

Diversity as a Plant Breeding Objective

Independent Consultant, 63100 Ascoli Piceno, Italy
*
Author to whom correspondence should be addressed.
Agronomy 2024, 14(3), 550; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030550
Submission received: 30 December 2023
/
Revised: 19 February 2024
/
Accepted: 6 March 2024
/
Published: 8 March 2024
(This article belongs to the Special Issue Novel Studies in Crop Breeding for Promoting Agro-Biodiversity)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:This review paper addresses the importance of increasing agrobiodiversity to cope with climate change and, at the same time, providing a sufficient amount of healthy food. This is in agreement with the messages from ecology and medicine indicating the advantages of biodiversity in general and agrobiodiversity in particular for the planet and for our health. Plant breeding is considered to be one of the causes of the decline in agrobiodiversity, and therefore, this paper illustrates alternatives to the commonly used approach based on centralized selection. The first alternative is decentralized participatory breeding, which adapts crops to both different agronomic environments and client preferences, representing an “option by context” model of research. The second alternative is evolutionary breeding, which is a more dynamic strategy than participatory plant breeding because it merges the advantages of decentralization with the ability of dynamic mixtures and evolutionary populations to cope with biotic and abiotic stresses and evolve, thus adapting to climate change and to the associated changes in the spectrum of pests. A crop capable of evolving as the environment around it evolves appears to be the most ideal way of responding to climate change and increasing agricultural biodiversity.
1. Introduction
Modern plant breeding needs to address the most important global challenges, such as climate change, poverty, malnutrition (which includes undernutrition and obesity), water scarcity, and the loss of biodiversity. These are the challenges most frequently debated in international reports [1]. In the plant breeding literature, climate change seems to be the issue most frequently addressed using terminologies such as “climate-resilient crops” [2], “climate-ready crops” [3], and “climate-smart crops” [4]. In our opinion, these are oversimplifications, as climate resilience is a complex breeding objective. The complexity is due to the uncertain quantification of temperature increase [5] or rainfall decrease [6], the location specificity of climate change [7], and the associated changes in the spectrum of diseases, insects, and weeds [8]. An additional element of complexity is that climate change is likely to be a moving breeding objective, as indicated by the progressive increase in the frequency of temperature anomalies in the last 50 years [9].
Climate change has already affected plant breeding by placing resilience as a top priority in breeding [10,11], while at the same time recognizing the unpredictability of the impact of climate change. This has prompted the use of predictive models to inform about breeding priorities [12], including the choice of the right crops to combine climate adaptation and healthy human nutrition [13].
Plant breeding has traditionally responded and continues to respond to this complexity by breeding almost exclusively uniform varieties [14,15], which, depending on the mating system, the objectives, and the crop, range from hybrids, clones, and pure lines. This trend was already criticized by Frankel in the 1950s [16] as follows (quote from pages 97–98): “From the early days of plant breeding, uniformity has been sought after with great determination. For this there are many reasons—technical, commercial, historical, psychological, aesthetic”. “It seems to me that the “concept of purity has not only been carried to unnecessary length but that it may be altogether inimical to the attainment of highest production” since it is “concerned with characters which are readily seen but often of little significance”. Note that Frankel never used terms such as “scientific” or “biological” as reasons to breed for uniformity. Also, Frankel could not have known that he was anticipating what ecology and medicine would recommend years after his publication. Despite these early warnings, which have been echoed more recently [17,18,19,20], and despite its potential benefits, agrobiodiversity was greatly reduced in terms of the number of crops and genetically homogenous varieties within crops [21], while ecology and medicine both emphasized the importance of diversity for resilience and human health.
2. Messages from Ecology
Ecology studies the capacity of the environment to support the requirements of contemporary human societies, namely the interactions between organisms and their environments [22]. Ecological field experiments have shown that greater diversity, expressed as species richness, is associated with greater productivity and greater stability [23,24,25,26]. The question of whether the ecosystems that are biologically more diverse are also more stable, very relevant today in the face of climate change and the need for sustainability, has fascinated ecologists for generations [25] and has been at the center of debate [27].
Before the 1970s, ecologists believed that more diverse communities enhanced ecosystem stability. These early intuitive ideas were challenged by the work of Robert May [28], who, using linear stability analysis, found that diversity tends to destabilize community dynamics. However, the most recent literature indicates a positive relationship between crop diversity, ecosystem stability, and productivity [29,30,31].
A number of experiments on organisms ranging from bacteria, fungi, plants, and animals have shown a positive relationship between species diversity and biomass production [32,33]. This effect, defined as the richness effect, occurs when a mixture (of species, genotypes, functional groups, etc.) performs better than the average performance of its components in monoculture and is also known as non-transgressive overyielding [34]. The phenomena explaining this effect are a combination of complementary effects (when species occupy different niches so that they use resources in a complementary way) and sampling effects; the sampling effect occurs because more diverse communities are more likely to host highly productive species. This also requires that the most productive species become predominant, and this is why the sampling effect is also called selection effects [21]. The main consequence of sampling effects is that a diverse community of plants, such as a mixture, is expected to yield more than the average of its components.
The controversy stems from studies showing that the most diverse communities do not always produce more biomass than their most productive component, suggesting that the results of such experiments are better explained by the sampling effect. However, the analysis of experiments conducted over a long time suggests that both effects occur and that the contribution of the most productive species is equal to or exceeded by species complementarity [25].
If we replace “species” with “genotypes”, the implications of these studies for plant breeding become obvious because they suggest that diversity could become an objective of plant breeding programs that have yield increases as one of their main objectives. What is of interest in relation to plant breeding strategies is that diversity is not the driver of this relationship; rather, ecosystem stability depends on the ability of communities to contain species, or functional groups, which are capable of differential responses [27].
3. Messages from Medicine
Several studies in medicine journals indicate an association between the decline in biodiversity and a number of human health problems. The reference to plant breeding, although not always explicit in these studies, is implicit when the effect of the diet on the composition and diversity of the microbiota—the complex of bacteria, viruses, fungi, yeasts, and protozoa in our intestine—on our physical and mental health are discussed. The prevalent dietary uniformity related to the decline in agrobiodiversity has been associated with the increase in inflammatory diseases in humans, ranging from inflammatory bowel disease to ulcerative colitis, cardiovascular disorders, various liver diseases, and many types of cancer [35]. The increased frequency of inflammatory diseases has been associated with a decreased efficiency of our immune defenses [35,36].
The relationship between agrobiodiversity and dietary diversity is complex, as both are affected by other factors [37]. A review of five databases revealed that agrobiodiversity has a small but consistent association with more diverse household and individual diets, although the magnitude of this association varies with the extent of farm diversification [38]. A study in Nepal revealed significant associations between child dietary diversity and agrobiodiversity [39].
Diet strongly influences the microbiota: a diet modification changes its composition in just 24 h. It takes 48 h after changing the diet back again for the microbiota to return to its initial conditions [40]. A recent study has shown a dramatic reduction in the diversity of the microbiota from more than 700 different species of microorganisms in the microbiota of an average adult of a Tanzanian tribe to less than 300 in the microbiota of an average adult in California [41].
More recently, the microbiota has been shown to regulate both brain and behavior, especially those associated with social functions [42], to be associated with the development of tumors [43] and with our immune system [44] to the point that microbiota transplant has become a focus in medicine.
Given the important roles of the microbiota on one hand, and the fact that it is so strongly and rapidly influenced by diet on the other, it is understandable that there have been many studies on the effect of various diets (western, omnivorous, Mediterranean, vegetarian, vegan, etc.) on its composition and diversity [40]. Recent results have demonstrated that diet diversity is of paramount importance for a healthy microbiota [45,46]. But how can we have a diversified diet if the agriculture that produces our food is predominantly based on uniformity, with three crops providing over 60% of the calories of plant origin we consume [47]?
The reduction in diversity, such as the loss of ancient wheat varieties, has health implications as those varieties have unique flavors and cultural significance to local cuisine [48], have a higher phenolic content [49,50], reduce glycemia [51], and have beneficial effects on various parameters linked to cardio-metabolic diseases such as lipid and glycemic profiles, as well as the inflammatory and oxidative status [49].
A recent review [52] compared the nutritive value of landraces with that of modern varieties in a number of crops ranging from tomato, melon, lettuce, beans, eggplants, garlic, onion, soybean, hazelnut, rice, lentil, chickpea, carrot, apple, grape, and maize. The comparison showed that landraces tend to have a higher content of vitamins (the precursor of vitamin A and vitamins C and E), micronutrients (such as Cu, Fe, Mn, and Zn), and phenolic compounds. Furthermore, a number of papers demonstrated the beneficial dietary effects of a larger agrobiodiversity on child nutrition [39,53,54,55].
More recently, the most extreme cases of biodiversity loss, such as deforestation and species extinction, have been related to an increased risk of pandemics such as COVID-19 [56].
4. Breeding Strategies to Increase Diversity
We have seen in Section 1 that plant breeding has contributed enormously to yield increases in major crops but, at the same time, has also reduced agrobiodiversity; in the following section, and after mentioning the main features of a plant breeding program, we will discuss how plant breeding can combine yield improvement with the maintenance or even the increase in agrobiodiversity.
4.1. Plant Breeding
- Generating genetic variability; this includes the selection of parents, making crosses, choice of type and number of crosses, induced mutation, introduction of accessions from germplasm banks, or material from other breeding programs or from farmers;
- Selection of the best genetic material within the genetic variability created or acquired in stage one. This stage lasts a number of cropping seasons (5 to 10) depending on the crop and on the methodologies used;
- Testing of breeding lines; this includes comparisons between existing cultivars and the breeding lines emerging from stage 2 using the appropriate experimental designs and statistical analysis to conduct such comparisons. This stage lasts at least three cropping seasons, and it is followed by the variety registration, the seed production, and, eventually, by farmers’ adoption.
As shown in Figure 1, a characteristic of a breeding program is that the best material that comes out of a breeding cycle returns to the next cycle as parental material.
The most common way of generating genetic variability in stage 1 is to make crosses among parental lines selected for specific traits. The number of crosses generated at the beginning of each cycle can vary from a few hundred in national breeding programs to several thousand in international breeding programs. Using accessions from germplasm banks is another way of bringing genetic variability into a breeding program. The use of material held in gene banks is not carried out often enough [58].
During stages 2 and 3, the genetic variability generated in stage 1 is gradually reduced via selection, and promising breeding lines are identified. While the number of breeding lines decreases, the quantity of available seed per line increases, and therefore, the material can be tested in an increasing number of locations [59]. The amount of material a breeding program handles on a yearly basis is large as a new cycle starts every year (or twice a year).
Molecular tools such as marker-assisted selection and genomic selection are designed to improve the efficiency in identifying useful genetic combinations in stage 2, but they do not necessarily translate into a higher breeding efficiency, i.e., into a higher efficiency of the entire process.
In its most common implementation, the key stage of a plant breeding program, namely the selection stage, takes place in one or a few research stations (centralized selection) under optimum or near-optimum agronomic conditions, including the use of fertilizers and pesticides. In some cases, breeding programs conducted on research stations are managed to simulate specific stress conditions [60]. The breeding philosophy of a breeding program based on centralized selection concerns two fundamental concepts in plant breeding, namely adaptation and stability. Adaptation is defined as the ability of a variety to perform well in a given location, and stability is defined as yield consistency across years [61,62,63]. The philosophy, which in the 1960s became predominant despite a number of critical voices, was that of “wide adaptation” [64], and this philosophy found its international expression in what is known as the “Green Revolution”. The “Green Revolution” model of agricultural development was based on the selection of varieties capable of excelling everywhere and at all times, having lost their sensitivity to photoperiod and the need for vernalization. Moreover, these varieties were short, with very strong culms, and were therefore able to withstand heavy nitrogen fertilization without lodging. The expression Green Revolution was coined in March 1968 by William S. Gaud, the then director of the US Agency for International Development (USAID), to indicate the effect of a development strategy based on (a) new varieties, (b) irrigation, (c) fertilizer, (d) pesticide, and (e) mechanization. Under this strategy, new varieties characterized by “extensive adaptation” were obtained. Not only was this exactly the opposite of what farmers had been carrying out for millennia but it also ran counter to the genetic improvement philosophy of the early part of the 20th century. According to that philosophy, selection had to be performed at the place where the varieties were to be grown, producing varieties with specific adaptations to local conditions [64].
The term “wide adaptation” was misleading because it referred to “geographical” rather than “environmental” adaptation [65]. In fact, the agricultural environments in which these “widely adapted” varieties were successful were very similar (high rainfall, or availability of irrigation water, and good soil fertility), although geographically very distant, or were made similar to each other with the addition of irrigation water, fertilizers, and pesticides.
One consequence of breeding for wide adaptation has been the replacements of landraces in many countries by the widely adapted, genetically uniform, semi-dwarf varieties of relatively few crops, such as rice and wheat [18], with a decrease in diversity both between and within crops. The genetic erosion has been estimated at 72.4% and 72.8%, respectively. For example, the number of rice cultivars declined in Indian farms from about 400,000 before colonialism to 30,000 in the mid-19th century, with unknown thousands more lost after the Green Revolution. Greece lost 95% of its wheat landraces after being encouraged to replace them with modern cultivars [66].
In the last 50 years, recombinant DNA technology has introduced a new dimension into plant breeding, making trait management more targeted and precise, and breaking the barriers between species [67]. The use of GMOs and of gene editing have raised a number of regulatory and ethical issues, and, in the case of GMOs and, more recently, of gene editing, controversy about their actual success [68].
Alternative plant breeding methodologies, such as participatory plant breeding and evolutionary plant breeding, the latter based on the use of non-uniform varieties such as mixtures and evolutionary populations, have shown the possibility of bringing back diversity in agriculture by combining productivity with resilience. The two methodologies can be combined in a participatory evolutionary breeding program.
4.2. Participatory Plant Breeding
Participatory plant breeding (PPB) is, like all breeding programs, characterized by the three stages described above, with three major differences from non-participatory breeding programs, namely:
- Objectives and type of final product (known as product profile) are discussed and decided in collaboration with the clients—generally, but not exclusively, the farmers;
- The program is conducted in farmers’ fields and therefore is decentralized, i.e., conducted in the target environment(s) rather than in the research station(s);
- The clients participate in all the most important decisions, including the choice of parents to be used for crossing and in the selection stage.
One of the main drivers of PPB has been the need to address the farmers bypassed by the Green Revolution and, in general, by the centralized breeding programs inspired by it, and the most marginal agronomic environments for either climatic or socioeconomic reasons or for a combination of the two [69,70,71]. Another important driver has been the exploitation of positive genotype x environment interactions by selecting for specific adaptation rather than for wide adaptation [72]. Using selection theory and the breeder’s equation, it is possible to demonstrate that PPB is more efficient than centralized-non participatory plant breeding when efficiency is measured by adoption, increase in agrobiodiversity, and a higher benefit/cost ratio [73].
When clients are only involved in stage 3, the term participatory variety selection (PVS) has been used. With PVS, clients have many more limited choices to make as compared with PPB, but PVS has been considered an easy-to-be-organized type of participation, which may eventually develop into a PPB program provided it is fully decentralized [72].
There are several examples of PPB programs, which have been reviewed in terms of crops, countries, and institutions globally [74], as well as for countries in the north [71].
The first review [74], which covered up to May 2018, was based on 254 publications on PPB and PVS, dealing with 47 crops, including self-pollinated, cross-pollinated, and vegetatively propagated crops. The crops most frequently addressed in PPB or PVS programs were rice, maize, bean, sorghum, barley, potato, bread wheat, and pearl millet. The countries that hosted PVS and PPB programs and were most frequently cited in the 254 publications were India, Ethiopia, Mali, Nepal, Nicaragua, the USA, Burkina Faso, and Syria.
The two reviews also indicated that the major successes achieved by PPB programs ranged from yield increases, as in the case of barley in Syria, cassava in Ghana, and tomato in Italy, to disease and lodging resistance combined with quality traits as in the case of rice in Nepal, greater yield stability as in the case of wheat in France, and adaptation to organic systems as in the case of several crops, particularly in the USA.
Of particular interest is that as many as 60 universities contributed publications to either PPB or PVS, with 75% of the publications contributed by 11 universities in developed countries, mostly from the USA, UK, and Italy. In those countries, PPB has been mainly implemented in organic agriculture [75]. This is what also emerges from the most recent review [71], which identified 47 projects across the United States, Canada, and Europe, including 22 crops. It is not surprising that a theme common to all these projects is adaptation to organic farming because, from a breeding viewpoint, organic agriculture represents a heterogenous population of target environments, which is very different from the mostly homogenous population of target environments typical of industrial agriculture [76].
We argue that because of this difference, organic and industrial agriculture cannot be served by the same breeding strategies and methodologies and by the same seed system, which is based on the deployment of a small number of uniform varieties over large areas [76].
PPB represents an “options by context” model of research [77], which focuses on local adaptation as a way to find a “basket of options” from which farmers can choose. PPB carries out exactly this in the realm of new varieties.
The major weaknesses, which emerged over the years and prevented a generalized institutionalization of PPB, have been the seed regulatory systems and the unsustainability of the collaborative mode that was often associated with the personal interest of individual managers rather than an institutional commitment.
These weaknesses prompted an interest in a more dynamic, decentralized, farmer-run (without necessarily excluding institutions) breeding strategy, such as evolutionary plant breeding.
4.3. Evolutionary Plant Breeding
Evolutionary plant breeding (EPB) was first proposed in 1956 [78], although the original idea emerged nearly 30 years earlier [79] using the term “composite hybrid mixtures”. This original paper exemplified the method first by describing how 28 barley varieties were chosen among a world collection of about 5000 barley varieties, and then how all 378 possible crosses between 28 barley varieties (excluding reciprocals and selfing) were made, and eventually, how “equivalent quantities” of the F2 seed were mixed. The authors thought that handling such a large number of crosses individually was burdensome and therefore suggested growing the mixtures in several locations and letting natural selection eliminate the “weaker combinations”. After a few years, the mixture would be made largely of homozygotes, and a single plant or head selection could be made to isolate desirable varieties.
The science of EPB has evolved ever since and produced a large body of research initially carried out mostly on self-pollinated crops such as barley, rice, and beans. There is a problem with the terminology in the literature, as mixtures, bulk populations, bulks, composites, composite cross (abbreviated as CC), composite cross populations (abbreviated as CCP), and evolutionary populations (EP, the term we will use thereafter) have been used. All the terms except “mixtures” correspond to the original description of Harlan and Martini [79]. The term mixture, used alone, indicates a physical mixture of seeds of different varieties or accessions. Mixtures are “static” when they are made up by mixing a given number of seeds of each component at the beginning of each cropping season. They are static because, although such physical seed mixtures are genetically more complex than monocultures and can therefore be subjected to natural selection, they do not capture the effects of natural selection occurring in the field. When part of the grain produced from mixtures is used as seed for the following season, thus capturing the effects of natural selection, the mixtures are “dynamic”. A dynamic mixture, over a few generations, becomes a “population” [80] as defined above because of out-crossing, which, even in low percentages, also occurs in self-pollinated crops, leading to segregation and new gene combinations.
The research conducted on mixtures and evolutionary populations (EP) has been summarized recently [81] with the following main findings:
- They evolve, becoming more and more productive. The first indication of a progressive evolution towards a higher yield came from work on barley and lima bean [84], followed by experiments with barley [78,85,86,87]. A recent study on barley shows that EPs are as productive, in terms of grain yield, as commercial varieties under low input conditions [88]. This has been confirmed in a number of studies [87,89,90,91,92,93,94,95,96], indicating a higher resilience of populations and mixtures as expected from ecological studies;
- EPs, and to a lesser extent mixtures, have a more stable yield over time than uniform varieties, but not over space, i.e., they become specifically adapted to a given location. The original paper on the superior stability of populations over mixtures and of mixtures over pure lines was published in 1961 [91]; the paper reported the results of an experiment with lima beans in which stability was measured as the consistency of ranks. The results showed that while the pure lines were successful in many environments (high frequency of low ranks) and unsuccessful in many others (high frequency of high ranks), the mixtures and the populations were intermediate in any one environment. The higher stability of mixtures and populations has been confirmed in oats [97], wheat [90,93,98,99,100], and barley [88,92,101];
- EPs and mixtures evolve, becoming more and more resistant to diseases. This effect of populations and mixtures has been by far the most extensively studied by both breeders and pathologists [92,102,103,104,105,106,107,108,109,110]. The most important mechanism to explain the reduction in disease severity in mixtures and EPs is the dilution of inoculum that occurs due to the distance between plants of the same genotype [104]. However, there is also a large variation in the efficacy of mixtures in reducing disease incidence;
- The ability of EPs and mixtures to control weeds, a particularly important issue in organic farming where options to control weeds are limited, has not been studied as extensively as disease resistance. One hypothesis [111] claimed that communities with more diversity are more resistant to invasive species than communities with less diversity. A study conducted with a perennial species showed that the weed biomass decreased by 32% from a community consisting of a single genotype (thus corresponding to a uniform variety) to a community consisting of 12 different genotypes [112]. This result not only confirms the hypothesis above but also extends it from diversity between species to diversity within species, which is particularly relevant for mixtures and EPs.
A number of studies have suggested that old cereal varieties and landraces control weeds better than modern varieties [113,114,115], and this seems to be associated with a higher biomass accumulation, high tillering ability, and tallness of the old varieties [116].
The ability of mixtures and EPs to control weeds better than modern varieties is likely associated with the fact that the former is much taller than the latter. The increase in plant height during the evolution of EPs was reported in barley [117] and in wheat [118], where the increase in plant height was accompanied by a decrease in the number of kernels per spike and kernel size. An increase in plant height was also found to be associated with late flowering time [119]. The relation between height and weed suppression was studied in spring barley, showing that mixtures of varieties of different heights are more effective in suppressing tall annual weeds than their best component [92].
One of the effects of climate change is a higher frequency of extreme events, including flooding. It is therefore of interest that a study conducted in Germany after the natural flooding of July 2013 showed that, on average, plant species were less affected by flooding when grown in high-diversity plots. Furthermore, soil porosity was higher in high-density plots, which were less affected by flooding, resulting in better plant performance [120].
Until recently, and despite all the proven advantages described above, EPs and mixtures have not been extensively used in agriculture, with the exception of the well-known example of malting barley mixtures [121]. In that case, the malting barley mixtures quickly spread on over 10,000 hectares in Poland, particularly in East Germany, where they were introduced in 1984. They were successful because the mixtures had been formulated specifically for powdery mildew resistance and malting quality. At the end of the 1980s, the mixtures covered almost all the spring malting barley area (over 300,000 hectares) with a reduction in the disease incidence from 50% to 10%, while the use of the fungicide was reduced to just one treatment on over 100,000 hectares. There was no decrease in production, while the malt quality was considered satisfactory. With the unification of the two Germanies, the project was abandoned due to the preference of Western European malters for malt obtained from individual varieties, even if treated with fungicides.
The major obstacle to translating into practice the research findings about the benefits of EPs and mixtures was likely seed legislation, which, in several countries, is based on the principles of variety registration and seed certification. Seeds can be marketed only if they belong to a variety that has been registered and if the seeds have been certified. To be registered, a variety must satisfy distinctness, uniformity, and stability requirements [122] regardless of whether it was obtained by conventional, participatory, or evolutionary breeding.
However, the European Union, recognizing the advantages of EPs and mixtures, has considerably invested in research on diversity in agriculture over the last 10 years. This research was conducted through projects such as “Strategies for Organic and Low-Input Integrated Breeding and Management” (SOLIBAM) from 2010 to 2014, “Embedding Crop Diversity and Networking for Local High Quality Food Systems” (DIVERSIFOOD) from 2015 to 2019, “Boosting Organic Seed and Plant Breeding across Europe” (LIVESEED), and eventually with the new EU “Farm to Fork” initiative. In 2014, likely as a consequence of the growing interest to bringing back diversity into agriculture, the European Commission (EC) started a temporary experiment “providing for certain derogations for the marketing of populations of the plant species wheat, barley, oats and maize” [123]. As a result of that experiment, 46 populations (5 maize, 3 barley, and 38 wheat populations) were authorized, and the seed of 31 populations (5 maize, 2 barley, and 24 wheat) was legally commercialized [124]. In 2018, the EC adopted a new regulation [125], which lays down new rules on the control of organic production and labeling of organic products. It makes available for use in organic production “plant reproductive material that does not belong to a variety, but rather belongs to a plant grouping within a single botanical taxon with a high level of genetic and phenotypic diversity between individual reproductive units” [125]. In this regulation, the material is defined as a heterogeneous material.
Although the regulation applies only to organic agriculture, it obviously paved the way for using EPs in organic breeding programs with the perspective of legally commercializing the seed of the final products, irrespective of their uniformity and stability. At the moment of writing, a number of populations have been notified to the appropriate authorities, but it is still too early to have precise figures.
4.4. Combining Participation with Evolutionary Plant Breeding
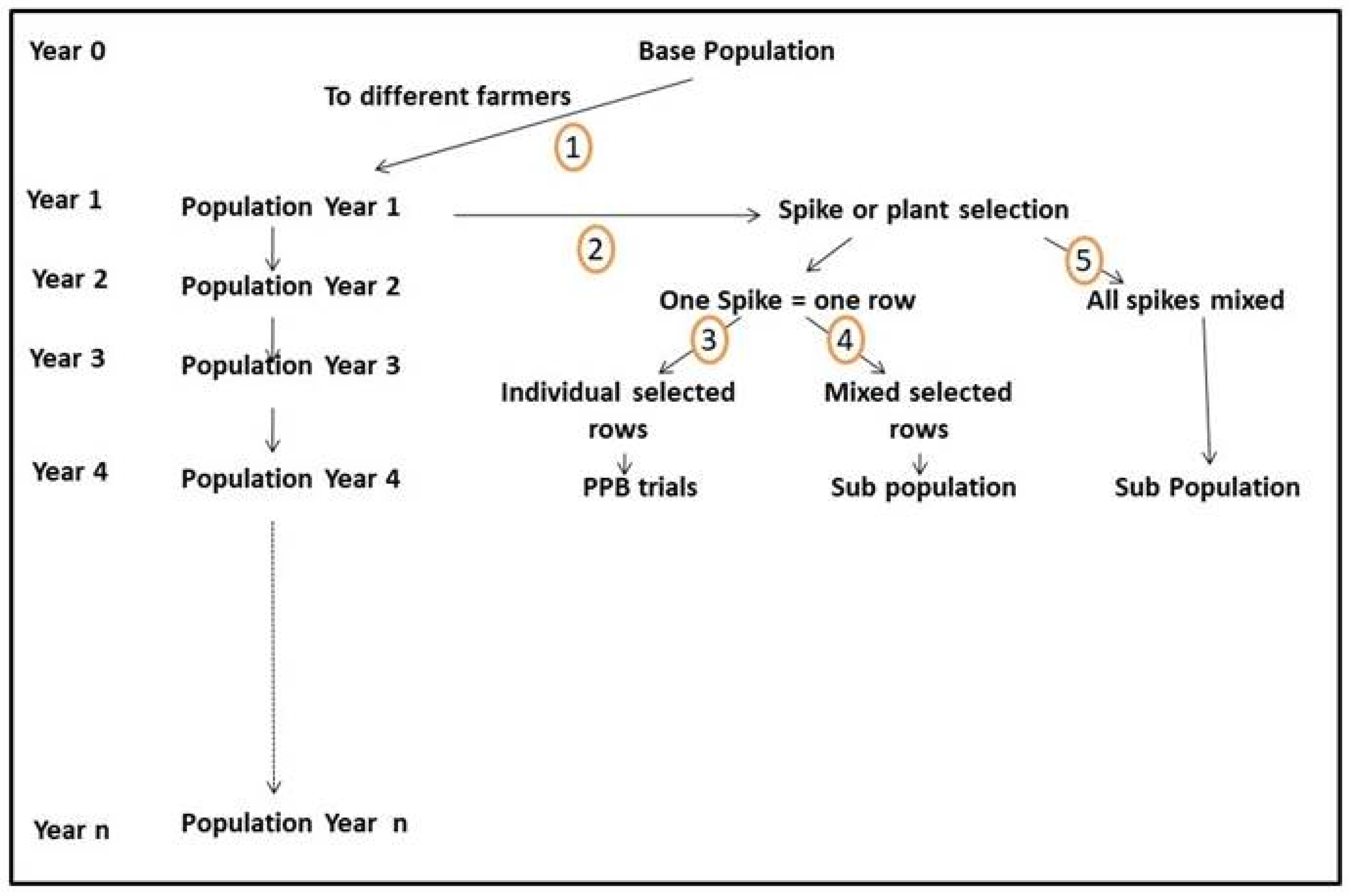
Participation can become a component of a breeding program using EPs or dynamic mixtures as a source of genetic diversity (step 1 in Figure 1).
The EPs can be the base of a breeding program, such as the one shown in Figure 2, in which a given EP is the dynamic source of diversity. One of the main advantages of such a breeding program is that it could be implemented in countries where research institutions do not have the capability/expertise to make crosses and to handle segregating populations. A second advantage is that the EP from which the future varieties and/or populations will be derived could be planted in any type of environment to generate varieties specifically adapted to that environment.
At intervals that depend on the flow of the breeding material from one generation to the next, the population can be subjected to selection.
If the population is planted in one or more farmers’ fields representing a sample of the target population of environments, and the selection is carried out by the farmers, the program becomes a decentralized participatory breeding program based on an evolving population.
Depending on the crop, the object of selection can be the plant, as in the case of spaced-planted crops such as maize, sorghum, and most vegetables, or a spike, a panicle, a pod, like in cereals such as wheat, barley, or rice, or several legumes. Even in spaced planted crops, plant selection can be followed by fruit selection, as in tomato or eggplant, or cob selection, as in maize.
In the examples below illustrating the different ways in which selection can be carried out within the EP, given that, at least in Europe, the final product meant for organic farmers does not necessarily need to be uniform, we will use for simplicity the term spike for the unit of selection. This can be generalized to the various crops, as indicated above.
The spikes selected in any one year can be used in one of the following ways:
- The selected spikes are threshed individually, and the seed of each spike is planted separately in a row. This can be carried out by breeders on stations or, preferably, by farmers in their fields. If, because of technical problems, the head rows are planted on stations, they should only be multiplied, and selection should be delayed until the following year;
- If the head rows are planted in farmers’ fields, they should be planted under the same conditions (for example, reduced irrigation, shallow soil, or heavy weed infestation) in which they were selected in order to continue the selection;
- The seed collected on the selected rows can be handled in three different ways (Figure 2, paths 3, 4, and 5).
4.4.1. Spike Selection to Feed a PPB Program (Figure 2, Paths 2–3)
The seed harvested on the selected head rows can be planted in small plots and further selected, as shown in path 3. The seed produced on the selected small plots should be in sufficient amount to establish a Stage 1 PPB trial as described in [126]: each selected row will be an individual entry in the Stage 1 trial. Paths 2 and 3 can be repeated every year, leading to a situation where, in the village or in the research station, there will be short rows and small plots, and in the village, PPB trials at the various stages.
This is how evolutionary populations can feed a participatory breeding program: the participation will first occur with farmers’ selection of spikes from within the evolutionary populations and second with the evaluation by farmers of the plots derived from those spikes in the various stages of the PPB program.
If this is considered difficult to manage, a new cycle of spike selection can be initiated when the material derived from the previous cycle has reached the final stage.
This is not conceptually different from a conventional breeding program of cereal crops as the selection can be conducted in such a way as to also generate pure line varieties. The main difference is that since the original parent is a single spike selected from within an evolutionary population, the pedigree of the final variety indicating the original cross will not be available, and the material can only be genetically identified by molecular tools.
4.4.2. Spike Selection to Create Sub-Populations (Figure 2 Paths 2–4 or 2–5)
Rather than being planted again for another cycle of selection, the seed of the selected rows can be mixed to create a sub-population. Alternatively, the individual spikes can be mixed after harvesting and threshed together, and the resulting seed can be planted as a sub-population.
The two sub-populations resulting from paths 2–4 and 2–5 represent a way to accelerate the process of adaptation—the first is expected to be more efficient because the selection is based on families rather than on individuals. The (improved) sub-population could eventually become the farmers’ crop. However, it is advisable to maintain a sufficiently large area of the base population, which has a much wider genetic base and therefore a higher evolutionary potential.
A critical aspect of the breeding program illustrated in Figure 2, and currently implemented, for example, in the wheat and barley breeding programs conducted by the National Agricultural Research Center (NARC) in Jordan, is the continuous evolution of the EP in the target environments. Because of the combined effect of recombination and natural selection, new genotypes continue to appear that can be exploited via selection.
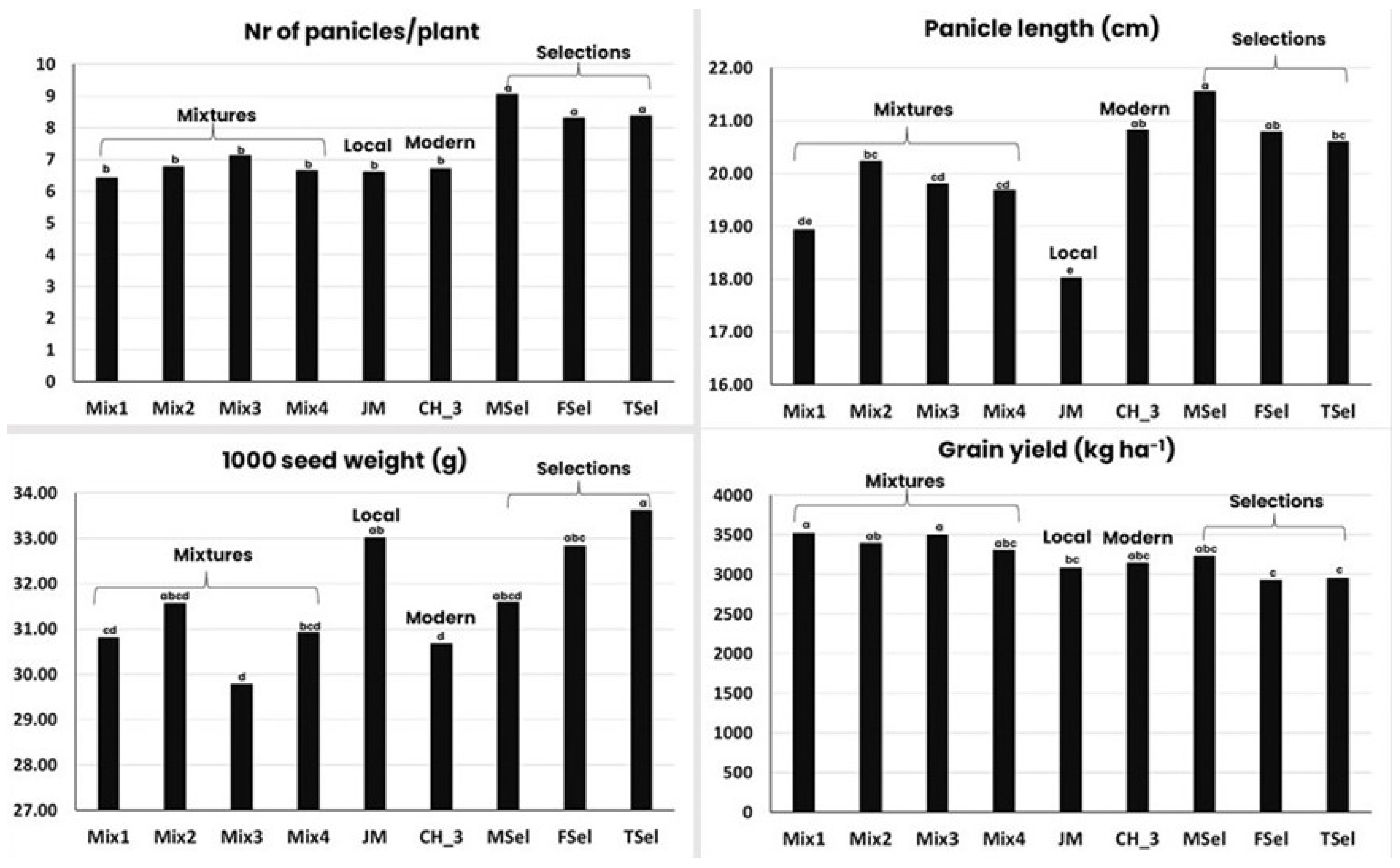
An example of how quickly artificial selection can change a number of traits in an evolving mixture comes from an experiment on rice in Nepal. The experiment comprised four types of dynamic mixtures: the first was composed of only local varieties from the area, the second was composed of local varieties from all over the country, the third was composed of improved varieties, and finally, the fourth, much more complex, composed of local varieties, improved varieties, and breeding lines. In this last mixture, the most diverse of the four, the farmers (men and women separately) and the technicians selected 100 plants they considered to be the best; the seeds of the 100 plants were mixed, and three newly selected mixtures were formed. The experiment also included a local variety and an improved variety as controls. The results [127] obtained with one cycle of visual selection on eight important traits are shown graphically in Figure 3 and Figure 4.
In a single selection cycle, both farmers and technicians were able, albeit with differences between them, to change flowering and maturity time, plant height, number of tillers (Figure 3), number of panicles (as an effect of increased tillering), panicle length, and seed size (Figure 4). Only the selection made by male farmers resulted in a population as productive as the modern variety and the most productive mixture, and in any case, none of the three selections was statistically higher yielding than the mixture within which the selection was made. This is probably due to the fact that, from a genetic point of view, yield is a very complex character, and more cycles of selection are needed to modify it significantly.
Overall, this experiment demonstrated how quickly and to what extent a number of agronomic and phenological traits can be changed by superimposing artificial selection to natural selection. This is not surprising because, even if the selection was only on a phenotypic basis, it was practiced on a broadly variable population. It also confirmed the “option-by-context” connotation of combining evolution with participation because of its capacity to elucidate gender-based differences in preference.
5. Discussion
One advantage of PPB and EPB is that, depending on the client profile, namely the type of client the breeding program aims to serve, it can produce different types of products. Even if the overall objective is to cultivate more diversity with the associated benefits described earlier, it is possible to reconcile diversity in the field and uniformity in the market for those crops where consumers’ preference is for uniform products. Some cases are described in [71]. For example, for crops and markets requiring uniformity, the breeder can follow the path 1-2-3 in Figure 2 and then use the PPB trials to produce uniform varieties. In a different context, such as organic farming in Europe, the breeder can follow the path 1-2-4 or 1-2-5, depending on the level of heterogeneity he/she wants to maintain in the final product.
It has been argued that PPB can be ineffective as it relies heavily on farmer selection, and therefore, farmer’s feedback is obtained only for those traits visible at the time and place of farmers’ selection and does not incorporate the requirements of millers, urban consumers, and other stakeholders [128]. This criticism does not consider that in PPB, there are a number of stakeholders that can actually be involved in selection. On the other hand, conventional plant breeding does not have to necessarily reduce agrobiodiversity, and this seems to be more the consequence of policies to the point that, although it may be counterintuitive that biodiversity laws will block breeding for diversity, current regulations seem to do exactly that [18]. Participatory plant breeding, besides being recognized as better suited to the needs of smallholder farmers, can be combined with genomics tools and crowdsourced citizen science [129].
A number of molecular tools are available to either provide alternative ways of modifying traits such as transgenics, mutation, and gene editing [2,130] or increase selection efficiency such as gene mapping or genomic selection [130], regardless of whether the selection is centralized or decentralized, participatory or non-participatory.
Recently, gene editing has been introduced as a method to manipulate the genome free of the disadvantages of genetically modified (GM) crops, such as allergic reactions in humans and reduced nutrition, negative environmental impact by releasing toxins into the soil, the development of pest resistance, and the disruption of crop biodiversity [68]. A number of papers have been published describing how modifying one or a few genes via gene editing can result in substantial increases in crop yields ranging from 10% to 68% in crops such as wheat, rice, maize, tobacco, tomato, and soybean [131,132,133,134,135]. However, a concern has been raised because “many studies are the results of tests conducted in greenhouses or in small-scale field trials—the latter typically involving plants grown in small plots [136]. Few, if any, have used the experimental designs needed to evaluate crop performance in real-world environments. And hardly any findings have translated into yield increases on actual farms”. The paper [136] adds that, in the specific case of gene editing, the narrative ignores issues such as the quantitative nature of several traits, including disease resistance and the strong environmental effects on the most important traits. To these concerns, we may add the lack of precision of the technique and the role of epigenetics, which seems to be ignored in the discussion of gene editing.
Concern has also been expressed about the lack of social, political, and economic context in the narrative of using molecular tools [137].
It has been recognized that the most important limitation of genetically modified organisms, which is also common to conventionally bred varieties possessing major resistance genes [138,139] and also to the products of gene editing, and in general to monoculture, is that they represent a solution to the problem of biotic stress resistance that is within the evolutionary capabilities of the organism to be controlled, with several examples, such as the evolution of resistance to Bt maize by western corn rootworm [140] and the evolution of herbicide resistance in weeds [141]. Maintaining the performance of cultivars in monoculture requires an expensive input in genetic resources, plant breeding, and pesticide use [142]. Therefore, and in addition to the limitations described above, the products of gene editing are, like the GMOs and the uniform varieties, a nondurable solution to the problem of biotic stress resistance, which can break down at any moment, damaging particularly organic farmers. Furthermore, their genetic uniformity makes them incapable of responding to the complexity of climatic changes and associated changes in biotic stresses. On the contrary, the implementation of a breeding program, such as the one described above, which combines the advantages of decentralization and participation with the continuous flow of novel genetic materials, is the complex response needed to cope with a complex problem such as climate change.
Author Contributions
Conceptualization, S.C. and S.G.; methodology, S.C. and S.G.; software, S.C. and S.G.; validation, S.C. and S.G.; formal analysis, S.C. and S.G.; investigation, S.C. and S.G.; resources, S.C. and S.G.; data curation, S.C. and S.G.; writing—original draft preparation, S.C. and S.G.; writing—review and editing, S.C. and S.G.; visualization, S.C. and S.G.; supervision, S.C. and S.G.; project administration, S.C. and S.G.; funding acquisition, S.C. and S.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
No new data were created.
Acknowledgments
The authors thank Micaela Colley for editing the revised version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2023. In Urbanization, Agrifood Systems Transformation and Healthy Diets across the Rural–Urban Continuum; FAO: Rome, Italy, 2023. [Google Scholar] [CrossRef]
- Kumar, J.; Choudhary, A.K.; Gupta, D.S.; Kumar, S. Towards Exploitation of Adaptive Traits for Climate-Resilient Smart Pulses. Int. J. Mol. Sci. 2019, 20, 2971. [Google Scholar] [CrossRef]
- Razzaq, A.; Kaur, P.; Akhter, N.; Wani, S.H.; Saleem, F. Next-Generation Breeding Strategies for Climate-Ready Crops. Front. Plant Sci. 2021, 12, 620420. [Google Scholar] [CrossRef]
- Varotto, S.; Tani, E.; Abraham, E.; Krugman, T.; Kapazoglou, A.; Melzer, R.; Radanović, A.; Miladinović, D. Epigenetics: Possible applications in climate-smart crop breeding. J. Exp. Bot. 2020, 71, 5223–5236. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.R.; Skea, J.; Slade, R.; van Diemen, R.; Haughey, E.; Malley, J.; Pathak, M.; Portugal Pereira, J. Technical Summary. In Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P.R., Skea, J., Calvo Buendia, E., Masson-Delmotte, V., Pörtner, H.-O., Roberts, D.C., Zhai, P., Slade, R., Connors, S., van Diemen, R., et al., Eds.; Cambridge University Press: Cambridge, UK, 2019. [Google Scholar] [CrossRef]
- United Nations Framework Convention on Climate Change. Climate Change: Impacts, Vulnerabilities and Adaptation in Developing Countries; United Nations: New York, NY, USA, 2007; Available online: https://unfccc.int/resource/docs/publications/impacts.pdf (accessed on 28 December 2023).
- Kambach, S.; Sabatini, F.M.; Attorre, F.; Biurrun, I.; Boenisch, G.; Bonari, G.; Čarni, A.; Carranza, M.L.; Chiarucci, A.; Chytrý, M.; et al. Climate-trait relationships exhibit strong habitat specificity in plant communities across Europe. Nat. Commun. 2023, 14, 712. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, K.; Matloob, A.; Aslam, F.; Florentine, S.K. Weeds in a Changing Climate: Vulnerabilities, Consequences, and Implications for Future Weed Management. Front. Plant Sci. 2017, 8, 95. [Google Scholar] [CrossRef] [PubMed]
- Copernicus: September 2023—Unprecedented Temperature Anomalies. 2023 on Track to be the Warmest Year on Record. Available online: https://climate.copernicus.eu/copernicus-september-2023-unprecedented-temperature-anomalies (accessed on 15 November 2023).
- Langridge, P.; Braun, H.; Hulke, B.; Ober, E.; Prasanna, B.M. Breeding crops for climate resilience. Theor. Appl. Genet. 2021, 134, 1607–1611. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Reynolds, M.; Xu, Y. Climate change challenges plant breeding. Curr. Opin. Plant Biol. 2022, 70, 102308. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, S.; Chapman, S.; Mahop, M.T.; Deva, C.; Masamba, K.; Mwamahonje, A. Exploring assumptions in crop breeding for climate resilience: Opportunities and principles for integrating climate model projections. Clim. Chang. 2021, 164, 38. [Google Scholar] [CrossRef]
- Manners, R.; van Etten, J. Are agricultural researchers working on the right crops to enable food and nutrition security under future climates? Glob. Environ. Chang. 2018, 53, 182–194. [Google Scholar] [CrossRef]
- Fess, T.L.; Kotcon, J.B.; Benedito, V.A. Crop Breeding for Low Input Agriculture: A Sustainable Response to Feed a Growing World Population. Sustainability 2011, 3, 1742–1772. [Google Scholar] [CrossRef]
- Brumlop, S.; Reichenbecher, W.; Tappeser, B.; Finckh, M.R. What is the SMARTest way to breed plants and increase agrobiodiversity? Euphytica 2013, 194, 53–66. [Google Scholar] [CrossRef]
- Frankel, O.H. The development and maintenance of superior genetic stocks. Heredity 1950, 4, 89–102. [Google Scholar] [CrossRef]
- Keneni, G.; Bekele, E.; Imtiaz, M.; Dagne, K. Genetic Vulnerability of Modern Crop Cultivars: Causes, Mechanism and Remedies. Int. J. Plant Res. 2012, 2, 69–79. [Google Scholar] [CrossRef]
- Louwaars, N.P. Plant breeding and diversity: A troubled relationship? Euphytica 2018, 214, 114. [Google Scholar] [CrossRef]
- Hufford, M.B.; Berny Mier y Teran, J.C.; Gepts, P. Crop Biodiversity: An Unfinished Magnum Opus of Nature. Annu. Rev. Plant Biol. 2019, 70, 727–751. [Google Scholar] [CrossRef]
- Gepts, P. Biocultural diversity and crop improvement. Emerg. Top. Life Sci. 2023, 7, 151–196. [Google Scholar] [CrossRef]
- Barot, S.; Allard, V.; Cantarel, A.; Enjalbert, J.; Gauffreteau, A.; Goldringer, I.; Lata, J.-C.; Le Roux, X.; Niboyet, A.; Porcher, E. Designing mixtures of varieties for multifunctional agriculture with the help of ecology: A review. Agron. Sustain. Dev. 2017, 37, 13. [Google Scholar] [CrossRef]
- Burger, J.R.; Allen, C.D.; Brown, J.H.; Burnside, W.R.; Davidson, A.D.; Fristoe, T.S.; Hamilton, M.J.; Mercado-Silva, N.; Nekola, J.C.; Okie, J.G.; et al. The Macroecology of Sustainability. PLoS Biol. 2012, 10, e1001345. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D. The ecological consequences of changes in biodiversity: A search for general principles. Ecology 1999, 80, 1455–1474. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Wright, J.P.; Cadotte, M.W.; Carroll, I.T.; Hector, A.; Srivastava, D.S.; Loreau, M.; Weis, J.J. Impacts of plant diversity on biomass production increase through time because of species complementarity. Proc. Natl. Acad. Sci. USA 2007, 104, 18123–181288. [Google Scholar] [CrossRef]
- Gross, K.; Cardinale, B.J.; Fox, J.W.; Gonzalez, A.; Loreau, M.; Polley, H.W.; Reich, P.B.; van Ruijven, J. Species richness and the temporal stability of biomass production: A new analysis of recent biodiversity experiments. Am. Nat. 2013, 183, 1–12. [Google Scholar] [CrossRef]
- Renard, D.; Tilman, D. National food production stabilized by crop diversity. Nature 2019, 571, 257–260. [Google Scholar] [CrossRef]
- McCann, K.S. The diversity–stability debate. Nature 2000, 404, 228–233. [Google Scholar] [CrossRef] [PubMed]
- May, R.M. Stability and Complexity in Model Ecosystems; Princeton University Press: Princeton, NJ, USA, 2001. [Google Scholar] [CrossRef]
- Tamburini, G.; Bommarco, R.; Wanger, C.T.; Kremen, C.; van der Heijden, M.G.A.; Liebman, M.; Hallin, S. Agricultural diversification promotes multiple ecosystem services without compromising yield. Sci. Adv. 2020, 6, eaba1715. [Google Scholar] [CrossRef] [PubMed]
- MacLaren, C.; Mead, A.; van Balen, D.; Claessens, L.; Etana, A.; de Haan, J.; Haagsma, W.; Jäck, O.; Keller, T.; Labuschagne, J.; et al. Long-term evidence for ecological intensification as a pathway to sustainable agriculture. Nat. Sustain. 2022, 5, 770–779. [Google Scholar] [CrossRef]
- Nelson, K.S.; Burchfield, E.K. Defining features of diverse and productive agricultural systems: An archetype analysis of U.S. agricultural counties. Front. Sustain. Food Syst. 2023, 7, 1081079. Available online: https://api.semanticscholar.org/CorpusID:257301561 (accessed on 28 December 2023). [CrossRef]
- Balvanera, P.; Pfisterer, A.B.; Buchmann, N.; He, J.-S.; Nakashizuka, T.; Raffaelli, D.; Schmid, B. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol. Lett. 2006, 9, 1146–1156. [Google Scholar] [CrossRef]
- Cardinale, B.; Srivastava, D.; Duffy, E.; Justin, J.; Wright, A.L. Downing, Mahesh Sankaran & Claire Jouseau. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 2006, 443, 989–992. [Google Scholar] [CrossRef]
- Stachowicz, J.J.; Bruno, J.F.; Duffy, J.E. Understanding the Effects of Marine Biodiversity on Communities and Ecosystems. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 739–766. [Google Scholar] [CrossRef]
- von Hertzen, L.; Hanski, I.; Haahtela, T. Natural immunity: Biodiversity loss and inflammatory diseases are two global megatrends that might be related. EMBO Rep. 2011, 12, 1089–1093. [Google Scholar] [CrossRef]
- Khamsi, R. A gut feeling about immunity. Nat. Med. 2015, 21, 674–676. [Google Scholar] [CrossRef]
- Sibhatu, K.T.; Qaim, N. Review: Meta-analysis of the association between production diversity, diets, and nutrition in smallholder farm households. Food Policy 2018, 77, 1–18. [Google Scholar] [CrossRef]
- Jones, A.D. Critical review of the emerging research evidence on agricultural biodiversity, diet diversity, and nutritional status in low- and middle-income countries. Nutr. Rev. 2017, 75, 769–782. [Google Scholar] [CrossRef]
- Mulmi, P.; Masters, W.A.; Ghosh, S.; Namirembe, G.; Rajbhandary, R.; Manohar, S.; Shrestha, S.; West, K.P.; Webb, P. Household food production is positively associated with dietary diversity and intake of nutrient-dense foods for older preschool children in poorer families: Results from a nationally-representative survey in Nepal. PLoS ONE 2017, 12, e0186765. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiota and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef]
- Carter, M.M.; Olm, M.R.; Merrill, B.D.; Dahan, D.; Tripathi, S.; Spencer, S.P.; Yu, F.B.; Jain, S.; Neff, N.; Jha, A.R.; et al. Ultra-deep sequencing of Hadza hunter-gatherers recovers vanishing gut microbes. Cell 2023, 186, 3111–3124.e13. [Google Scholar] [CrossRef]
- Ritz, N.L.; Brocka, M.; Butler, M.I.; Cowan, C.S.M.; Barrera-Bugueño, C.; Turkington, C.J.R.; Draper, L.A.; Bastiaanssen, T.F.S.; Turpin, V.; Morales, L.; et al. Social anxiety disorder-associated gut microbiota increases social fear. Proc. Natl. Acad. Sci. USA 2024, 121, e2308706120. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yin, F.; Guo, Z.; Li, R.; Sun, W.; Wang, Y.; Geng, Y.; Sun, C.; Sun, D. Association between gut microbiota and glioblastoma: A Mendelian randomization study. Front. Genet. 2024, 14, 1308263. [Google Scholar] [CrossRef]
- Galván-Peña, S.; Zhu, Y.; Hanna, B.S.; Mathis, D.; Benoist, C. A dynamic atlas of immunocyte migration from the gut. Sci. Immunol. 2024, 9, eadi0672. [Google Scholar] [CrossRef] [PubMed]
- Heiman, M.L.; Greenway, F.L. A healthy gastrointestinal microbiome is dependent on dietary diversity. Mol. Metab. 2016, 5, 317–320. [Google Scholar] [CrossRef]
- Ceccarelli, S. Nurturing diversity in our guts and on our farms to reduce health risks and increase food system resilience. In Bioversity International, Agrobiodiversity Index Report: Risk and Resilience; Bioversity International: Rome, Italy, 2019; pp. 107–113. [Google Scholar]
- Willett, W.; Rockström, J.; Loken, B.; Springmann, M.; Lang, T.; Vermeulen, S.; Garnett, T.; Tilman, D.; DeClerck, F.; Wood, A.; et al. Food in the Anthropocene: The EAT–Lancet Commission on healthy diets from sustainable food systems. Lancet Comm. 2019, 393, 447–492. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, S.; Goldman, I.; Ortiz, R. Pursuing the Potential of Heirloom Cultivars to Improve Adaptation, Nutritional, and Culinary Features of Food Crops. Agronomy 2019, 9, 441. [Google Scholar] [CrossRef]
- Dinu, M.; Whittaker, A.; Pagliai, G.; Benedettelli, S.; Sofi, F. Ancient wheat species and human health: Biochemical and clinical implications. J. Nutr. Biochem. 2018, 52, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gotti, R.; Amadesi, E.; Fiori, J.; Bosi, S.; Bregola, V.; Marotti, I.; Dinelli, G. Differentiation of modern and ancient varieties of common wheat by quantitative capillary electrophoretic profile of phenolic acids. J. Chromatogr. A 2018, 1532, 208–215. [Google Scholar] [CrossRef]
- Sofi, F.; Dinu, M.; Pagliai, G.; Cei, L.; Sacchi, G.; Benedettelli, D.; Stefani, G.; Gagliardi, E.; Tosi, P.; Bocci, R.; et al. Health and Nutrition Studies Related to Cereal Biodiversity: A Participatory Multi-Actor Literature Review Approach. Nutrients 2018, 10, 1207. [Google Scholar] [CrossRef]
- Medina-Lozano, I.; Díaz, A. Nutritional Value and Phytochemical Content of Crop Landraces and Traditional Varieties. In Landraces; Elkelish, A., Ed.; IntechOpen: Rijeka, Croatia, 2021; pp. 1–22. [Google Scholar] [CrossRef]
- Kumar, N.; Harris, J.; Rawat, R. If They Grow It, Will They Eat and Grow? Evidence from Zambia on Agricultural Diversity and Child Undernutrition. J. Dev. Stud. 2015, 51, 1060–1077. [Google Scholar] [CrossRef]
- Saaka, M.; Osman, S.M.; Hoeschle-Zeledon, I. Relationship between agricultural biodiversity and dietary diversity of children aged 6–36 months in rural areas of Northern Ghana. Food Nutr. Res. 2017, 61, 1391668. [Google Scholar] [CrossRef]
- Luna-González, D.V.; Sørensen, M. Higher agrobiodiversity is associated with improved dietary diversity, but not child anthropometric status, of Mayan Achí people of Guatemala. Public Health Nutr. 2018, 21, 2128–2141. [Google Scholar] [CrossRef]
- Tollefson, J. Why deforestation and extinctions make pandemics more likely. Nature 2020, 584, 175. [Google Scholar] [CrossRef]
- Schnell, F.W. A synoptic study of the methods and categories of plant breeding. Z. Pflanzenzüchtung 1982, 89, 1–18. [Google Scholar]
- Galluzzi, G.; Seyoum, A.; Halewood, M.; López Noriega, I.; Welch, E.W. The Role of Genetic Resources in Breeding for Climate Change: The Case of Public Breeding Programmes in Eighteen Developing Countries. Plants 2020, 9, 1129. [Google Scholar] [CrossRef]
- Ceccarelli, S. Main stages of a plant breeding programme. In Plant Breeding and Farmer Participation; Ceccarelli, S., Guimaraes, E.P., Weltzien, E., Eds.; FAO: Rome, Italy, 2009; pp. 63–74. [Google Scholar]
- Venuprasad, R.; Lafitte, H.R.; Atlin, G.N. Response to Direct Selection for Grain Yield under Drought Stress in Rice. Crop Sci. 2007, 47, 285–293. [Google Scholar] [CrossRef]
- Barah, B.C.; Binswanger, H.P.; Rana, B.S.; Rao, N.G.P. The use of risk aversion in plant breeding: Concept and application. Euphytica 1981, 30, 451–458. [Google Scholar] [CrossRef]
- Lin, C.S.; Binns, M.R. A method of analyzing cultivar × location × year experiments: A new stability parameter. Theor. Appl. Genet. 1988, 76, 425–430. [Google Scholar] [CrossRef]
- Evans, L.T. Crop Evolution, Adaptation and Yield; Cambridge University Press: New York, NY, USA, 1993. [Google Scholar]
- Baranski, M.R. The Globalization of Wheat: A Critical History of the Green Revolution; University of Pittsburgh Press: Pittsburgh, PA, USA, 2022; p. 247. [Google Scholar]
- Ceccarelli, S. Wide Adaptation. How Wide? Euphytica 1989, 40, 197–205. [Google Scholar] [CrossRef]
- Hammer, K.; Knüpffer, H.; Xhuveli, L.; Perrino, P. Estimating genetic erosion in landraces—Two case studies. Genet. Resour. Crop Evol. 1996, 43, 329–336. [Google Scholar] [CrossRef]
- Eriksson, D. The evolving EU regulatory framework for precision breeding. Theor. Appl. Genet. 2019, 132, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Yadav, A.; Ahmad, R.; Dwivedi, U.N.; Yadav, K. CRISPR-Based Genome Editing for Nutrient Enrichment in Crops: A Promising Approach Toward Global Food Security. Front. Genet. 2022, 13, 932859. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, S.; Grando, S.; Bailey, E.; Amri, A.; El Felah, M. Farmer Participation in Barley Breeding in Syria, Morocco and Tunisia. Euphytica 2001, 122, 521–536. [Google Scholar] [CrossRef]
- Bellon, M.R.; Morris, M.L. Linking Global and Local Approaches to Agricultural Technology Development: The Role of Participatory Plant Breeding Research in the CGIAR; Economic Working Paper 02–03; International Maize and Wheat Improvement Center: El Batan, Mexico, 2003. [Google Scholar]
- Colley, M.R.; Dawson, J.C.; McCluskey, C.; Myers, J.R.; Tracy, W.F.; Lammerts van Bueren, E.T. Exploring the emergence of participatory plant breeding in countries of the Global North—A review. J. Agric. Sci. 2021, 159, 320–338. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Grando, S.; Tutwiler, R.; Baha, J.; Martini, A.M. A Methodological Study on Participatory Barley Breeding. I. Selection Phase. Euphytica 2000, 111, 91–104. [Google Scholar] [CrossRef]
- Ceccarelli, S. Efficiency of plant breeding. Crop Sci. 2015, 55, 87–97. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Grando, S. Participatory Plant Breeding: Who did it, who does it and where? Exp. Agric. 2020, 56, 1–11. [Google Scholar] [CrossRef]
- Shelton, A.C.; Tracy, W.F. Recurrent Selection and Participatory Plant Breeding for Improvement of Two Organic Open-Pollinated Sweet Corn (Zea mays L.) Populations. Sustainability 2015, 7, 5139–5152. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Grando, S. Organic agriculture and evolutionary populations to merge mitigation and adaptation strategies to fight climate change. South Sustain. 2020, 1, e002. [Google Scholar] [CrossRef]
- Descheemaeker, K.; Ronner, E.; Ollenburger, M.; Franke, A.; Klapwijk, C.; Falconnier, G.N.; Wichern, J.; Giller, K. Which Options Fit Best? Operationalizing The Socio-Ecological Niche Concept. Exp. Agric. 2019, 55, 169–190. [Google Scholar] [CrossRef]
- Suneson, C.A. An Evolutionary Plant Breeding Method. Agron. J. 1956, 48, 188–191. [Google Scholar] [CrossRef]
- Harlan, H.V.; Martini, M.L. A composite hybrid mixture. J. Am. Soc. Agron. 1929, 21, 487–490. [Google Scholar] [CrossRef]
- Wolfe, M.S.; Ceccarelli, S. The need to use more diversity in cereal cropping requires more descriptive precision. J. Sci. Food Agric. 2020, 100, 4119–4123. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, S.; Grando, S. Evolutionary Plant Breeding with an Introduction to Participatory Plant Breeding; Mimesis Edizioni Srl: Milano, Italy, 2022; 175p, Available online: https://archive.org/details/evolutionary-plant-breeding (accessed on 28 December 2023).
- Allard, R.W.; Jain, S.K. Population Studies in Predominantly Self-Pollinated Species. II. Analysis of Quantitative Genetic Changes in a Bulk-Hybrid Population of Barley. Evolution 1962, 16, 90–101. [Google Scholar] [CrossRef]
- Rhoné, B.; Vitalis, R.; Goldringer, I.; Bonnin, I. Evolution of flowering time in experimental wheat populations: A comprehensive approach to detect genetic signatures of natural selection. Evolution 2010, 64, 2110–2125. [Google Scholar] [CrossRef] [PubMed]
- Allard, R.W.; Hansche, P.E. Some parameters of population variability and their implications in plant breeding. Adv. Agron. 1964, 16, 281–325. [Google Scholar] [CrossRef]
- Patel, J.D.; Reinbergs, E.; Mather, D.E.; Choo, T.M.; Sterling, J.D.E. Natural selection in a double-haploid mixture and a composite cross of barley. Crop Sci. 1987, 27, 474–479. [Google Scholar] [CrossRef]
- Rasmusson, D.C.; Beard, B.H.; Johnson, F.K. Effect of Natural Selection on Performance of a Barley Population. Crop Sci. 1967, 7, 543. [Google Scholar] [CrossRef]
- Soliman, K.M.; Allard, R.W. Grain Yield of Composite Cross Populations of Barley: Effects of Natural Selection. Crop Sci. 1991, 31, 705–708. [Google Scholar] [CrossRef]
- Raggi, L.; Ciancaleoni, S.; Torricelli, R.; Terzi, V.; Ceccarelli, S.; Negri, V. Evolutionary breeding for sustainable agriculture: Selection and multi-environmental evaluation of barley populations and lines. Field Crops Res. 2017, 204, 76–88. [Google Scholar] [CrossRef]
- Goldringer, I.; Enjalbert, J.; David, J.; Paillard, S.; Pham, J.; Brabant, P. Dynamic management of genetic resources: A 13-year experiment on wheat. In Broadening the Genetic Base of Crop Production; Cooper, H., Spillane, C., Hodgkin, T., Eds.; CABI: Wallingford, UK, 2001; pp. 245–260. [Google Scholar]
- Baresel, J.P.; Bülow, L.; Finckh, M.R.; Frese, L.; Knapp, S.; Schmidhalter, U.; Weedon, O.D. Performance and evolutionary adaptation of heterogeneous wheat populations. Euphytica 2022, 218, 137. [Google Scholar] [CrossRef]
- Allard, R.W. Relationship Between Genetic Diversity and Consistency of Performance in Different Environments. Crop Sci. 1961, 1, 127–133. [Google Scholar] [CrossRef]
- Kiær, L.P.; Skovgaard, I.M.; Østergård, H. Effects of inter-varietal diversity, biotic stresses and environmental productivity on grain yield of spring barley variety mixtures. Euphytica 2012, 185, 123–138. [Google Scholar] [CrossRef]
- Brumlop, S.; Pfeiffer, T.; Finckh, M. Evolutionary Effects on Morphology and Agronomic Performance of Three Winter Wheat Composite Cross Populations Maintained for Six Years under Organic and Conventional Conditions. Org. Farming 2017, 3, 34–50. [Google Scholar] [CrossRef]
- Weedon, O.D.; Finckh, M.R. Heterogeneous Winter Wheat Populations Differ in Yield Stability Depending on their Genetic Background and Management System. Sustainability 2019, 11, 6172. [Google Scholar] [CrossRef]
- Weedon, O.D.; Finckh, M.R. Response of Wheat Composite Cross Populations to Disease and Climate Variation Over 13 Generations. Front. Agric. Sci. Eng. 2021, 8, 400–415. [Google Scholar]
- Weedon, O.D.; Brumlop, S.; Haak, A.; Baresel, J.P.; Borgen, A.; Döring, T.; Goldringer, I.; Lammerts van Bueren, E.; Messmer, M.M.; Mikó, P.; et al. High Buffering Potential of Winter Wheat Composite Cross Populations to Rapidly Changing Environmental Conditions. Agronomy 2023, 13, 1662. [Google Scholar] [CrossRef]
- Frey, K.J.; Maldonado, U. Relative Productivity of Homogeneous and Heterogeneous Oat Cultivars in Optimum and Suboptimum Environments. Crop Sci. 1967, 7, 532–535. [Google Scholar] [CrossRef]
- Dubin, H.J.; Wolfe, M.S. Comparative behavior of three wheat cultivars and their mixture in India, Nepal and Pakistan. Field Crops Res. 1994, 39, 71–83. [Google Scholar] [CrossRef]
- Merrick, L.F.; Lyon, S.R.; Balow, K.A.; Murphy, K.M.; Jones, S.S.; Carter, A.H. Utilization of evolutionary plant breeding increases stability and adaptation of winter wheat across diverse precipitation zones. Sustainability 2020, 12, 9728. [Google Scholar] [CrossRef]
- van Frank, G.; Rivière, P.; Pin, S.; Baltassat, R.; Berthellot, J.-F.; Caizergues, F.; Dalmasso, C.; Gascuel, J.-S.; Hyacinthe, A.; Mercier, F.; et al. Genetic Diversity and Stability of Performance of Wheat Population Varieties Developed by Participatory Breeding. Sustainability 2020, 12, 384. [Google Scholar] [CrossRef]
- Salimi, M.; Razavi, K.C.; Amiri, M.N.; Esmaeili, M.; Khorramdel, S.; Moghani, H.; Grando, S.; Ceccarelli, S. Stability of Agronomic Traits of Barley Evolutionary Populations under Drought Conditions in Iran. Agronomy 2023, 13, 1931. [Google Scholar] [CrossRef]
- Simmonds, N.W. Variability in Crop Plants, Its Use and Conservation. Biol. Rev. 1962, 37, 422–465. [Google Scholar] [CrossRef]
- Smithson, J.B.; Lenné, J.M. Varietal mixtures: A viable strategy for sustainable productivity in subsistence agriculture. Ann. Appl. Biol. 1996, 128, 127–158. [Google Scholar] [CrossRef]
- Mundt, C.C. Use of multiline cultivars and cultivar mixtures for disease management. Annu. Rev. Phytopathol. 2002, 40, 381–410. [Google Scholar] [CrossRef]
- Jackson, L.F.; Webster, R.K.; Allard, R.W.; Kahler, A.L. Genetic Analysis of Changes in Scald Resistance in Barley Composite Cross V. Phytopathology 1982, 72, 1069–1072. [Google Scholar] [CrossRef]
- Allard, R.W. The Genetics of Host-Pathogen Coevolution: Implications for Genetic Resource Conservation. J. Hered. 1990, 81, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, K.M.; Barrett, J.A. Evolution of mildew resistance in a hybrid bulk population of barley. Heredity 1991, 67, 247–256. [Google Scholar] [CrossRef]
- McDonald, B.A.; Allard, R.W.; Webster, R.K. Responses of Two, Three, and Four Component Barley Mixtures to a Variable Pathogen Population. Crop Sci. 1988, 28, 447–452. [Google Scholar] [CrossRef]
- Finckh, M.R.; Gacek, E.; Goyeau, H.; Lannou, C.; Merz, U.; Mundt, C.C.; Munk, L.; Nadziak, L.; Newton, A.C.; de Vallavieille-Popec, C.; et al. Cereal variety and species mixtures in practice, with emphasis on disease resistance. Agron. EDP Sci. 2000, 20, 813–837. [Google Scholar] [CrossRef]
- Finckh, M.R.; Wolfe, M. Diversification strategies. In The Epidemiology of Plant Diseases; Cooke, B., Jones, D., Kaye, B., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 269–307. [Google Scholar] [CrossRef]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; Methuen: London, UK, 1958. [Google Scholar]
- Crutsinger, G.M.; Souza, L.; Sanders, N.J. Intraspecific diversity and dominant genotypes resist plant invasions. Ecol. Lett. 2008, 11, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Lemerle, D.; Gill, G.S.; Murphy, C.E.; Walker, S.R.; Cousens, R.D.; Mokhtari, S.; Peltzer, S.J.; Coleman, R.; Luckett, D.J. Genetic improvement and agronomy for enhanced wheat competitiveness with weeds. Aust. J. Agric. Res. 2001, 52, 527–548. [Google Scholar] [CrossRef]
- Bertholdsson, N.O. Early vigour and allelopathy—Two useful traits for enhanced barley and wheat competitiveness against weeds. Weed Res. 2005, 45, 94–102. [Google Scholar] [CrossRef]
- Lazzaro, M.; Costanzo, A.; Farag, D.H.; Bàrberi, P. Grain yield and competitive ability against weeds in modern and heritage common wheat cultivars are differently influenced by sowing density. Ital. J. Agron. 2017, 12, 343–349. [Google Scholar] [CrossRef]
- Hoad, S.P.; Bertholdsson, N.O.; Neuhoff, D.; Köpke, U. Approaches to Breed for Improved Weed Suppression in Organically Grown Cereals. In Organic Crop Breeding; Lammerts van Bueren, E., Myers, J.R., Eds.; Wiley-Blackwell: Chichester, UK; Ames, IA, USA, 2012; pp. 61–76. [Google Scholar] [CrossRef]
- Hockett, E.A.; Eslick, R.F.; Qualset, C.O.; Dubbs, A.L.; Stewart, V.R. Effects of Natural Selection in Advanced Generations of Barley Composite Cross II. Crop Sci. 1983, 23, 752–756. [Google Scholar] [CrossRef]
- Goldringer, I.; Enjalbert, J.; Raquin, A.-L.; Brabant, P. Strong selection in wheat populations during ten generations of dynamic management. Genet. Sel. Evol. 2001, 33 (Suppl. S1), S441–S463. [Google Scholar] [CrossRef]
- Knapp, S.; Döring, T.F.; Jones, H.E.; Snape, J.; Wingen, L.U.; Wolfe, M.S.; Leverington-Waite, M.; Griffiths, S. Natural Selection Towards Wild-Type in Composite Cross Populations of Winter Wheat. Front. Plant Sci. 2020, 10, 1757. [Google Scholar] [CrossRef]
- Wright, A.J.; de Kroon, H.; Visser, E.J.W.; Buchmann, T.; Ebeling, A.; Eisenhauer, N.; Fischer, C.; Hildebrandt, A.; Ravenek, J.; Roscher, C.; et al. Plants are less negatively affected by flooding when growing in species-rich plant communities. New Phytol. 2017, 213, 645–656. [Google Scholar] [CrossRef]
- Wolfe, M.S.; Brändle, U.; Koller, B.; Limpert, E.; McDermott, J.M.; Müller, K.; Schaffner, D. Barley mildew in Europe: Population biology and host resistance. Euphytica 1992, 63, 125–139. [Google Scholar] [CrossRef]
- Winge, T. Seed Legislation in Europe and Crop Genetic Diversity. In Sustainable Agriculture Reviews; Lichtfouse, E., Ed.; Springer: Cham, Switzerland, 2015; Volume 15. [Google Scholar]
- Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32014D0150 (accessed on 29 November 2023).
- Available online: https://food.ec.europa.eu/system/files/2022-04/prm_temp-exp_pop-exp_en.pdf (accessed on 28 January 2024).
- Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32018R0848 (accessed on 29 November 2023).
- Ceccarelli, S. Plant Breeding with Farmers—A Technical Manual; ICARDA: Aleppo, Syria, 2012; pp. xi + 126. [Google Scholar]
- Neupane, S.P.; Joshi, B.K.; Ayer, D.K.; Ghimire, K.H.; Gauchan, D.; Karkee, A.; Jarvis, D.J.; Mengistu, D.K.; Grando, S.; Ceccarelli, S. Farmers’ Preferences and Agronomic Evaluation of Dynamic Mixtures of Rice and Bean in Nepal. Diversity 2023, 15, 660. [Google Scholar] [CrossRef]
- Cobb, J.N.; Juma, R.U.; Biswas, P.S.; Arbelaez, J.D.; Rutkoski, J.; Atlin, G.; Hagen, T.; Quinn, M.; Ng, E.H. Enhancing the rate of genetic gain in public-sector plant breeding programs: Lessons from the breeder’s equation. Theor. Appl. Genet. 2019, 132, 627–645. [Google Scholar] [CrossRef] [PubMed]
- Fadda, C.; Mengistu, D.J.; Kidane, Y.G.; Dell’Acqua, M.; Pè, M.E.; Van Etten, J. Integrating Conventional and Participatory Crop Improvement for Smallholder Agriculture Using the Seeds for Needs Approach: A Review. Front. Plant Sci. 2020, 11, 559515. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, P.; Zou, C.; Lu, Y.; Xie, C.; Zhang, X.; Prasanna, B.M.; Olsen, M.S. Enhancing genetic gain in the era of molecular breeding. J. Exp. Bot. 2017, 68, 2641–2666. [Google Scholar] [CrossRef]
- De Souza, A.P.; Burgess, S.J.; Doran, L.; Hansen, J.; Manukyan, L.; Maryn, N.; Gotarkar, D.; Leonelli, L.; Niyogi, K.K.; Long, S.P. Soybean photosynthesis and crop yield are improved by accelerating recovery from photoprotection. Science 2022, 377, 851–854. [Google Scholar] [CrossRef]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Li, X.; Lu, Z.; Zhang, H.; Ye, X.; Zhou, Y.; Li, J.; Yan, Y.; Pei, H.; Duan, F.; et al. A transcriptional regulator that boosts grain yields and shortens the growth duration of rice. Science 2022, 377, eabi8455. [Google Scholar] [CrossRef]
- Yu, Q.; Liu, S.; Yu, L.; Xiao, Y.; Zhang, S.; Wang, X.; Xu, Y.; Yu, H.; Li, Y.; Yang, J.; et al. RNA demethylation increases the yield and biomass of rice and potato plants in field trials. Nat. Biotechnol. 2021, 39, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Lawit, S.J.; Weers, B.; Sun, J.; Mongar, N.; Van Hemert, J.; Melo, R.; Meng, X.; Rupe, M.; Clapp, J.; et al. Overexpression of zmm28 increases maize grain yield in the field. Proc. Natl. Acad. Sci. USA 2019, 116, 23850–23858. [Google Scholar] [CrossRef]
- Khaipho-Burch, M.; Cooper, M.; Crossa, J.; de Leon, N.; Holland, J.; Lewis, R.; McCouch, S.; Murray, S.C.; Rabbi, I.; Ronald, P.; et al. Scale up trials to validate modified crops’ benefits. Nature 2023, 621, 470–473. [Google Scholar] [CrossRef]
- Fischer, K.; Rock, J.S. The scientific narrative around new food technologies needs to change. Nat. Rev. Bioeng. 2023, 1, 786–787. [Google Scholar] [CrossRef]
- Robinson, R.A. Return to Resistance—Breeding Crops to Reduce Pesticide Dependence; agAccess: Davis, CA, USA, 1996; 480p, ISBN 0-932857-17-5. [Google Scholar]
- Wolfe, M.S. Crop strength through diversity. Nature 2000, 406, 681–682. [Google Scholar] [CrossRef]
- Ceccarelli, S. GMO, Organic Agriculture and Breeding for Sustainability. Sustainability 2014, 6, 4273–4286. [Google Scholar] [CrossRef]
- Vencill, W.; Nichols, R.; Webster, T.; Soteres, J.; Mallory-Smith, C.; Burgos, N.; Johnson, W.G.; McClelland, M. Herbicide Resistance: Toward an Understanding of Resistance Development and the Impact of Herbicide-Resistant Crops. Weed Sci. 2012, 60, 2–30. [Google Scholar] [CrossRef]
- Wolfe, M.S.; Finckh, M.R. Diversity of host resistance within the crop: Effects on host, pathogen and disease. In Plant Resistance to Fungal Diseases; Hartleb, H., Heitefuss, R., Hoppe, H.H., Eds.; G. Fischer Verlag: Jena, Germany, 1997; pp. 378–400. [Google Scholar]
Figure 1.
The stages of a plant breeding program.

Figure 2.
A participatory evolutionary breeding program: in path 1, the evolutionary population evolves as it is planted and harvested in each of many sites as the farmer’s crop (here, only one is shown as an example). Path 2 is a single spike (or panicles, or cobs, or pods) selection, which can be followed by head rows (paths 3 and 4) or by the creation of sub-populations (paths 4 and 5).
Figure 2.
A participatory evolutionary breeding program: in path 1, the evolutionary population evolves as it is planted and harvested in each of many sites as the farmer’s crop (here, only one is shown as an example). Path 2 is a single spike (or panicles, or cobs, or pods) selection, which can be followed by head rows (paths 3 and 4) or by the creation of sub-populations (paths 4 and 5).

Figure 3.
Means over three seasons of the flowering date, maturity date, plant height, and number of tillers per plant of four rice mixtures, a local variety, a modern variety, and the selections made by male farmers (Msel), female farmers (Fsel), and technicians (Tsel) within mixture 4 (Mix4). The columns that have letters in common are not statistically (p < 0.05) different based on the L.S.D. test (modified from [127]).
Figure 3.
Means over three seasons of the flowering date, maturity date, plant height, and number of tillers per plant of four rice mixtures, a local variety, a modern variety, and the selections made by male farmers (Msel), female farmers (Fsel), and technicians (Tsel) within mixture 4 (Mix4). The columns that have letters in common are not statistically (p < 0.05) different based on the L.S.D. test (modified from [127]).

Figure 4.
Means over three seasons of a number of panicles/plants, panicle length, 1000 seed weight, and grain yield of four rice mixtures, a local variety, a modern variety, and the selections made by male farmers (Msel), female farmers (Fsel), and technicians (Tsel) within mixture 4 (Mix4). The columns that have letters in common are not statistically (p < 0.05) different based on the L.S.D. test (modified from [127]).
Figure 4.
Means over three seasons of a number of panicles/plants, panicle length, 1000 seed weight, and grain yield of four rice mixtures, a local variety, a modern variety, and the selections made by male farmers (Msel), female farmers (Fsel), and technicians (Tsel) within mixture 4 (Mix4). The columns that have letters in common are not statistically (p < 0.05) different based on the L.S.D. test (modified from [127]).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ceccarelli, S.; Grando, S. Diversity as a Plant Breeding Objective. Agronomy 2024, 14, 550. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030550
AMA Style
Ceccarelli S, Grando S. Diversity as a Plant Breeding Objective. Agronomy. 2024; 14(3):550. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030550
Chicago/Turabian StyleCeccarelli, Salvatore, and Stefania Grando. 2024. "Diversity as a Plant Breeding Objective" Agronomy 14, no. 3: 550. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030550
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.