypic differences in response of durum wheat (Triticum durum Desf.) to lime-induced iron chlorosis. Plant Direct 2022, 6, e377. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, R.; Brumbarova, T.; Bauer, P. Fitting into the harsh reality: Regulation of iron-deficiency responses in dicotyledonous plants. Mol. Plants 2012, 5, 27–42. [Google Scholar] [CrossRef]
- Robinson, N.-J.; Procter, C.-M.; Connolly, E.-L.; Guerinot, M.-L. A ferric-chelate reductase for iron uptake from soils. Nature 1999, 397, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Vert, G.; Grotz, N.; Dedaldechamp, F.; Gaymard, F.; Guerinot, M.-L.; Briat, J.-F.; Curie, C. IRT1, an Arabidopsis transporter essential for iron uptake from the soil and for plant growth. Plant Cell 2002, 14, 1223–1233. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Madison, I.; Long, T.-A.; Williams, C.-M. Computational solutions for modeling and controlling plant response to abiotic stresses: A review with focus on iron deficiency. Curr. Opin. Plant Biol. 2020, 57, 8–15. [Google Scholar] [CrossRef]
- Hamrouni, L.; Hanana, M.; Abdelly, C.; Ghorbel, A. Chloride exclusion and sodium inclusion: Two concomitant mechanisms of salinity tolerance in wild vine Vitis vinifera subsp. sylvestris (var. #x2018;Séjnène’). Biotechnol. Agron. Soc. Environ. 2011, 15, 387–400. [Google Scholar]
- Hewitt, E.-J. Sand and Water Culture Methods Used in the Study of Plant Nutrition, 2nd ed.; Commonwealth Bureau of Horticulture Technical Communications: Farnham Royal, UK, 1966; p. 22. [Google Scholar]
- Tang, C.; Robson, A.; Dilworth, M.-I. The role of iron in nodulation and nitrogen fixation in Lupinus angustifolius L. New Phytol. 1990, 114, 173–182. [Google Scholar] [CrossRef]
- Morales, F.; Abadia, A.; Abadia, J. Characterization of the xanthophyll cycle and other photosynthetic pigment changes induced by iron deficiency in sugar beet (Beta vulgaris L.). Plant Physiol. 1990, 94, 607–613. [Google Scholar] [CrossRef]
- Krouma, A.; Gharsalli, M.; Abdelly, C. Differences in response to iron deficiency among some lines of common bean. J. Plant Nutr. 2003, 26, 2295–2305. [Google Scholar] [CrossRef]
- Arnon, D. Copper enzymes in isolated chloroplast. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Köseoglu, A.-T.; Açikgöz, V. Determination of iron chlorosis with extractable iron analysis in peach leaves. J. Plant Nutr. 1995, 18, 153–161. [Google Scholar] [CrossRef]
- Ferhi, J.; Gharsalli, M.; Abdelly, C.; Krouma, A. Potential of the physiological response of pea plants (Pisum sativum L.) to iron deficiency (direct or lime-induced). Biosci. J. 2017, 33, 1208–1218. [Google Scholar] [CrossRef]
- Barhoumi, S.; Ellouzi, H.; Krouma, A. Functional Analysis of the Genotypic Differences in Response of Pea (Pisum sativum L.) to Calcareous-Induced Iron Deficiency. Phyton Int. J. Exp. Bot. 2023, 92, 522–536. [Google Scholar] [CrossRef]
- Thoiron, S.; Pascal, N.; Briat, J.F. Impact of iron deficiency and iron re-supply during the early stages of vegetative development in maize (Zea mays L.). Plant Cell Environ. 1997, 20, 1051–1060. [Google Scholar] [CrossRef]
- Athanassios, N.M.; Grigorios, C.D.; Ioannis, N.T.; Vassilios, T.; Kortessa, N.D. Oxidative stress, antioxidant activity and Fe(III)-chelate reductase activity of five Prunus rootstocks explants in response to Fe deficiency. Plant Growth Regul. 2005, 46, 69–78. [Google Scholar] [CrossRef]
- Fageria, N.-K.; Barbosa Filho, M.-P.; Moreira, A.; Guimarães, C.-M. Foliar Fertilization of Crop Plants. Plant Nutr. 2009, 32, 1044–1064. [Google Scholar] [CrossRef]
- Ye, L.; Li, L.; Wang, L.; Wang, S.; Li, S.; Du, J.; Shou, H. MPK3/MPK6 are involved in iron deficiency-induced ethylene production in Arabidopsis. Front. Plant Sci. 2015, 6, 953. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nishizawa, N.K. Iron uptake, translocation, and regulation in higher plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef]
- Zargar, S.M.; Agrawal, G.K.; Rakwal, R.; Fukao, Y. Quantitative proteomics reveals role of sugar in decreasing photosynthetic activity due to Fe deficiency. Front. Plant Sci. 2015, 6, 592. [Google Scholar] [CrossRef]
- Rutherford, A.-W. Orientation of EPR signals arising from components in photosystem II membranes. Biochim. Biophys. Acta 1985, 807, 189–201. [Google Scholar] [CrossRef]
- Sandmann, G. Consequences of iron deficiency on photosynthetic and respiratory electron transport in blue-green algae. Photosynth. Res. 1985, 6, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Spence, M.J.; Henzl, M.T.; Lammers, P.J. The structure of a Phaseolus vulgaris cDNA encoding the iron storage protein ferritin. Plant Mol. Biol. 1991, 117, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Briat, J.-F.; Dubos, C.; Gaymard, F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 2015, 20, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Nozoye, T.; Otani, M.; Senoura, T.; Nakanishi, H.; Nishizawa, N.-K. Overexpression of barley nicotianamine synthase 1 confers tolerance in the sweet potato to iron deficiency in calcareous soil. Plant Soil 2016, 418, 75–88. [Google Scholar] [CrossRef]
- Bisht, S.S.; Nautiyal, B.D.; Sharma, C.P. Biochemical changes under iron deficiency and recovery in tomato. Ind. J. Plant Physiol. 2002, 7, 183–186. [Google Scholar]
- López-Millán, A.F.; Morales, F.; Abadía, A.; Abadía, J. Iron deficiency-associated changes in the composition of the leaf apoplastic fluid from field-grown pear (Pyrus communis L.) Trees. J. Exp. Bot. 2001, 52, 1489–1496. [Google Scholar] [CrossRef]
- Smith, B.-N. Iron in higher plants: Storage and metabolic role. J. Plant Nutr. 1984, 7, 759–766. [Google Scholar] [CrossRef]
- Briat, J.F. Iron homeostasis in plants. Cah. Agric. 2004, 183–201. [Google Scholar]
- Stocking, C.-R. Iron deficiency and the structure and physiology of maize chloroplasts. Plant Physiol. 1975, 55, 626–631. [Google Scholar] [CrossRef]
- Sharma, S.; Sanwal, G.-G. Effect of iron deficiency on the photosynthetic system of maize. Plant Physiol. 1992, 140, 527–530. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, M.; Liang, Z.; Li, H.; Yu, F.; Liu, S. Analysis of contrast iron chlorosis tolerance in the pear cv. ‘Huangguan’ grafted onto Pyrus betulifolia and quince A grown in calcareous soils. Sci. Hort. 2020, 271, 109488. [Google Scholar] [CrossRef]
- Kumar, O.; Sridevi, O.; Naidu, G.K.; Patil, B.-C. Association between Productivity Parameters and Iron Deficiency Chlorosis in Groundnut (Arachis hypogaea L.). Pharma Innov. J. 2022, 11, 1123–1127. [Google Scholar]
- Mehrotra, R.; Verma, R.-K.; Pal, A. Iron deficiency chlorosis in aromatic grasses—A review. Environ. Chall. 2022, 9, 100646. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CN | CB | |||
|---|---|---|---|---|
| Control | Stressed | Control | Stressed | |
| Chl-a (mg g−1 FW) | 510 ± 32 b | 179 ± 15.2 c | 574 ± 41.1 a | 99 ± 8.4 d |
| Chl-b (mg g−1 FW) | 233 ± 18.5 b | 74 ± 6.3 c | 342 ± 30.4 a | 54 ± 4.2 d |
| Chl-tot (mg g−1 FW) | 742 ± 33.2 b | 253 ± 18.8 c | 815 ± 51.2 a | 116 ± 8.9 d |
| spad index | 34.57 ± 3.3 a | 27.31 ± 2.5 b | 31.81 ± 2.5 ab | 17.19 ± 1.5 c |
| DW (g plant−1) | 683 ± 53.3 a | 543 ± 32.4 c | 581 ± 41.5 b | 334 ± 22.6 d |
| CN | CB | |||
|---|---|---|---|---|
| Control | Stressed | Control | Stressed | |
| FeQ-Sh | 1505 ± 48.5 a | 873 ± 36.6 c | 1375 ± 58.5 b | 642 ± 33.7 d |
| FeQ-R | 2624 ± 101.1 a | 807 ± 43.4 b | 2559 ± 121.3 ab | 686 ± 44.6 c |
| FeQ-tot | 4128 ± 125.3 a | 168 ± 88.6 c | 3933 ± 142.4 b | 1328 ± 78.4 d |
| FeT (%) | 36 ± 2.7 b | 52 ± 4.5 a | 35 ± 2.9 b | 48 ± 3.8 a |
| RAC | Fe-CRC | |||
|---|---|---|---|---|
| Time (mn) | CB | CN | CB | CN |
| 30 | −0.11 ± 0.01 f | −0.44 ± 0.04 e | 25 ± 2.2 h | 30 ± 2.6 g |
| 60 | −0.74 ± 0.06 de | −0.97 ± 0.07 c | 69 ± 5.3 f | 88 ± 6.4 ef |
| 90 | −0.73 ± 0.05 de | −1.27 ± 0.12 bc | 95 ± 6.9 e | 133 ± 10.2 b |
| 120 | −0.79 ± 0.06 d | −1.48 ± 0.14 a | 111 ± 8.4 d | 146 ± 10.5 a |
| 150 | −0.74 ± 0.05 de | −1.31 ± 0.11 b | 105 ± 9.0 de | 122 ± 9.7 c |
| Mb-E | CB | CN | ||
|---|---|---|---|---|
| Time (mn) | Control | Stressed | Control | Stressed |
| 30 | 3.1 ± 0.33 l | 7.2 ± 0.58 h | 3.1 ± 0.30 l | 8.4 ± 0.62 g |
| 60 | 3.1 ± 0.25 l | 14.7 ± 1.44 f | 3.3 ± 0.27 l | 19.1 ± 1.7 e |
| 90 | 3.7 ± 0.33 kl | 20.2 ± 1.8 e | 4.1 ± 0.35 k | 30.4 ± 2.6 b |
| 120 | 6.0 ± 0.51 i | 26.3 ± 2.2 c | 8.5 ± 0.66 g | 41.0 ± 3.7 a |
| 150 | 5.2 ± 0.48 j | 23.0 ± 2.1 d | 6.6 ± 0.49 hi | 31.1 ± 2.9 b |
| CN | CB | |||
|---|---|---|---|---|
| Control | Stressed | Control | Stressed | |
| FeUE-DW | 0.67 ± 0.063 b | 0.89 ± 0.072 a | 0.60 ± 0.054 c | 0.67 ± 0.057 b |
| FeUE-Pn | 4.13 ± 0.35 a | 2.41 ± 0.18 b | 4.19 ± 0.38 a | 1.77 ± 0.15 c |
| FeUE-chl | 0.72 ± 0.066 b | 0.42 ± 0.033 c | 0.85 ± 0.068 a | 0.23 ± 0.020 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nsiri, K.; Krouma, A. The Key Physiological and Biochemical Traits Underlying Common Bean (Phaseolus vulgaris L.) Response to Iron Deficiency, and Related Interrelationships. Agronomy 2023, 13, 2148. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13082148
Nsiri K, Krouma A. The Key Physiological and Biochemical Traits Underlying Common Bean (Phaseolus vulgaris L.) Response to Iron Deficiency, and Related Interrelationships. Agronomy. 2023; 13(8):2148. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13082148
Chicago/Turabian StyleNsiri, Khawla, and Abdelmajid Krouma. 2023. "The Key Physiological and Biochemical Traits Underlying Common Bean (Phaseolus vulgaris L.) Response to Iron Deficiency, and Related Interrelationships" Agronomy 13, no. 8: 2148. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy13082148