
Cadmium Exposure Impairs Development, Detoxification Mechanisms and Gene Expression of Glyphodes pyloalis Walker (Lepidoptera: Pyralidae)

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Cd Exposure
2.2. Effects of Cd Exposure on the Growth and Development of G. pyloalis
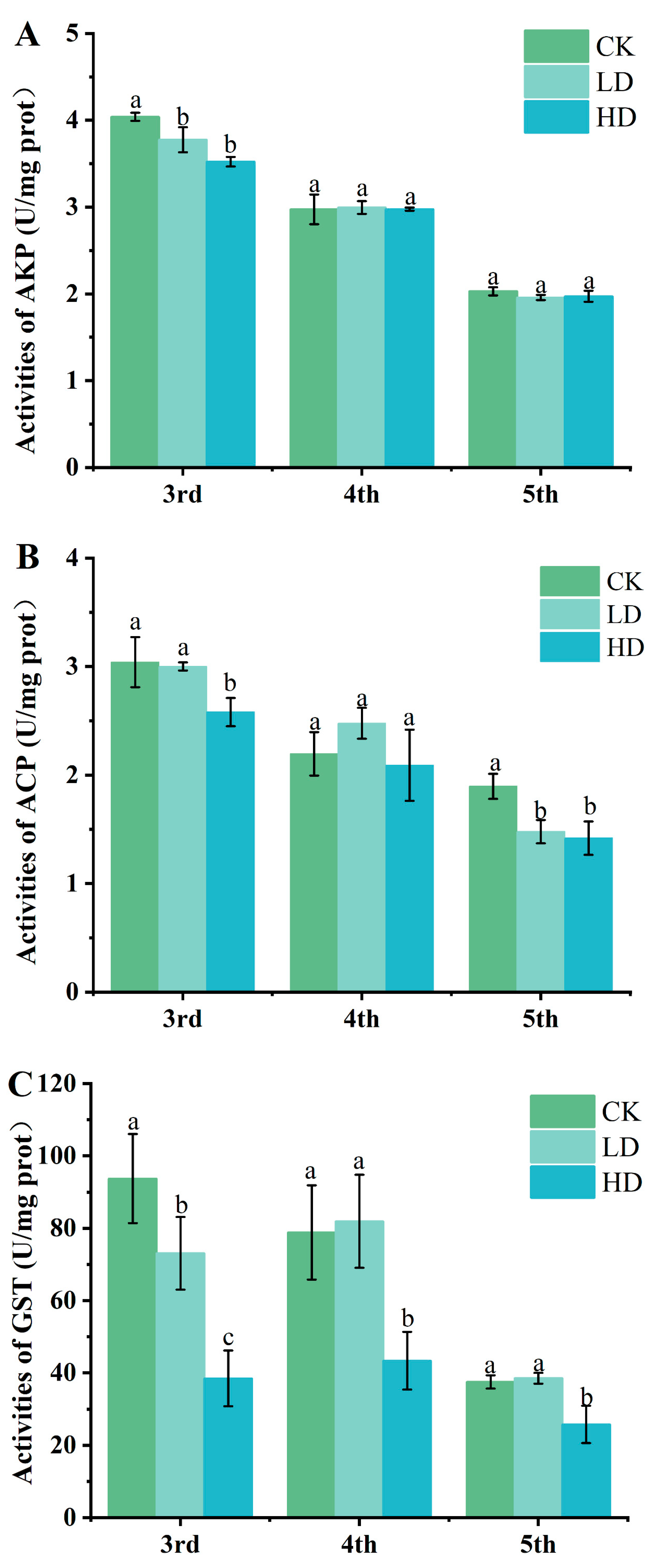
2.3. Assay of Detoxification Enzymes
| ACP and AKP (U/mg protein) = (measurement group absorbance value/standard group absorbance value) × the amount of phenol in standard group/(protein concentration of sample to be teste × sampling amount of homogenate supernatant) |
| GST activity (U/mg protein) = [(control group absorbance value − measurement group absorbance value)/(standard group absorbance value − blank group absorbance value)] × concentration of standard preparation × dilution multiple/reaction time/(sampling amount of homogenate supernatant × protein concentration of sample to be tested) |
2.4. RNA Extraction, Library Construction and Illumina Sequencing
2.5. De Novo Assembly and Annotation
2.6. Analysis of DEGs
2.7. RT-qPCR Analysis
2.8. RNAi and Bioassay
2.9. Statistical Analysis
3. Result
3.1. Effect of Cd on the Growth and Development of G. pyloalis
3.2. Effects of Cd Exposure on Activities of Detoxification Enzymes
3.3. Sequencing of the G. pyloalis Transcriptome and De Novo Assembly
3.4. Functional Annotation and Classification of Genes
3.5. SSR Analysis
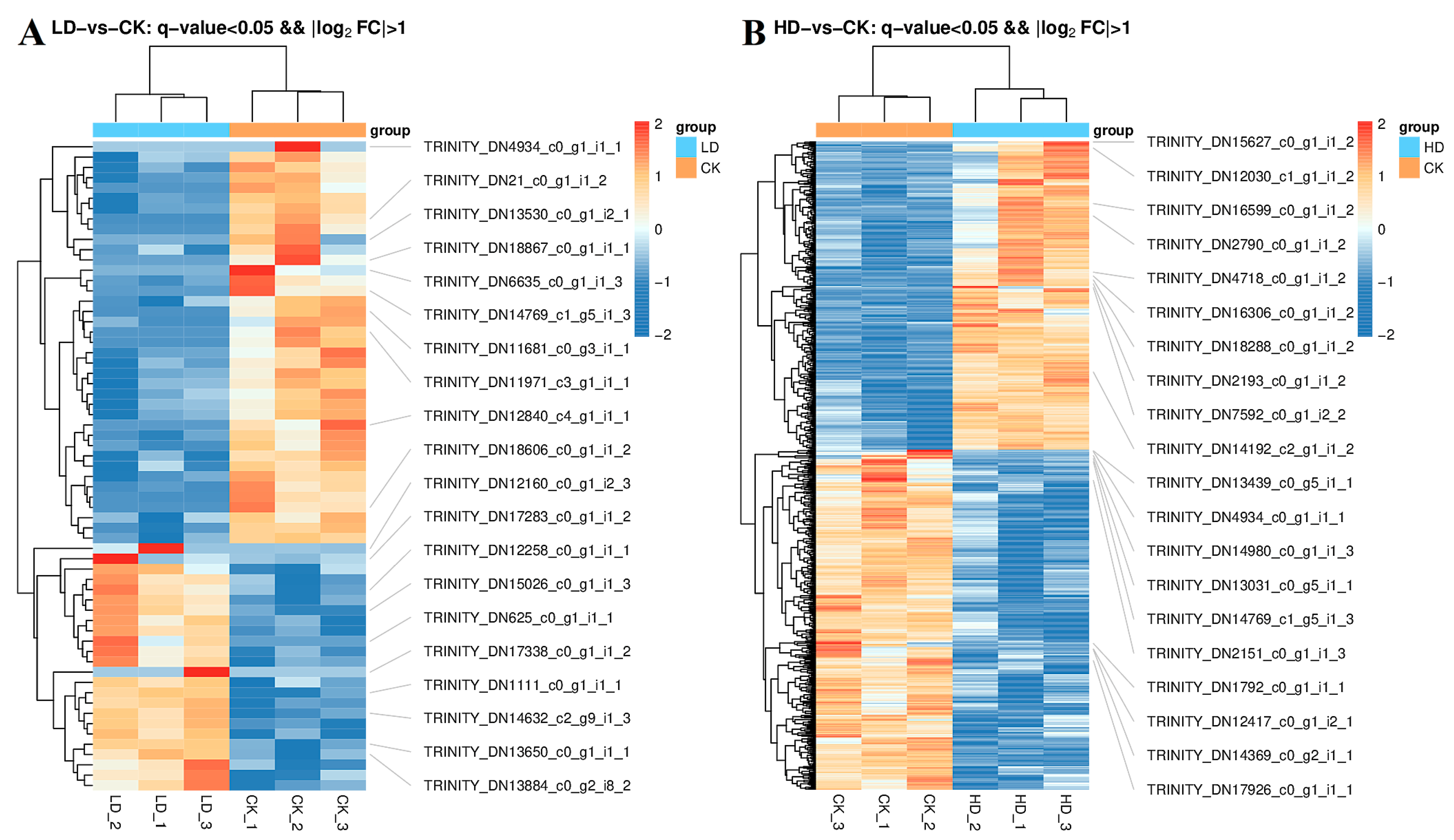
3.6. Statistical Analysis of DEG Number in Different Treatments
3.7. GO Analysis of DEGs
3.8. KEGG Analysis of DEGs
3.9. Validation by RT-qPCR
3.10. RNAi and Survivial Rate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, Y.G.; He, X.L.S.; Huang, J.H.; Luo, R.; Ge, H.Z.; Wołowicz, A.; Wawrzkiewicz, M.; Gładysz-Płaska, A.; Li, B.; Yu, Q.X.; et al. Impacts of Heavy Metals and Medicinal Crops on Ecological Systems, Environmental Pollution, Cultivation, and Production Processes in China. Ecotoxicol. Environ. Saf. 2021, 219, 112336. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Xu, X.R.; Sun, Y.X.; Liu, J.L.; Li, H. Bin Heavy Metal Pollution in Coastal Areas of South China: A Review. Mar. Pollut. Bull. 2013, 76, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Zhao, C.; Li, Q.; Feng, Y.; Zhang, X.; Lu, Y.; Ying, R.; Yin, A.; Ji, W. Heavy Metals Can Affect Plant Morphology and Limit Plant Growth and Photosynthesis Processes. Agronomy 2023, 13, 2601. [Google Scholar] [CrossRef]
- Shephard, A.M.; Mitchell, T.S.; Henry, S.B.; Oberhauser, K.S.; Kobiela, M.E.; Snell-Rood, E.C. Assessing Zinc Tolerance in Two Butterfly Species: Consequences for Conservation in Polluted Environments. Insect Conserv. Divers. 2020, 13, 201–210. [Google Scholar] [CrossRef]
- Nikolić, T.V.; Kojić, D.; Orčić, S.; Batinić, D.; Vukašinović, E.; Blagojević, D.P.; Purać, J. The Impact of Sublethal Concentrations of Cu, Pb and Cd on Honey Bee Redox Status, Superoxide Dismutase and Catalase in Laboratory Conditions. Chemosphere 2016, 164, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.H.; Zhao, H.Y.; Du, C.; Deng, M.M.; Du, E.X.; Hu, Z.Q.; Hu, X.S. Life Table Evaluation of Survival and Reproduction of the Aphid, Sitobion avenae, Exposed to Cadmium. J. Insect Sci. 2012, 12, 44. [Google Scholar] [CrossRef] [PubMed]
- Di, N.; Hladun, K.R.; Zhang, K.; Liu, T.X.; Trumble, J.T. Laboratory Bioassays on the Impact of Cadmium, Copper and Lead on the Development and Survival of Honeybee (Apis mellifera L.) Larvae and Foragers. Chemosphere 2016, 152, 108839. [Google Scholar] [CrossRef]
- Vlahović, M.; Matić, D.; Mutić, J.; Trifković, J.; Đurđić, S.; Perić Mataruga, V. Influence of Dietary Cadmium Exposure on Fitness Traits and Its Accumulation (with an Overview on Trace Elements) in Lymantria Dispar Larvae. Comp. Biochem. Physiol. Part—C Toxicol. Pharmacol. 2017, 200, 27–33. [Google Scholar] [CrossRef]
- Vlahović, M.; Ilijin, L.; Mrdaković, M.; Todorović, D.; Matić, D.; Lazarević, J.; Mataruga, V.P. Glutathione S-Transferase in the Midgut Tissue of Gypsy Moth (Lymantria dispar) Caterpillars Exposed to Dietary Cadmium. Environ. Toxicol. Pharmacol. 2016, 44, 13–17. [Google Scholar] [CrossRef]
- Mirčić, D.; Blagojević, D.; Perić-Mataruga, V.; Ilijin, L.; Mrdaković, M.; Vlahović, M.; Lazarević, J. Cadmium Effects on the Fitness-Related Traits and Antioxidative Defense of Lymantria dispar L. Larvae. Environ. Sci. Pollut. Res. 2013, 20, 209–218. [Google Scholar] [CrossRef]
- Zhao, J.; Sun, Y.; Xiao, L.; Tan, Y.; Bai, L. Molecular Characterization and Expression of Vitellogenin Gene from Spodoptera exigua Exposed to Cadmium Stress. Gene 2016, 593, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Borowska, J.; Pyza, E. Effects of Heavy Metals on Insect Immunocompetent Cells. J. Insect Physiol. 2011, 57, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Li, W.; Cheng, Y.; Ni, H.; Chen, X.; Li, Y.; Tang, B.; Sun, X.; Li, Y.; Liu, T.; et al. Copper Exposure Enhances Spodoptera Litura Larval Tolerance to β-Cypermethrin. Pestic. Biochem. Physiol. 2019, 160, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Poupardin, R.; Reynaud, S.; Strode, C.; Ranson, H.; Vontas, J.; David, J.P. Cross-Induction of Detoxification Genes by Environmental Xenobiotics and Insecticides in the Mosquito Aedes Aegypti: Impact on Larval Tolerance to Chemical Insecticides. Insect Biochem. Mol. Biol. 2008, 38, 540–551. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, M.; Aras, P.; Jumarie, C.; Boily, M. Low Dietary Levels of Al, Pb and Cd May Affect the Non-Enzymatic Antioxidant Capacity in Caged Honey Bees (Apis mellifera). Chemosphere 2016, 144, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Stone, D.; Jepson, P.; Laskowski, R. Trends in Detoxification Enzymes and Heavy Metal Accumulation in Ground beetles (Coleoptera: Carabidae) Inhabiting a Gradient of Pollution. Comp. Biochem. Physiol.—C Toxicol. Pharmacol. 2002, 132, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Ilijin, L.; Mrdaković, M.; Todorović, D.; Vlahović, M.; Gavrilović, A.; Mrkonja, A.; Perić-Mataruga, V. Life History Traits and the Activity of Antioxidative Enzymes in Lymantria dispar L. (Lepidoptera, Lymantriidae) Larvae Exposed to Benzo[a]Pyrene. Environ. Toxicol. Chem. 2015, 34, 2618–2624. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.K.; Velmurugan, K.; Plöttner, S.; Siegrist, K.J.; Romo, D.; Welge, P.; Brüning, T.; Xiong, K.N.; Käfferlein, H.U. Environmentally Prevalent Polycyclic Aromatic Hydrocarbons Can Elicit Co-Carcinogenic Properties in an in Vitro Murine Lung Epithelial Cell Model. Arch. Toxicol. 2018, 92, 1311–1322. [Google Scholar] [CrossRef]
- Xu, Z.B.; Zou, X.P.; Zhang, N.; Feng, Q.L.; Zheng, S.C. Detoxification of Insecticides, Allechemicals and Heavy Metals by Glutathione S-Transferase SlGSTE1 in the Gut of Spodoptera litura. Insect Sci. 2015, 22, 503–511. [Google Scholar] [CrossRef]
- Calvo-Marzal, P.; Rosatto, S.S.; Granjeiro, P.A.; Aoyama, H.; Kubota, L.T. Electroanalytical Determination of Acid Phosphatase Activity by Monitoring P-Nitrophenol. Anal. Chim. Acta 2001, 441, 207–214. [Google Scholar] [CrossRef]
- Vlahović, M.; Lazarević, J.; Perić-Mataruga, V.; Ilijin, L.; Mrdaković, M. Plastic Responses of Larval Mass and Alkaline Phosphatase to Cadmium in the Gypsy Moth Larvae. Ecotoxicol. Environ. Saf. 2009, 72, 1148–1155. [Google Scholar] [CrossRef] [PubMed]
- Vlahović, M.; Ilijin, L.; Matić, D.; Lazarević, J.; Nenadović, V.; Perić-Mataruga, V.; Mrdaković, M. Cadmium Effects on the Ratio of Activities of Lysosomal and Total Acid Phosphatases (ACPLys/ACPTot) in Lymantria dispar Larvae. Arch. Biol. Sci. 2013, 65, 345–352. [Google Scholar] [CrossRef]
- Stubberud, H.E.; Hønsi, T.G.; Stenersen, J. Purification and Partial Characterisation of Tentatively Classified Acid Phosphatase from the Earthworm Eisenia Veneta. Comp. Biochem. Physiol.—B Biochem. Mol. Biol. 2000, 126, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, Z.; Gao, Z.; Wen, Y.; Wang, W.; Liu, W.; Wang, X.; Zhu, F. Identification of Three Metallothioneins in the Black Soldier Fly and Their Functions in Cd Accumulation and Detoxification. Environ. Pollut. 2021, 286, 117146. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, R.; Sendi, J.J. Biology and Demography of Glyphodes Pyloalis Walker (Lepidoptera: Pyralidae) on Mulberry. J. Asia. Pac. Entomol. 2010, 13, 273–276. [Google Scholar] [CrossRef]
- Shao, Z.M.; Li, Y.J.; Zhang, X.R.; Chu, J.; Ma, J.H.; Liu, Z.X.; Wang, J.; Sheng, S.; Wu, F.A. Identification and Functional Study of Chitin Metabolism and Detoxification-related Genes in Glyphodes Pyloalis Walker (Lepidoptera: Pyralidae) Based on Transcriptome Analysis. Int. J. Mol. Sci. 2020, 21, 1904. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Zhu, Q.; Xu, Y.; Li, W.; Ding, Y.; Han, L.; Rensing, C.; Wang, R. Safe Utilization of Polluted Soil by Arsenic, Cadmium and Lead through an Integrated Sericultural Measure. Sci. Total Environ. 2019, 659, 1234–1241. [Google Scholar] [CrossRef]
- Jiang, D.; Tan, M.; Zheng, L.; Wu, H.; Li, Y.; Yan, S. Cd Exposure-Triggered Metabolic Disruption Increases the Susceptibility of Lymantria dispar (Lepidoptera: Erebidae) Larvae to Mamestra Brassicae Nuclear Polyhedrosis Virus: A Multi-Omics Study. Ecotoxicol. Environ. Saf. 2022, 232, 113280. [Google Scholar] [CrossRef]
- Jiang, D.; Zhou, Y.; Tan, M.; Zhang, J.; Guo, Q.; Yan, S. Cd Exposure-Induced Growth Retardation Involves in Energy Metabolism Disorder of Midgut Tissues in the Gypsy Moth Larvae. Environ. Pollut. 2020, 266, 115173. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Trinity: Reconstructing a Full-Length Transcriptome without a Genome from RNA-Seq Data HHS Public Access. Nat. Biotechnol. Nat. Biotechnol 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A Universal Tool for Annotation, Visualization and Analysis in Functional Genomics Research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Pertea, G.; Huang, X.; Liang, F.; Antonescu, V.; Sultana, R.; Karamycheva, S.; Lee, Y.; White, J.; Cheung, F.; Parvizi, B.; et al. TIGR Gene Indices Clustering Tools (TGICL): A Software System for Fast Clustering of Large EST Datasets. Bioinformatics 2003, 19, 651–652. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript Assembly and Quantification by RNA-Seq Reveals Unannotated Transcripts and Isoform Switching during Cell Differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nature Methods. Nat. Publ. Gr. 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative C(T) Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Dong, B.; Liu, X.Y.; Li, B.; Li, M.Y.; Li, S.G.; Liu, S. A Heat Shock Protein Protects against Oxidative Stress Induced by Lambda-Cyhalothrin in the Green Peach Aphid Myzus Persicae. Pestic. Biochem. Physiol. 2022, 181, 104995. [Google Scholar] [CrossRef]
- Dai, Y.; Zhang, Y.; Sun, W.; Chen, Y.; Wang, X.; Xin, T.; Wan, B.; Xia, B.; Zhong, L.; Zou, Z. The Metabolism and Detoxification Effects of Lead Exposure on Aleurolyphus ovatus (Acari: Acaridae) via Transcriptome Analysis. Chemosphere 2023, 333, 138886. [Google Scholar] [CrossRef]
- Su, H.; Wu, J.; Zhang, Z.; Ye, Z.; Chen, Y.; Yang, Y. Effects of Cadmium Stress at Different Concentrations on the Reproductive Behaviors of Beet Armyworm Spodoptera exigua (Hübner). Ecotoxicology 2021, 30, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jiang, D.; Dong, X.; Meng, Z.; Yan, S. Accumulation of Cd and Pb in Various Body Parts, Organs and Tissues of Lymantria dispar Asiatica (Lepidoptera: Erebidae). J. Asia. Pac. Entomol. 2020, 23, 963–969. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, J.; Shu, Y. Lead Stress Affects the Reproduction of Spodoptera litura but Not by Regulating the Vitellogenin Gene Promoter. Ecotoxicol. Environ. Saf. 2021, 208, 111581. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Yan, S. Effects of Cd, Zn or Pb Stress in Populus Alba Berolinensis on the Development and Reproduction of Lymantria dispar. Ecotoxicology 2017, 26, 1305–1313. [Google Scholar] [CrossRef] [PubMed]
- Bednarska, A.J.; Stachowicz, I. Costs of Living in Metal Polluted Areas: Respiration Rate of the Ground Beetle Pterostichus oblongopunctatus from Two Gradients of Metal Pollution. Ecotoxicology 2013, 22, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Moolman, L.; Van Vuren, J.H.J.; Wepener, V. Comparative Studies on the Uptake and Effects of Cadmium and Zinc on the Cellular Energy Allocation of Two Freshwater Gastropods. Ecotoxicol. Environ. Saf. 2007, 68, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Stolpe, C.; Müller, C. Effects of Single and Combined Heavy Metals and Their Chelators on Aphid Performance and Preferences. Environ. Toxicol. Chem. 2016, 35, 3023–3030. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Kong, J.; Seng, Y. Effects of the Heavy Metal Cu2+ on Growth, Development, and Population Dynamics of Spodoptera litura (Lepidoptera: Noctuidae). J. Econ. Entomol. 2012, 105, 288–294. [Google Scholar] [CrossRef]
- Tylko, G.; Banach, Z.; Borowska, J.; Niklińska, M.; Pyza, E. Elemental Changes in the Brain, Muscle, and Gut Cells of the Housefly, Musca Domestica, Exposed to Heavy Metals. Microsc. Res. Tech. 2005, 66, 239–247. [Google Scholar] [CrossRef]
- Sosa, V.; Moliné, T.; Somoza, R.; Paciucci, R.; Kondoh, H.; LLeonart, M.E. Oxidative Stress and Cancer: An Overview. Ageing Res. Rev. 2013, 12, 376–390. [Google Scholar] [CrossRef]
- Tiwari, S.; Pelz-Stelinski, K.; Stelinski, L.L. Effect of Candidatus Liberibacter Asiaticus Infection on Susceptibility of Asian Citrus Psyllid, Diaphorina citri, to Selected Insecticides. Pest Manag. Sci. 2011, 67, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Yi, Y. Effects of Dietary Heavy Metals on the Immune and Antioxidant Systems of Galleria mellonella Larvae. Comp. Biochem. Physiol. Part—C Toxicol. Pharmacol. 2015, 167, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Bream, A.S. Laboratory Evaluation of Heavy Metals Stress on Certain Biochemical Parameters of the Aquatic Insect, Sphaerodema Urinator Duf. (Hemiptera: Belostomatidae). Commun. Agric. Appl. Biol. Sci. 2003, 68, 291–297. [Google Scholar]
- Mazorra, M.T.; Rubio, J.A.; Blasco, J. Acid and Alkaline Phosphatase Activities in the Clam Scrobicularia Plana: Kinetic Characteristics and Effects of Heavy Metals. Comp. Biochem. Physiol.—B Biochem. Mol. Biol. 2002, 131, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Renella, G.; Ortigoza, A.L.R.; Landi, L.; Nannipieri, P. Additive Effects of Copper and Zinc on Cadmium Toxicity on Phosphatase Activities and ATP Content of Soil as Estimated by the Ecological Dose (ED50). Soil Biol. Biochem. 2003, 35, 1203–1210. [Google Scholar] [CrossRef]
- Kumar, S.; Asif, M.H.; Chakrabarty, D.; Tripathi, R.D.; Dubey, R.S.; Trivedi, P.K. Differential Expression of Rice Lambda Class GST Gene Family Members During Plant Growth, Development, and in Response to Stress Conditions. Plant Mol. Biol. Rep. 2013, 31, 569–580. [Google Scholar] [CrossRef]
- Wang, Y.; Fang, J.; Leonard, S.S.; Rao, K.M.K. Cadmium Inhibits the Electron Transfer Chain and Induces Reactive Oxygen Species. Free Radic. Biol. Med. 2004, 36, 1434–1443. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of Its Protective Roles, Measurement, and Biosynthesis. Mol. Aspects Med. 2009, 30, 1–12. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Song, D.N.; Wu, H.H.; Yang, H.M.; Zhang, J.Z.; Li, L.J.; Ma, E.B.; Guo, Y.P. Effect of Dietary Cadmium on the Activity of Glutathione S-Transferase and Carboxylesterase in Different Developmental Stages of the Oxya chinensis (Orthoptera: Acridoidea). Environ. Entomol. 2014, 43, 171–177. [Google Scholar] [CrossRef]
- Bernabò, P.; Gaglio, M.; Bellamoli, F.; Viero, G.; Lencioni, V. DNA Damage and Translational Response during Detoxification from Copper Exposure in a Wild Population of Chironomus Riparius. Chemosphere 2017, 173, 235–244. [Google Scholar] [CrossRef]
- Lee, S.M.; Lee, S.B.; Park, C.H.; Choi, J. Expression of Heat Shock Protein and Hemoglobin Genes in Chironomus Tentans (Diptera, Chironomidae) Larvae Exposed to Various Environmental Pollutants: A Potential Biomarker of Freshwater Monitoring. Chemosphere 2006, 65, 1074–1081. [Google Scholar] [CrossRef] [PubMed]
- El-Gendy, A.H.; Augustyniak, M.; Toto, N.A.; Al Farraj, S.; El-Samad, L.M. Oxidative Stress Parameters, DNA Damage and Expression of HSP70 and MT in Midgut of Trachyderma hispida (Forskål, 1775) (Coleoptera: Tenebrionidae) from a Textile Industry Area. Environ. Pollut. 2020, 267, 115661. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Yang, H.L.; Peng, Y.D.; Wang, J.; Zeng, Z.; Li, N.; Tang, Y.E.; Wang, Z.; Song, Q.S. Cadmium Exposure Alters Expression of Protective Enzymes and Protein Processing Genes in Venom Glands of the Wolf Spider Pardosa pseudoannulata. Environ. Pollut. 2021, 268, 115847. [Google Scholar] [CrossRef] [PubMed]
- del Rio, L.A. Reactive Oxygen Species, Antioxidant Systems and Nitric Oxide in Peroxisomes. J. Exp. Bot. 2002, 53, 1255–1272. [Google Scholar] [CrossRef]
- Stern, D. Body-Size Control: How an Insect Knows It Has Grown Enough. Curr. Biol. 2003, 13, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Barg, R.; Yin, M.; Gueta-Dahan, Y.; Leikin-Frenkel, A.; Salts, Y.; Shabtai, S.; Ben-Hayyim, G. Modulated Fatty Acid Desaturation via Overexpression of Two Distinct ω-3 Desaturases Differentially Alters Tolerance to Various Abiotic Stresses in Transgenic Tobacco Cells and Plants. Plant J. 2005, 44, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Harrop, T.W.R.; Pearce, S.L.; Daborn, P.J.; Batterham, P. Whole-Genome Expression Analysis in the Third Instar Larval Midgut of Drosophila melanogaster. G3 Genes Genomes Genet. 2014, 4, 2197–2205. [Google Scholar] [CrossRef]
- Poupardin, R.; Riaz, M.A.; Vontas, J.; David, J.P.; Reynaud, S. Transcription Profiling of Eleven Cytochrome P450s Potentially Involved in Xenobiotic Metabolism in the Mosquito Aedes aegypti. Insect Mol. Biol. 2010, 19, 185–193. [Google Scholar] [CrossRef]
- Jin, Y.; Cong, B.; Wang, L.; Gao, Y.; Zhang, H.; Dong, H.; Lin, Z. Differential Gene Expression Analysis of the Epacromius coerulipes (Orthoptera: Acrididae) Transcriptome. J. Insect Sci. 2016, 16, 42. [Google Scholar] [CrossRef]
- Pan, X.; Ding, J.-H.; Zhao, S.-Q.; Shi, H.-C.; Miao, W.-L.; Wu, F.-A.; Sheng, S.; Zhou, W.-H. Identification and Functional Study of Detoxification-Related Genes in Response to Tolfenpyrad Stress in Glyphodes pyloalis Walker (Lepidoptera: Pyralidae). Pestic. Biochem. Physiol. 2023, 194, 105503. [Google Scholar] [CrossRef]
- Meng, X.; Zhang, N.; Yang, X.; Miao, L.; Jiang, H.; Ji, C.; Xu, B.; Qian, K.; Wang, J. Sublethal Effects of Chlorantraniliprole on Molting Hormone Levels and MRNA Expressions of Three Halloween Genes in the Rice Stem Borer, Chilo Suppressalis. Chemosphere 2020, 238, 124676. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, G.; Zhou, Q. Molecular Characterization of Cytochrome P450 CYP6B47 CDNAs and 5′-Flanking Sequence from Spodoptera litura (Lepidoptera: Noctuidae): Its Response to Lead Stress. J. Insect Physiol. 2012, 58, 726–736. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, S.; Miao, W.; Sheng, S.; Pan, X.; Li, P.; Zhou, W.; Wu, F. Cadmium Exposure Impairs Development, Detoxification Mechanisms and Gene Expression of Glyphodes pyloalis Walker (Lepidoptera: Pyralidae). Agronomy 2024, 14, 626. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030626
Zhao S, Miao W, Sheng S, Pan X, Li P, Zhou W, Wu F. Cadmium Exposure Impairs Development, Detoxification Mechanisms and Gene Expression of Glyphodes pyloalis Walker (Lepidoptera: Pyralidae). Agronomy. 2024; 14(3):626. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030626
Chicago/Turabian StyleZhao, Shuaiqi, Wanglong Miao, Sheng Sheng, Xin Pan, Ping Li, Weihong Zhou, and Fuan Wu. 2024. "Cadmium Exposure Impairs Development, Detoxification Mechanisms and Gene Expression of Glyphodes pyloalis Walker (Lepidoptera: Pyralidae)" Agronomy 14, no. 3: 626. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy14030626