
The Combined Analysis of the Transcriptome and Metabolome Revealed the Possible Mechanism of Flower Bud Formation in Amorphophallus bulbifer

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material Cultivation
2.2. Transcriptome Analysis
2.2.1. RNA Extraction from Flower Buds and Leaf Buds of A. bulbifer
2.2.2. Transcriptome Sequencing Analysis
2.2.3. Quantitative RT–PCR (qRT–PCR) Analysis
2.3. Determination of Sugar Content in A. bulbifer Corms
2.4. Targeted Metabolic Analysis of A. bulbifer Flower Buds and Leaf Bud Hormone Content
2.4.1. Sample Extraction
2.4.2. Liquid-Phase Parameters
2.4.3. Mass Spectrometry Parameters
2.5. Nontargeted Metabolic Analysis of A. bulbifer Flower Bud and Leaf Bud Metabolite Contents
2.5.1. Metabolite Extraction
2.5.2. UPLC–MS Analysis
2.5.3. Data Processing and Analysis
2.6. Combined Transcriptomic and Metabolomic Analysis
3. Results
3.1. Morphological Observation of Flower Buds and Leaf Buds of A. bulbifer
3.2. Transcriptome Analysis
3.2.1. Transcriptome Sequencing Data Processing, Transcript Splicing and Functional Annotation
3.2.2. Screening of Differentially Expressed Genes
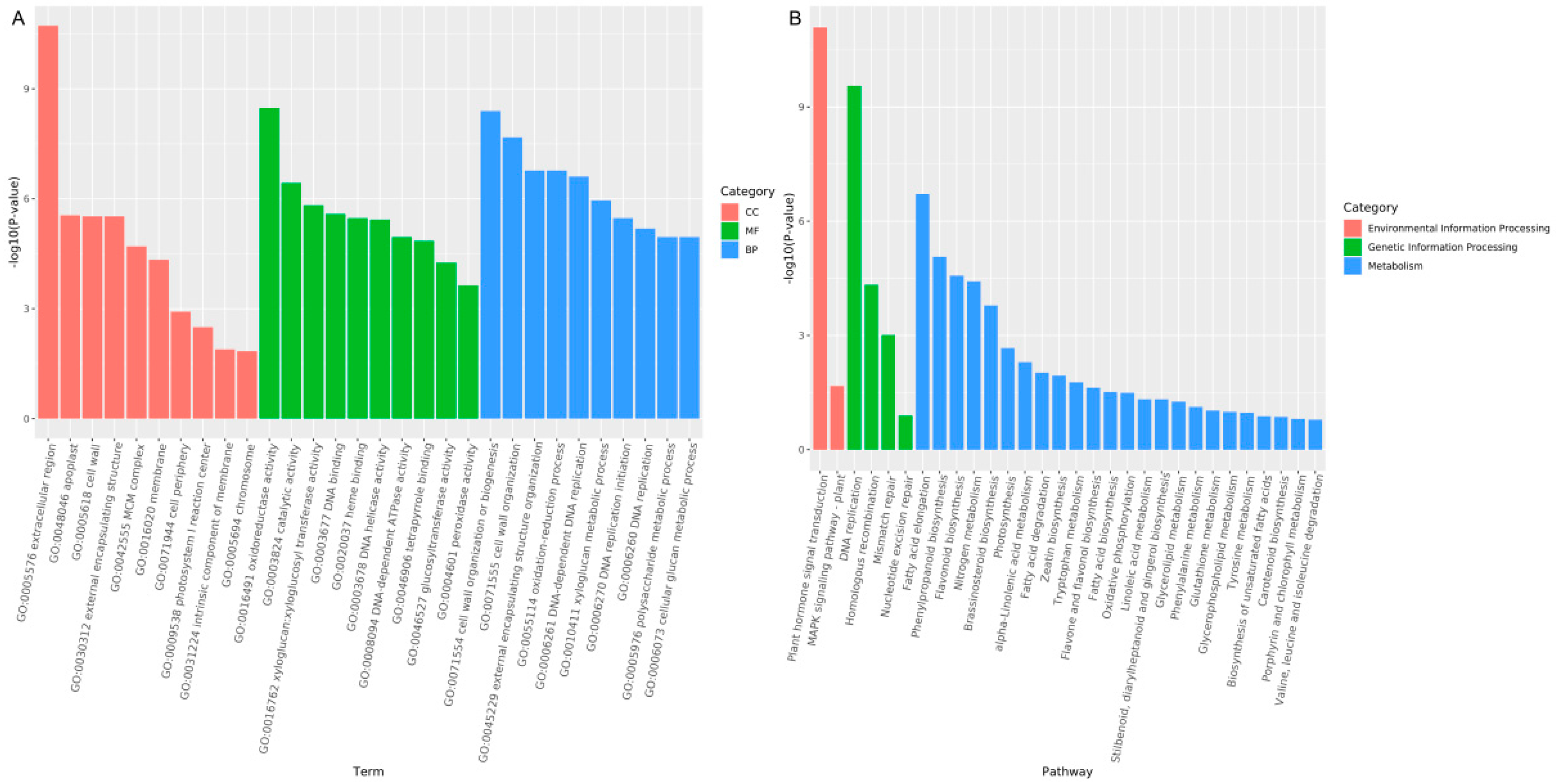
3.2.3. Functional Annotation and Classification of Differentially Expressed Genes
3.2.4. Validation of the Transcriptome Data
3.3. Determination of Total Soluble Sugar and Starch Content in A. bulbifer Corms
3.4. Metabolomic Analysis
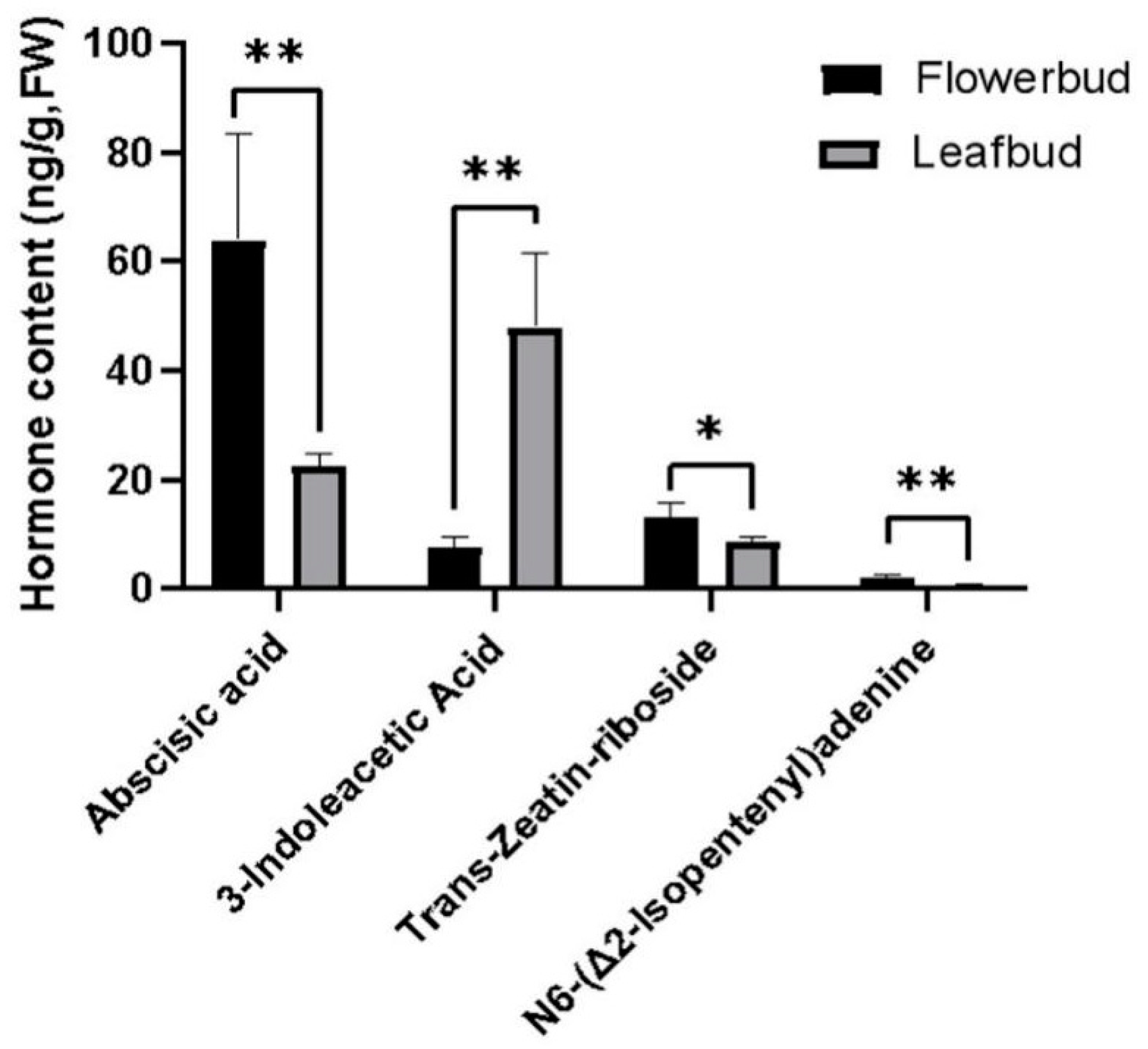
3.4.1. Targeted Metabolic Analysis of Endogenous Hormone Contents in Flower Buds and Leaf Buds of A. bulbifer with Corms
3.4.2. Nontargeted Analysis of the Metabolite Content in Flower Buds and Leaf Buds of A. bulbifer
3.4.3. Identification of Differentially Accumulated Metabolites
3.5. Correlation Analysis between Differentially Expressed Genes and Differentially Accumulated Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jie, P. A Study on Chinese Amorphophallus Resources. Resour. Sci. 2001, 23, 87–89. [Google Scholar]
- Liu, P.Y. Konjac; China Agricultural Press: Beijing, China, 2004; p. 348. [Google Scholar]
- Choi, K.H.; Kim, S.T.; Bin, B.H.; Park, P.J. Effect of Konjac Glucomannan (KGM) on the Reconstitution of the Dermal Environment against UVB-Induced Condition. Nutrients 2020, 12, 2779. [Google Scholar] [CrossRef]
- Si, Y.; Liu, X.; Ye, K.; Bonfini, A.; Hu, X.Y.; Buchon, N.; Gu, Z. Glucomannan Hydrolysate Promotes Gut Proliferative Homeostasis and Extends Life Span in Drosophila melanogaster. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2019, 74, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Liu, J.; Ding, Y. Recent progress in biological activities and health benefits of konjac glucomannan and its derivatives. Bioact. Carbohydr. Diet. Fibre 2021, 26, 100270. [Google Scholar] [CrossRef]
- Zhao, C.; Harijati, N.; Liu, E.; Jin, S.; Diao, Y.; Hu, Z. First report on DNA content of three species of Amorphophallus. J. Genet. 2020, 99, 36. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, D.; Zhao, J.P.; Srzednicki, G.; Borompichaichartkul, C.; Kanlayanarat, S. Morphological and growth characteristics of Amorphophallus muelleri blume A commercially important konjac species. Acta Hortic. 2010, 875, 501–508. [Google Scholar] [CrossRef]
- Gao, H.; Zhao, Y.; Huang, L.H.; Huang, Y.; Chen, J.J.; Zhou, H.Y.; Zhang, X.W. Comparative analysis of buds transcriptome and identification of two florigen gene AkFTs in Amorphophallus konjac. Sci. Rep. 2022, 12, 6782. [Google Scholar] [CrossRef] [PubMed]
- Susila, H.; Nasim, Z.; Ahn, J.H. Ambient Temperature-Responsive Mechanisms Coordinate Regulation of Flowering Time. Int. J. Mol. Sci. 2018, 19, 3196. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.A.; Li, C.; Lin, H.; Joe, A.; Padilla, M.; Woods, D.P.; Dubcovsky, J. EARLY FLOWERING 3 interactions with PHYTOCHROME B and PHOTOPERIOD1 are critical for the photoperiodic regulation of wheat heading time. PLoS Genet. 2023, 19, e1010655. [Google Scholar] [CrossRef]
- Gai, Z.; Zhang, M.; Zhang, P.; Zhang, J.; Liu, J.; Cai, L.; Yang, X.; Zhang, N.; Yan, Z.; Liu, L.; et al. 2-Oxoglutarate contributes to the effect of foliar nitrogen on enhancing drought tolerance during flowering and grain yield of soybean. Sci. Rep. 2023, 13, 7274. [Google Scholar] [CrossRef]
- Yu, H.; Ma, H.; Fang, X.; Lai, W.; Yu, S. Review on Mechanism of Strawberry Flower Bud Differentiation and Application of Regulation Techniques. Acta Agric. Jiangxi 2011, 23, 58–61. [Google Scholar]
- Lu, X.Y.; Li, J.J.; Chen, H.B.; Hu, J.Q.; Liu, P.X.; Zhou, B.Y. RNA-seq analysis of apical meristem reveals integrative regulatory network of ROS and chilling potentially related to flowering in Litchi chinensis. Sci. Rep. 2017, 7, 10183. [Google Scholar] [CrossRef]
- Wang, Q.; Zuo, Z.; Wang, X.; Liu, Q.; Gu, L.; Oka, Y.; Lin, C. Beyond the photocycle-how cryptochromes regulate photoresponses in plants? Curr. Opin. Plant Biol. 2018, 45 Pt A, 120–126. [Google Scholar] [CrossRef]
- Gao, D.; Ji, X.; Yuan, Q.; Pei, W.; Zhang, X.; Li, F.; Han, Q.; Zhang, S. Effects of total daily light integral from blue and broad-band red LEDs on flowering of saffron (Crocus sativus L.). Sci. Rep. 2023, 13, 7175. [Google Scholar] [CrossRef]
- Li, J.; Li, G.; Wang, H.; Wang Deng, X. Phytochrome signaling mechanisms. Arab. Book 2011, 9, e0148. [Google Scholar] [CrossRef]
- Woods, D.P.; Li, W.; Sibout, R.; Shao, M.; Laudencia-Chingcuanco, D.; Vogel, J.P.; Dubcovsky, J.; Amasino, R.M. PHYTOCHROME C regulation of photoperiodic flowering via PHOTOPERIOD1 is mediated by EARLY FLOWERING 3 in Brachypodium distachyon. PLoS Genet. 2023, 19, e1010706. [Google Scholar] [CrossRef]
- Lin, C. Photoreceptors and Regulation of Flowering Time1. Plant Physiol. 2000, 123, 39–50. [Google Scholar] [CrossRef]
- Lee, J.; Lee, I. Regulation and function of SOC1, a flowering pathway integrator. J. Exp. Bot. 2010, 61, 2247–2254. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Ying, H.; Helliwell, C.A.; Taylor, J.M.; Peacock, W.J.; Dennis, E.S. FLOWERING LOCUS C (FLC) regulates development pathways throughout the life cycle of Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 6680–6685. [Google Scholar] [CrossRef] [PubMed]
- Lastdrager, J.; Hanson, J.; Smeekens, S. Sugar signals and the control of plant growth and development. J. Exp. Bot. 2014, 65, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Mesejo, C.; Martinez-Fuentes, A.; Reig, C.; Agusti, M. The flower to fruit transition in Citrus is partially sustained by autonomous carbohydrate synthesis in the ovary. Plant Sci. 2019, 285, 224–229. [Google Scholar] [CrossRef]
- Gu, J.H.; Zeng, Z.; Wang, Y.R.; Lyu, Y.M. Transcriptome Analysis of Carbohydrate Metabolism Genes and Molecular Regulation of Sucrose Transport Gene LoSUT on the Flowering Process of Developing Oriental Hybrid Lily ‘Sorbonne’ Bulb. Int. J. Mol. Sci. 2020, 21, 3092. [Google Scholar] [CrossRef]
- Yue, Z.; Wang, Y.; Zhang, N.; Zhang, B.; Niu, Y. Expression of the Amorphophallus albus heat stress transcription factor AaHsfA1 enhances tolerance to environmental stresses in Arabidopsis. Ind. Crops Prod. 2021, 174, 114231. [Google Scholar] [CrossRef]
- Liu, T.; Hu, Y.Q.; Li, X.X. Comparison of dynamic changes in endogenous hormones and sugars between abnormal and normal Castanea mollissima. Prog. Nat. Sci.-Mater. Int. 2008, 18, 685–690. [Google Scholar] [CrossRef]
- Yan, B.B.; Hou, J.L.; Cui, J.; He, C.; Li, W.B.; Chen, X.Y.; Li, M.; Wang, W.Q. The Effects of Endogenous Hormones on the Flowering and Fruiting of Glycyrrhiza uralensis. Plants 2019, 8, 519. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Chen, Y.; Yang, W.; Li, M.; Sun, B.; Song, H.; Tang, W.; Zhang, Y.; Gong, R. Joint metabolome and transcriptome analysis of the effects of exogenous GA (3) on endogenous hormones in sweet cherry and mining of potential regulatory genes. Front. Plant Sci. 2022, 13, 1041068. [Google Scholar] [CrossRef] [PubMed]
- Mi, L.; Ma, D.; Lv, S.; Xu, S.; Zhong, B.; Peng, T.; Liu, D.; Liu, Y. Comparative Transcriptome and sRNAome Analyses Reveal the Regulatory Mechanisms of Fruit Ripening in a Spontaneous Early-Ripening Navel Orange Mutant and Its Wild Type. Genes 2022, 13, 1706. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Liu, B.; Ge, L.; Weng, Y.; Cao, X.; Liu, F.; Mao, P.; Ma, X. Identification and characterization of regulatory pathways involved in early flowering in the new leaves of alfalfa (Medicago sativa L.) by transcriptome analysis. BMC Plant Biol. 2021, 21, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.X.; Luo, T.; Zhang, H.; Shao, J.Z.; Peng, J.Y.; Sun, J.S. Variation of Endogenous Hormones during Flower and Leaf Buds Development in ‘Tianhong 2’ Apple. Hortscience 2020, 55, 1794–1798. [Google Scholar] [CrossRef]
- Liang, Y.; Bai, J.; Xie, Z.; Lian, Z.; Guo, J.; Zhao, F.; Liang, Y.; Huo, H.; Gong, H. Tomato sucrose transporter SlSUT4 participates in flowering regulation by modulating gibberellin biosynthesis. Plant Physiol. 2023, 192, 1080–1098. [Google Scholar] [CrossRef]
- Wu, L.; Ma, N.; Jia, Y.; Zhang, Y.; Feng, M.; Jiang, C.Z.; Ma, C.; Gao, J. An Ethylene-Induced Regulatory Module Delays Flower Senescence by Regulating Cytokinin Content. Plant Physiol. 2016, 173, 853–862. [Google Scholar] [CrossRef]
- Wu, Z.; Jiang, Z.; Li, Z.; Jiao, P.; Zhai, J.; Liu, S.; Han, X.; Zhang, S.; Sun, J.; Gai, Z.; et al. Multi-omics analysis reveals spatiotemporal regulation and function of heteromorphic leaves in Populus. Plant Physiol. 2023, 192, 188–204. [Google Scholar] [CrossRef]
- Tan, C.; Li, S.; Qian, C.; Wang, H. Regulatory effects of polysaccharides and hormones on multi-leaf formation of Amorphophallus bulbifer. J. Northwest A F Univ. 2022, 50, 69–79. [Google Scholar]
- Xiao, M.; Zhang, Y.; Chen, X.; Lee, E.J.; Barber, C.J.S.; Chakrabarty, R.; Desgagné Penix, I.; Haslam, T.M.; Kim, Y.B.; Liu, E.; et al. Transcriptome analysis based on next-generation sequencing of non-model plants producing specialized metabolites of biotechnological interest. J. Biotechnol. 2013, 166, 122–134. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, A.H.; Miao, J.H.; Sun, H.; Han, Y.; Yan, G.L.; Wu, F.F.; Wang, X.J. Metabolomics biotechnology, applications, and future trends: A systematic review. RSC Adv. 2019, 9, 37245–37257. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR andthe 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Mcgrance, S.J.; Cornell, H.J.; Rix, C.J. A Simple and Rapid Colorimetric Method for the Determination of Amylose in Starch Products. Starch-Stärke 1998, 50, 158–163. [Google Scholar] [CrossRef]
- Šimura, J.; Antoniadi, I.; Široká, J.; Tarkowská, D.E.; Strnad, M.; Ljung, K.; Novák, O. Plant Hormonomics: Multiple Phytohormone Profiling by Targeted Metabolomics. Plant Physiol. 2018, 177, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Glauser, G.; Grund, B.; Gassner, A.-L.; Menin, L.; Henry, H.; Bromirski, M.; Schütz, F.; Mcmullen, J.; Rochat, B. Validation of the Mass-Extraction-Window for Quantitative Methods Using Liquid Chromatography High Resolution Mass Spectrometry. Anal. Chem. 2016, 88, 3264–3271. [Google Scholar] [CrossRef]
- Vasilev, N.; Boccard, J.; Lang, G.; Grömping, U.; Fischer, R.; Goepfert, S.; Rudaz, S.; Schillberg, S. Structured plant metabolomics for the simultaneous exploration of multiple factors. Sci. Rep. 2016, 6, 37390. [Google Scholar] [CrossRef]
- Hesham, O.; Othman, E.; Refaat, J.; Yehia, S.; Pimentel-Elardo, S.; Nodwell, J.; Schirmeister, T.; Tawfike, A.; Abdelmohsen, U. Metabolomics analysis and biological investigation of three Malvaceae plants. Phytochem. Anal. 2020, 31, 204–214. [Google Scholar]
- Monnerat, G.; Seara, F.A.C.; Evaristo, J.A.M.; Carneiro, G.; Evaristo, G.P.C.; Domont, G.; Nascimento, J.H.M.; Mill, J.G.; Nogueira, F.C.S.; Campos De Carvalho, A.C. Aging-related compensated hypogonadism: Role of metabolomic analysis in physiopathological and therapeutic evaluation. J. Steroid Biochem. Mol. Biol. 2018, 183, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Want, E.J.; O’maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]
- Trygg, J.; Wold, S. Orthogonal Projections to Latent Structures (O-PLS). J. Chemom. 2002, 16, 119–128. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Coneva, V.; Zhu, T.; Colasanti, J. Expression differences between normal and indeterminate1 maize suggest downstream targets of ID1, a floral transition regulator in maize. J. Exp. Bot. 2007, 58, 3679–3693. [Google Scholar] [CrossRef]
- Goyal, A. Osmoregulation in Dunaliella, Part II: Photosynthesis and starch contribute carbon for glycerol synthesis during a salt stress in Dunaliella tertiolecta. Plant Physiol. Biochem. 2007, 45, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Nagao, M.; Minami, A.; Arakawa, K.; Fujikawa, S.; Takezawa, D. Rapid degradation of starch in chloroplasts and concomitant accumulation of soluble sugars associated with ABA-induced freezing tolerance in the moss Physcomitrella patens. J. Plant Physiol. 2005, 162, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Sami, F.; Yusuf, M.; Faizan, M.; Faraz, A.; Hayat, S. Role of sugars under abiotic stress. Plant Physiol. Biochem. 2016, 109, 54–61. [Google Scholar] [CrossRef]
- Lu, B.; Chen, L.; Hao, J.; Zhang, Y.; Huang, J. Comparative transcription profiles reveal that carbohydrates and hormone signalling pathways mediate flower induction in Juglans sigillata after girdling. Ind. Crop. Prod. 2020, 153, 112556. [Google Scholar] [CrossRef]
- Wei, L.; Yu, B.; Song, E. Responses of Endogenous Hormones and Amino Acids to Flower Bud Differentiation of Cassava. Southwest China J. Agric. Sci. 2021, 34, 1400–1406. [Google Scholar]
- Su, H.; Xu, K.; Liu, W. Changes of endogenous homones during the process of flower bud differentiation of Welsh onion. Acta Hortic. Sin. 2007, 34, 671–676. [Google Scholar]
- Liu, Z.; Zeng, L.; Du, X.; Peng, Y.; Tao, Y.; Li, Y.; Qin, J. Flower Bud Differentiation and Endogenous Hormone Changes of RosaAngela’. Bull. Bot. Res. 2021, 41, 37–43. [Google Scholar]
- Skogerbo, G. Effects of root pruning and trunk girdling on xylem cytokinin content of apple (Malus x domestica Borkh.). Norweigain J. Agric. Sci. 1992, 6, 499–527. [Google Scholar]
- Kojima, K.; Yamada, Y.; Yamamoto, M. Effects of Abscisic Acid Injection on Sugar and Organic Acid Contents of Citrus Fruit. J. Jpn. Soc. Hortic. Sci. 1995, 64, 17–21. [Google Scholar] [CrossRef]
- Ito, A.; Yaegaki, H.; Hayama, H.; Kusaba, S.-N.-S.; Yamaguchi, I.; Yoshioka, H.J.H. Bending Shoots Stimulates Flowering and Influences Hormone Levels in Lateral Buds of Japanese Pear. HortScience 1999, 34, 1224–1228. [Google Scholar] [CrossRef]
- Goldschmidt, E.E. Endogenous abscisic acid and 2-trans-abscisic acid in alternate bearing ‘Wilking’ mandarin trees. Plant Growth Regul. 1984, 2, 9–13. [Google Scholar] [CrossRef]
- Kieber, J.J.; Schaller, G.E. Cytokinin signaling in plant development. Development 2018, 145, dev149344. [Google Scholar] [CrossRef]
- Jameson, P.E.; Dhandapani, P.; Novak, O.; Song, J. Cytokinins and Expression of SWEET, SUT, CWINV and AAP Genes Increase as Pea Seeds Germinate. Int. J. Mol. Sci. 2016, 17, 2013. [Google Scholar] [CrossRef]
- Corbesier, L.; Prinsen, E.; Jacqmard, A.; Lejeune, P.; Van Onckelen, H.; Périlleux, C.; Bernier, G. Cytokinin levels in leaves, leaf exudate and shoot apical meristem of Arabidopsis thaliana during floral transition. J. Exp. Bot. 2003, 54, 2511–2517. [Google Scholar] [CrossRef]
- He, Y.W.; Loh, C.S. Induction of early bolting in Arabidopsis thaliana by triacontanol, cerium and lanthanum is correlated with increased endogenous concentration of isopentenyl adenosine (iPAdos). J. Exp. Bot. 2002, 53, 505–512. [Google Scholar] [CrossRef]
- Song, Y.; Dou, L.; Zhang, H. Changes of endogenous hormones during flower bud formation of different blueberry varieties. Fruit Trees South. China 2014, 43, 106–108+114. [Google Scholar]
- Du, J.; Li, P.; Gu, Y. Study on the changes of endogenous hormones and carbohydrate content during flower bud differentiation of olive. West. For. Sci. 2018, 47, 122–126. [Google Scholar]
- Ai, J.; Wang, Y.P.; Li, C.Y.; Guo, X.W.; Li, A.M. The changes of three endogenous hormones during flower bud differentiation of Schisandga chinensis. China J. Chin. Mater. Medica 2006, 31, 24–26. [Google Scholar]
- Kinet, J.M. Environmental, Chemical, and Genetic Control of Flowering. Hortic. Rev. 2010, 15, 279–334. [Google Scholar]
- Grochowska, M.J.; Hodun, M. The dwarfing effect of a single application of growth inhibitors to the root-stem connection “the collar tissue” of five species of fruit trees. J. Hortic. Sci. 1997, 72, 83–91. [Google Scholar] [CrossRef]
- Li, Y.; Xing, L.; Zhang, D.; Shen, Y.; Zhang, S.; Han, M. Mechanism of Spraying IAA Inhibiting Flower Bud Inoculation of Fuji Young Trees. Acta Agric. Boreali-Occi-Dent. Sin. 2015, 24, 84–89. [Google Scholar]
- Gomes, G.L.B.; Scortecci, K.C. Auxin and its role in plant development: Structure, signalling, regulation and response mechanisms. Plant Biol. 2021, 23, 894–904. [Google Scholar] [CrossRef] [PubMed]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 516. [Google Scholar] [CrossRef]
- Wen, Z.; Li, M.; Meng, J.; Miao, R.; Liu, X.; Fan, D.; Lv, W.; Cheng, T.; Zhang, Q.; Sun, L. Genome-Wide Identification of the MAPK and MAPKK Gene Families in Response to Cold Stress in Prunus mume. Int. J. Mol. Sci. 2023, 24, 8829. [Google Scholar] [CrossRef]
- Zhao, F.; Zheng, Y.F.; Zeng, T.; Sun, R.; Yang, J.Y.; Li, Y.; Ren, D.T.; Ma, H.; Xu, Z.H.; Bai, S.N. Phosphorylation of SPOROCYTELESS/NOZZLE by the MPK3/6 Kinase Is Required for Anther Development. Plant Physiol. 2017, 173, 2265–2277. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Li, S.; Zhang, M.; Yang, L.; Liu, Y.; Xu, J.; Zhang, S. Regulation of GDSL Lipase Gene Expression by the MPK3/MPK6 Cascade and Its Downstream WRKY Transcription Factors in Arabidopsis Immunity. Mol. Plant-Microbe Interact. MPMI 2019, 32, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Duan, P.G.; Rao, Y.C.; Zeng, D.L.; Yang, Y.L.; Xu, R.; Zhang, B.L.; Dong, G.J.; Qian, Q.; Li, Y.H. SMALL GRAIN 1, which encodes a mitogen-activated protein kinase kinase 4, influences grain size in rice. Plant J. 2014, 77, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Duan, P.; Yu, H.; Zhou, Z.; Zhang, B.; Wang, R.; Li, J.; Zhang, G.; Zhuang, S.; Lyu, J.; et al. Control of Grain Size and Weight by the OsMKKK10-OsMKK4-OsMAPK6 Signaling Pathway in Rice. Mol. Plant 2018, 11, 860–873. [Google Scholar] [CrossRef]
- Pei, L.; Gao, Y.; Feng, L.; Zhang, Z.; Liu, N.; Yang, B.; Zhao, N. Phenolic Acids and Flavonoids Play Important Roles in Flower Bud Differentiation in Mikania micrantha: Transcriptomics and Metabolomics. Int. J. Mol. Sci. 2023, 24, 16550. [Google Scholar] [CrossRef]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The Flavonoid Biosynthesis Network in Plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Tian, Y.; Wu, K.; Ye, Y.; Yu, J.; Zhang, J.; Liu, Q.; Hu, M.; Li, H.; Tong, Y.; et al. Modulating plant growth–metabolism coordination for sustainable agriculture. Nature 2018, 560, 595–600. [Google Scholar] [CrossRef]
- He, M.; Qin, C.X.; Wang, X.; Ding, N.Z. Plant Unsaturated Fatty Acids: Biosynthesis and Regulation. Front. Plant Sci. 2020, 11, 390. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, S. Changes of nucleic acid content in fruit and unfruitful trees during citrus flower bud differentiation. J. Southwest Univ. 2008, 6, 56–59. [Google Scholar]
- Yu, Z.; Jia, D.; Liu, T. Polyamine Oxidases Play Various Roles in Plant Development and Abiotic Stress Tolerance. Plants 2019, 8, 184. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Shumayla; Verma, P.C.; Singh, K.; Upadhyay, S.K. Molecular characterization of ascorbate peroxidase (APX) and APX-related (APX-R) genes in Triticum aestivum L. Genomics 2020, 112, 4208–4223. [Google Scholar] [CrossRef] [PubMed]
- Vaish, S.; Gupta, D.; Mehrotra, R.; Mehrotra, S.; Basantani, M.K. Glutathione S-transferase: A versatile protein family. 3 Biotech 2020, 10, 321. [Google Scholar] [CrossRef] [PubMed]
- Gho, Y.S.; Kim, S.-J.; Jung, K.H. Phenylalanine ammonia-lyase family is closely associated with response to phosphate deficiency in rice. Genes Genom. 2020, 42, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Yuyama, I.; Ishikawa, M.; Nozawa, M.; Yoshida, M.A.; Ikeo, K. Transcriptomic changes with increasing algal symbiont reveal the detailed process underlying establishment of coral-algal symbiosis. Sci. Rep. 2018, 8, 16802. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Kim, S.; Sagong, H.Y.; Son, H.F.; Jin, K.S.; Kim, I.K.; Kim, K.J. Structural basis for cytokinin production by LOG from Corynebacterium glutamicum. Sci. Rep. 2016, 6, 31390. [Google Scholar] [CrossRef]
- Zhang, S. Recent Advances of Polyphenol Oxidases in Plants. Molecules 2023, 28, 2158. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Names | Foldchange (Y-vs.-H) |
|---|---|---|
| TRINITY_DN8398_c0_g1 | nonexpressor of pathogenesis-related protein 1-like 5 proteins | 24.93 |
| TRINITY_DN650_c1_g1 | indole-3-acetic acid-induced protein ARG7-like | 7.54 |
| TRINITY_DN4000_c1_g1 | auxin-responsive protein IAA30-like | 5.01 |
| TRINITY_DN1329_c1_g1 | indole-3-acetic acid-amido synthetase GH3.6-like | 6.91 |
| TRINITY_DN72594_c0_g1 | BRI1 kinase inhibitor 1-like | 30.17 |
| TRINITY_DN3428_c0_g1 | protein TIFY 6B-like | 6.20 |
| TRINITY_DN7228_c0_g1 | transcription factor TGA2-like isoform X2 | 15.13 |
| TRINITY_DN7013_c1_g1 | protein TIFY 9 | 22.51 |
| TRINITY_DN10361_c0_g1 | Pathogenesis-related protein 1 | 56.05 |
| TRINITY_DN5341_c0_g1 | Transcription factor HBP-1b(c1) | 38.52 |
| TRINITY_DN7106_c2_g1 | auxin-induced protein 6B-like | 43.38 |
| TRINITY_DN34019_c0_g1 | Transcription factor MYC2 | 6.79 |
| TRINITY_DN5260_c1_g1 | auxin-induced protein 15A-like | 98.23 |
| TRINITY_DN17023_c2_g1 | hypothetical protein ZOSMA_82G00740 | 8.18 |
| TRINITY_DN9064_c0_g1 | regulatory protein NPR5 | 10.40 |
| TRINITY_DN3222_c2_g1 | hypothetical protein OsI_24760 | 21.02 |
| DAM | Mean_H | Mean_Y | Fold Change_H/Y | log2(FC_H/Y) | p Value |
|---|---|---|---|---|---|
| L-4-Hydroxyglutamate semialdehyde | 14,854,137.1 | 1737381.2 | 8.55 | 3.10 | 0.03 |
| N, N-Dimethylaniline | 5,953,932,280.7 | 118,333,143.3 | 50.32 | 5.65 | 0.01 |
| 2-Heptanone | 478,865,629.4 | 988.8 | 484,260.0 | 18.89 | 0.01 |
| 1-Pyrroline-5-carboxylic acid | 80,591,608.6 | 46,463,691.7 | 1.73 | 0.79 | 0.01 |
| 1-Benzyl-1,2,3,4-tetrahydroisoquinoline | 68,788,352.0 | 988.8 | 6,9564.0 | 16.09 | 0.00 |
| 2-Methylbenzoic acid | 949,974,505.8 | 57,384,295.4 | 16.56 | 4.05 | 0.01 |
| gamma-Glutamyl-beta-aminopropiononitrile | 5,588,130.7 | 988.8 | 5,651.1 | 12.46 | 0.00 |
| Pyrrolidonecarboxylic acid | 30,079,215.7 | 3,836,016.5 | 7.84 | 2.97 | 0.02 |
| Ketoleucine | 26,500,401.0 | 988.8 | 26,799.0 | 14.71 | 0.00 |
| (R)-5,6-Dihydrothymine | 279,791,870.5 | 114,946,419.6 | 2.43 | 1.28 | 0.01 |
| Fenfluramine | 10,258,146.5 | 5,721,365.4 | 1.79 | 0.84 | 0.01 |
| Sulfamethazine | 22,024,321.9 | 12,887,857.9 | 1.71 | 0.77 | 0.01 |
| Porphobilinogen | 156,535,147.6 | 115,434,618.3 | 1.36 | 0.44 | 0.01 |
| Gyromitrin | 284,576,546.4 | 988.8 | 287,790.0 | 18.14 | 0.00 |
| 3-Methyl-2-oxovaleric acid | 180,493,405.8 | 988.8 | 182,530.0 | 17.48 | 0.00 |
| Phenylethylamine | 81,993,597.7 | 988.8 | 82,918.0 | 16.34 | 0.00 |
| Capsidiol | 120,373,230.9 | 105,812,770.2 | 1.14 | 0.19 | 0.02 |
| Biomarker | Pathway | Pathway ID | Up Number | Down Number | Total Number | p-Value | KEGG ID | VIP | log2(FC-H/Y) | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|
| L-4-Hydroxyglutamate semialdehyde | Arginine and proline metabolism | ko00330 | 3 | 3 | 55 | 0.874 | C05938 | 1.138 | 3.096 | 0.031 |
| 1-Pyrroline-5-carboxylic acid | Arginine and proline metabolism | ko00330 | 3 | 5 | 43 | 0.507 | C04322 | 1.734 | 0.795 | 0.010 |
| γ-Glutamyl-β-aminopropiononitrile | Cyanoamino acid metabolism | ko00460 | 2 | 5 | 43 | 0.507 | C06114 | 1.088 | 12.464 | 0.003 |
| Pyrrolidonecarboxylic acid | Glutathione metabolism | ko00480 | 14 | 5 | 90 | 0.094 | C01879 | 1.115 | 2.971 | 0.025 |
| Phenylethylamine | Phenylalanine metabolism | ko00360 | 9 | 3 | 50 | 0.076 | C05332 | 1.392 | 16.339 | 0.003 |
| 1-Benzyl-1,2,3,4-tetrahydroisoquinoline | Isoquinoline alkaloid biosynthesis | ko00950 | 3 | 1 | 20 | 0.375 | C05201 | 1.478 | 16.086 | 0.003 |
| (R)-5,6-Dihydrothymine | Pyrimidine metabolism | ko00240 | 4 | 9 | 71 | 0.300 | C21028 | 1.679 | 1.283 | 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Xu, P.; Qian, C.; Zhao, X.; Xu, H.; Li, K. The Combined Analysis of the Transcriptome and Metabolome Revealed the Possible Mechanism of Flower Bud Formation in Amorphophallus bulbifer. Agronomy 2024, 14, 519. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030519
Li W, Xu P, Qian C, Zhao X, Xu H, Li K. The Combined Analysis of the Transcriptome and Metabolome Revealed the Possible Mechanism of Flower Bud Formation in Amorphophallus bulbifer. Agronomy. 2024; 14(3):519. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030519
Chicago/Turabian StyleLi, Wenchao, Peng Xu, Cheng Qian, Xing Zhao, Huini Xu, and Kunzhi Li. 2024. "The Combined Analysis of the Transcriptome and Metabolome Revealed the Possible Mechanism of Flower Bud Formation in Amorphophallus bulbifer" Agronomy 14, no. 3: 519. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030519