
The Enzyme Activity of Dual-Domain β-Propeller Alkaline Phytase as a Potential Factor in Improving Soil Phosphorus Fertility and Triticum aestivum Growth
Abstract
:1. Introduction
2. Materials and Methods
2.1. PhyHT Construction, Expression, and Purification
2.2. Analysis of Phytase Properties
2.3. Soil Samples and Their Chemical Properties, Organic Fertilizer Composition, and Crop Residue Samples
2.4. Factors Affecting Phytase Activity of phyHT in Soil
2.5. Effect of phyHT on Available P Content in Soil
2.6. Effect of phyHT on Soil Physical and Chemical Properties
2.7. Effect of phyHT on the Available P Content in the Samples of Crop Residues and Organic Fertilizer
2.8. Effect of phyHT on Wheat Growth
2.9. Data and Statistical Analyses
3. Results and Discussion
3.1. The Implications for Temporal Stability and Resistance to Microbial Degradation of phyHT in Soil
3.2. Effect of Temperature and pH on Inoculated phyHT Activity in Soil
3.3. Efficacy of phyHT in Hydrolysis of Phytate in Soil
3.4. Effects of phyHT on Soil Physical and Chemical Properties
3.5. Effectiveness of phyHT for Hydrolysis of Phytate in Crop Residues and Organic Fertilizer
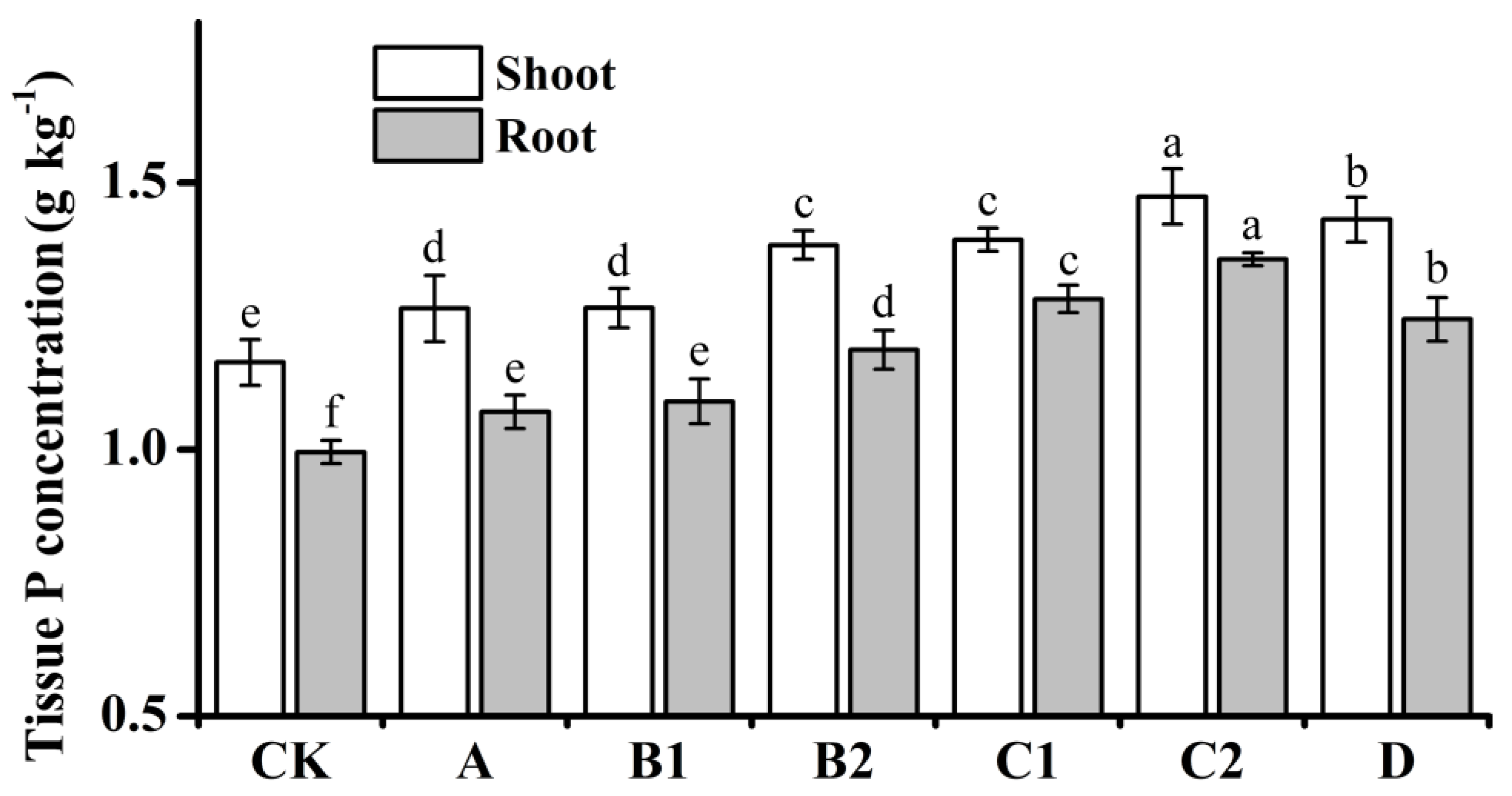
3.6. Effect of phyHT on Plant Growth
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hillman, J.R. Food, Energy, and Society. Third Edition. Edited by D. Pimentel and M. H. Pimentel. Boca Raton, Fl, USA: CRC Press/Taylor and Francis Group (2008), pp. 380, £48.99. ISBN 978-1-4200-4667-0. Ex. Agric. 2009, 45, 128–129. [Google Scholar] [CrossRef]
- Ramesh, A.; Sharma, S.K.; Joshi, O.P.; Khan, I.R. Phytase, Phosphatase Activity and P-Nutrition of Soybean as Influenced by Inoculation of Bacillus. Indian J. Microbiol. 2011, 51, 94–99. [Google Scholar] [CrossRef]
- Gerke, J. Phytate (Inositol Hexakisphosphate) in Soil and Phosphate Acquisition from Inositol Phosphates by Higher Plants. A Review. Plants 2015, 4, 253–266. [Google Scholar] [CrossRef]
- Liu, X.; Han, R.; Cao, Y.; Turner, B.L.; Ma, L.Q. Enhancing Phytate Availability in Soils and Phytate-P Acquisition by Plants: A Review. Environ. Sci. Technol. 2022, 56, 9196–9219. [Google Scholar] [CrossRef]
- Celi, L.; Barberis, E.; Turner, B.L.; Frossard, E.; Baldwin, D.S. Abiotic Stabilization of Organic Phosphorus in the Environment. In Organic Phosphorus in the Environment; CABI: Wallingford, UK, 2005; pp. 113–132. [Google Scholar] [CrossRef]
- Balwani, I.; Chakravarty, K.; Gaur, S. Role of Phytase Producing Microorganisms towards Agricultural Sustainability. Biocatal. Agric. Biotechnol. 2017, 12, 23–29. [Google Scholar] [CrossRef]
- Jain, J.; Sapna; Singh, B. Characteristics and Biotechnological Applications of Bacterial Phytases. Process Biochem. 2016, 51, 159–169. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2017, 54, 655–670. [Google Scholar] [CrossRef]
- George, T.S.; Richardson, A.E.; Simpson, R.J. Behaviour of Plant-derived Extracellular Phytase upon Addition to Soil. Soil Biol. Biochem. 2005, 37, 977–988. [Google Scholar] [CrossRef]
- Kedi, B.; Sei, J.; Quiquampoix, H.; Staunton, S. Persistence of Catalytic Activity of Fungal Phosphatases Incubated in Tropical Soils. Soil Biol. Biochem. 2013, 56, 69–74. [Google Scholar] [CrossRef]
- Giaveno, C.; Celi, L.; Richardson, A.E.; Simpson, R.J.; Barberis, E. Interaction of Phytases with Minerals and Availability of Substrate Affect the Hydrolysis of Inositol Phosphates. Soil Biol. Biochem. 2010, 42, 491–498. [Google Scholar] [CrossRef]
- Sun, D.; Zhang, W.; Feng, H.; Li, X.; Han, R.; Turner, B.L.; Qiu, R.; Cao, Y.; Ma, L.Q. Novel Phytase PvPHY1 from the As-Hyperaccumulator Pteris Vittata Enhances P Uptake and Phytate Hydrolysis, and Inhibits As Translocation in Plant. J. Hazard. Mater. 2022, 423, 127106. [Google Scholar] [CrossRef]
- Giles, C.D.; Hsu, P.-C.; Richardson, A.E.; Hurst, M.R.H.; Hill, J.E. Plant Assimilation of Phosphorus from an Insoluble Organic Form Is Improved by Addition of an Organic Anion Producing Pseudomonas Sp. Soil Biol. Biochem. 2014, 68, 263–269. [Google Scholar] [CrossRef]
- Singh, B.; Satyanarayana, T. Microbial Phytases in Phosphorus Acquisition and Plant Growth Promotion. Physiol. Mol. Biol. Plants 2011, 17, 93–103. [Google Scholar] [CrossRef]
- Singh, B.; Boukhris, I.; Pragya; Kumar, V.; Yadav, A.N.; Farhat-Khemakhem, A.; Kumar, A.; Singh, D.; Blibech, M.; Chouayekh, H. Contribution of Microbial Phytases to the Improvement of Plant Growth and Nutrition: A Review. Pedosphere 2020, 30, 295–313. [Google Scholar] [CrossRef]
- Kumar, V.; Yadav, A.N.; Verma, P.; Sangwan, P.; Saxena, A.; Kumar, K.; Singh, B. β-Propeller Phytases: Diversity, Catalytic Attributes, Current Developments and Potential Biotechnological Applications. Int. J. Biol. Macromol. 2017, 98, 595–609. [Google Scholar] [CrossRef]
- Cheng, C.; Lim, B.L. Beta-Propeller Phytases in the Aquatic Environment. Arch. Microbiol. 2006, 185, 1–13. [Google Scholar] [CrossRef]
- Lu, F.; Zhang, B.; Liu, Y.; Song, Y.; Guo, G.; Yang, P.; Yao, B.; Guo, S.; Gao, W. Sequencing and characterization of dual-domain β-propeller alkaline phytase. Amic. Sin. 2018, 58, 1582–1592. [Google Scholar] [CrossRef]
- Lu, F.; Guo, G.; Li, Q.; Feng, D.; Liu, Y.; Huang, H.; Yang, P.; Gao, W.; Yao, B. Preparation, Purification, Crystallization and Preliminary Crystallographic Analysis of Dual-Domain β-Propeller Phytase from Bacillus Sp. HJB17. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2014, 70, 1671–1674. [Google Scholar] [CrossRef]
- Gao, W.; Lu, F.; Li, Q.; Guo, G.C.N. A kind of method for largely preparing difunctional domain β-pleated sheet bucket phytase disome. Patent 103740672B, 14 July 2017. [Google Scholar]
- Qvirist, L.; Carlsson, N.; Andlid, T. Assessing Phytase Activity–Methods, Definitions and Pitfalls. J. Biol. Methods 2015, 2, e16. [Google Scholar] [CrossRef]
- Li, Y.; Fang, F.; Wei, J.; Wu, X.; Cui, R.; Li, G.; Zheng, F.; Tan, D. Humic Acid Fertilizer Improved Soil Properties and Soil Microbial Diversity of Continuous Cropping Peanut: A Three-Year Experiment. Sci. Rep. 2019, 9, 12014. [Google Scholar] [CrossRef]
- Menezes-Blackburn, D.; Jorquera, M.; Gianfreda, L.; Rao, M.; Greiner, R.; Garrido, E.; De La Luz Mora, M. Activity Stabilization of Aspergillus niger and Escherichia coli Phytases Immobilized on Allophanic Synthetic Compounds and Montmorillonite Nanoclays. Bioresour. Technol. 2011, 102, 9360–9367. [Google Scholar] [CrossRef]
- Gujar, P.D.; Bhavsar, K.P.; Khire, J.M. Effect of Phytase from Aspergillus Niger on Plant Growth and Mineral Assimilation in Wheat (Triticum aestivum Linn.) and Its Potential for Use as a Soil Amendment. J. Sci. Food Agric. 2013, 93, 2242–2247. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Z.; Liu, X.; Huang, G.; Xiao, W.; Han, L. The Composition Characteristics of Different Crop Straw Types and Their Multivariate Analysis and Comparison. Waste Manag. 2020, 110, 87–97. [Google Scholar] [CrossRef]
- Kumar, V.; Singh, P.; Jorquera, M.A.; Sangwan, P.; Kumar, P.; Verma, A.K.; Agrawal, S. Isolation of Phytase-Producing Bacteria from Himalayan Soils and Their Effect on Growth and Phosphorus Uptake of Indian Mustard (Brassica juncea). World J. Microbiol. Biotechnol. 2013, 29, 1361–1369. [Google Scholar] [CrossRef]
- Singh, P.; Kumar, V.; Agrawal, S. Evaluation of Phytase Producing Bacteria for Their Plant Growth Promoting Activities. Int. J. Microbiol. 2014, 2014, 426483. [Google Scholar] [CrossRef]
- Hu, X.; Roberts, D.P.; Xie, L.; Maul, J.E.; Yu, C.; Li, Y.; Zhang, S.; Liao, X. Development of a Biologically Based Fertilizer, Incorporating Bacillus megaterium A6, for Improved Phosphorus Nutrition of Oilseed Rape. Can. J. Microbiol. 2013, 59, 231–236. [Google Scholar] [CrossRef]
- Liu, L.; Du, W.; Luo, W.; Su, Y.; Hui, J.; Ma, S. Development of an Engineered Soil Bacterium Enabling to Convert Both Insoluble Inorganic and Organic Phosphate into Plant Available Phosphate and Its Use as a Biofertilizer. Mol. Biotechnol. 2015, 57, 419–429. [Google Scholar] [CrossRef]
- Meng, Y.; Zhang, J.; Zhao, Q. Research Progress on the Application of Plant Phytase in Feed. Feed. Ind. 2009, 30, 16–18. [Google Scholar]
- Yang, X.; Cao, X.; Wu, B.; Xu, Q.; Liu, X. Activity of Microbial Phytase in Soil and Its Improvement Method and Application. MB China 2023, 50, 2687–2708. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | phyHT Treatment |
|---|---|---|
| pH (1:2.5) | 8.78 | 8.16 ** |
| EC (a) (dS m−1) | 0.29 | 0.68 ** |
| OM (b) (g kg−1) | 38.78 | 42.87 * |
| Available N (mg kg−1) | 78.59 | 93.04 ** |
| Available P (mg kg−1) | 17.98 | 22.03 ** |
| Available K (mg kg−1) | 122.49 | 156.17 * |
| CaCO3 (%) | 13.67 | 10.12 * |
| Phytate (mg kg−1) | 597.56 | 495.98 ** |
| phytase activity (mg P2O5 (g×d)−1) | 0.25 | 0.61 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Li, J.; Lu, F.; Wang, S.; Ren, Y.; Guo, S.; Wang, B.; Gao, W. The Enzyme Activity of Dual-Domain β-Propeller Alkaline Phytase as a Potential Factor in Improving Soil Phosphorus Fertility and Triticum aestivum Growth. Agronomy 2024, 14, 614. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030614
Zhang Y, Li J, Lu F, Wang S, Ren Y, Guo S, Wang B, Gao W. The Enzyme Activity of Dual-Domain β-Propeller Alkaline Phytase as a Potential Factor in Improving Soil Phosphorus Fertility and Triticum aestivum Growth. Agronomy. 2024; 14(3):614. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030614
Chicago/Turabian StyleZhang, Yitong, Jiangtao Li, Fang Lu, Siqi Wang, Yangjie Ren, Shiyang Guo, Ben Wang, and Wei Gao. 2024. "The Enzyme Activity of Dual-Domain β-Propeller Alkaline Phytase as a Potential Factor in Improving Soil Phosphorus Fertility and Triticum aestivum Growth" Agronomy 14, no. 3: 614. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030614