Characterization for Drought Tolerance and Physiological Efficiency in Novel Cytoplasmic Male Sterile Sources of Sunflower (Helianthus annuus L.)


Abstract
:1. Introduction
2. Results
2.1. Variation in Parents and Their Hybrids
2.2. Combining Ability Estimates
2.2.1. Number of Leaves per Plant
2.2.2. Leaf Area (m2)
2.2.3. Specific Leaf Weight (g)
2.2.4. Leaf Area Index
2.2.5. Leaf Water Potential (MPa)
2.2.6. Relative Leaf Water Content (%)
2.2.7. Photosynthetic Efficiency (Chlorophyll Meter Reading)
2.2.8. Proline Content (mg/g of Dry Weight)
2.2.9. Seed Yield (g/Plant)
2.2.10. Biological Yield (g/Plant)
2.2.11. Harvest Index (%)
2.2.12. Oil Content (%)
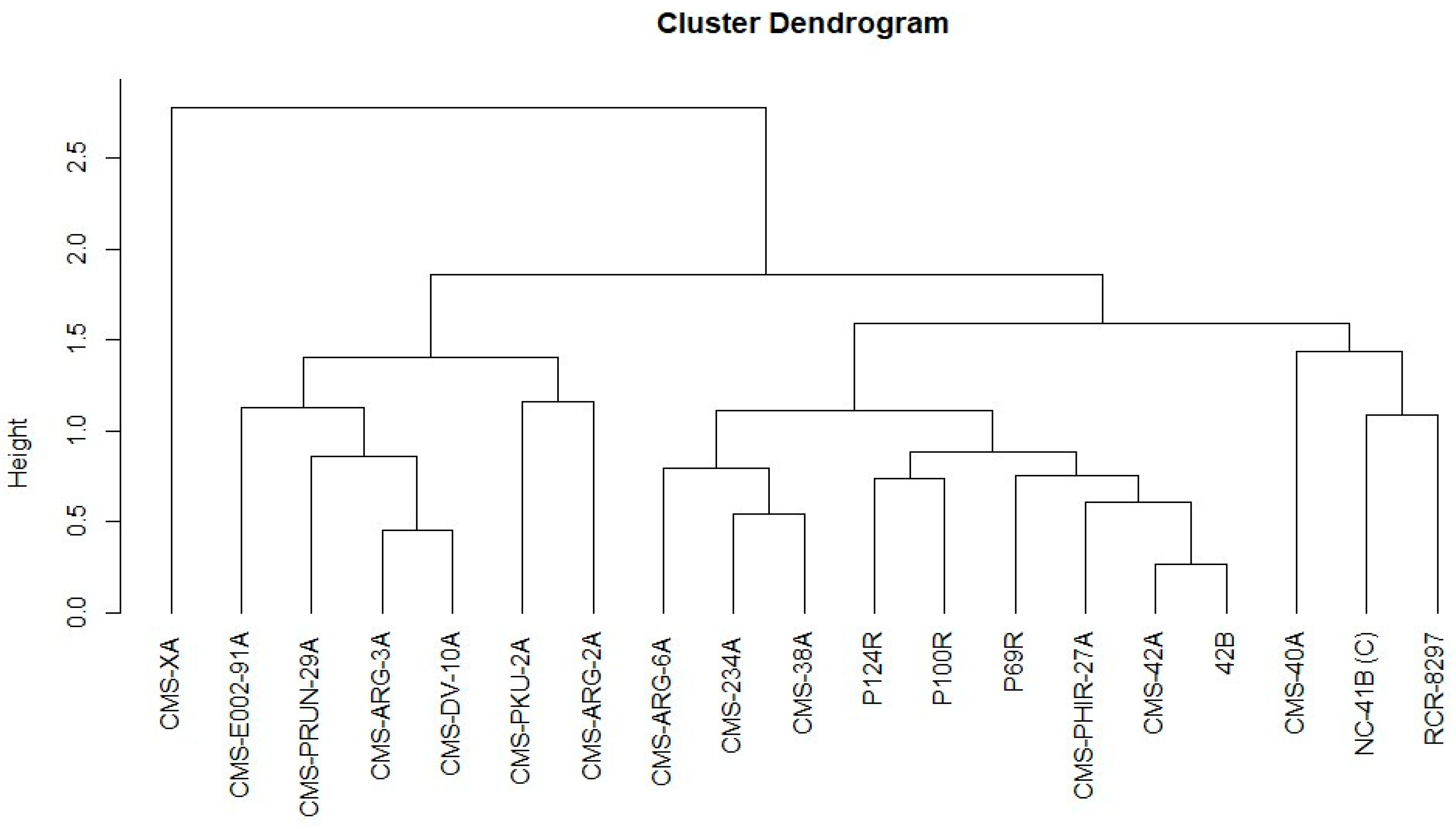
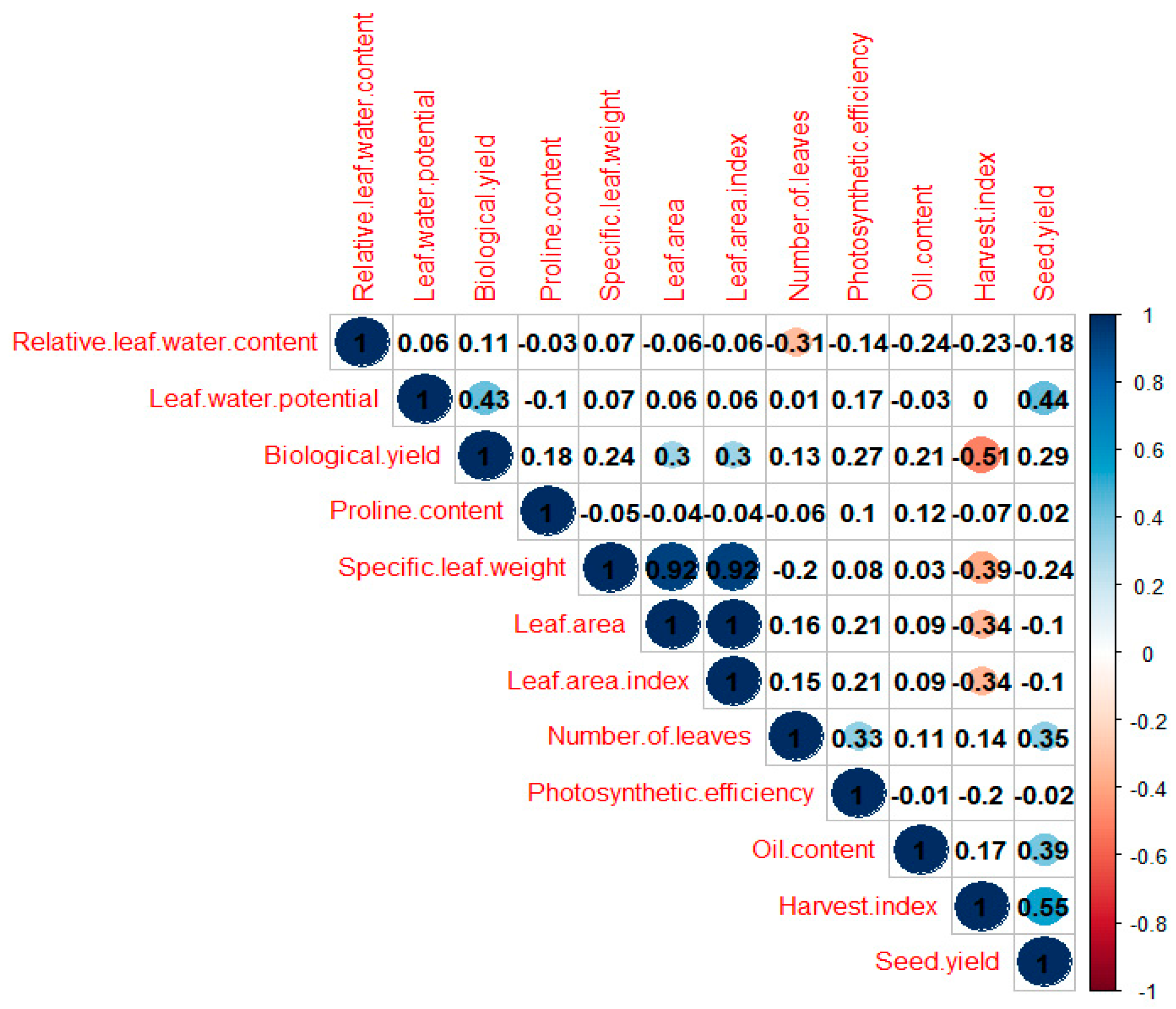
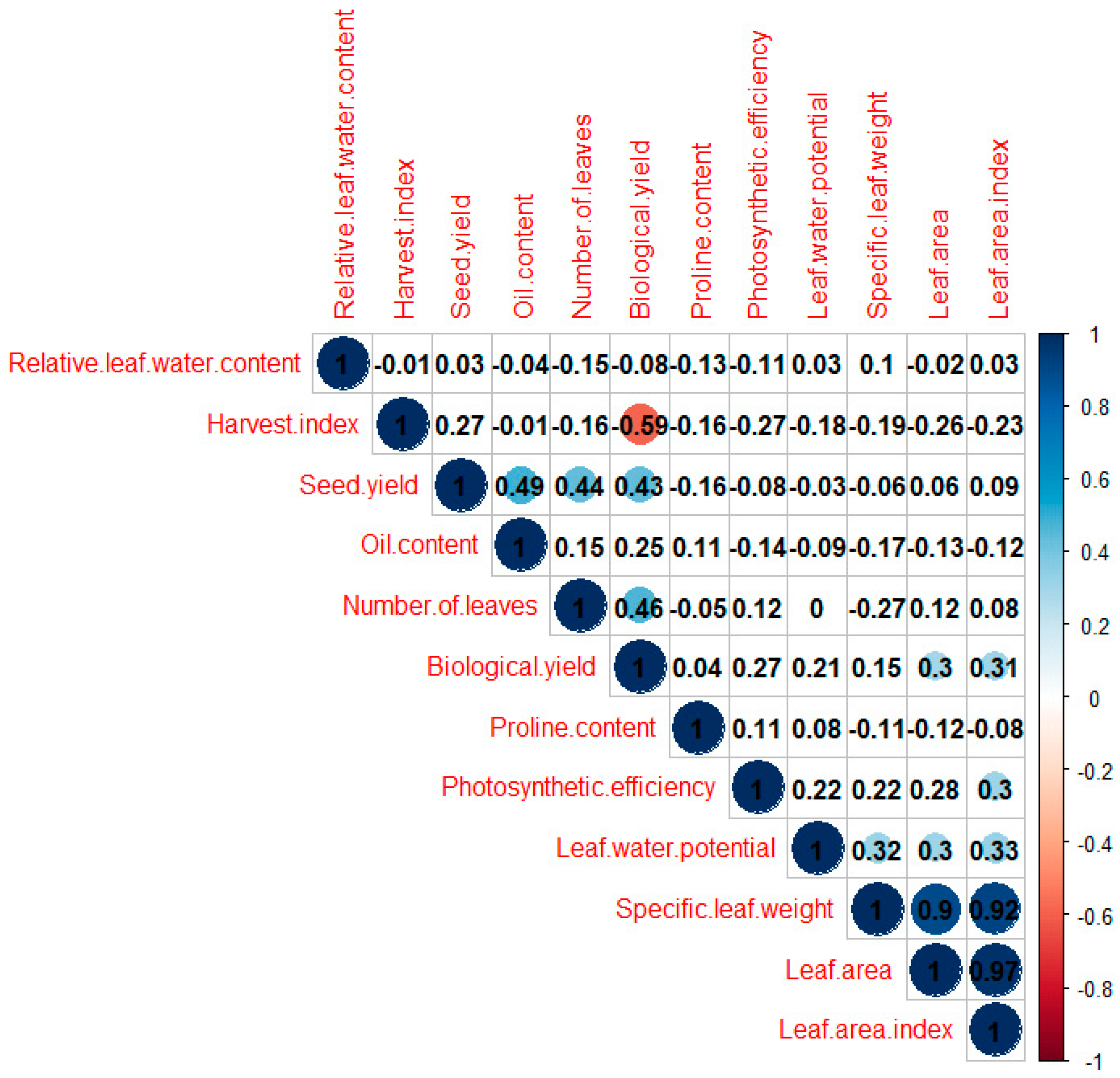
2.3. Correlations
3. Discussion
4. Materials and Methods
4.1. Experimental Layout and Material
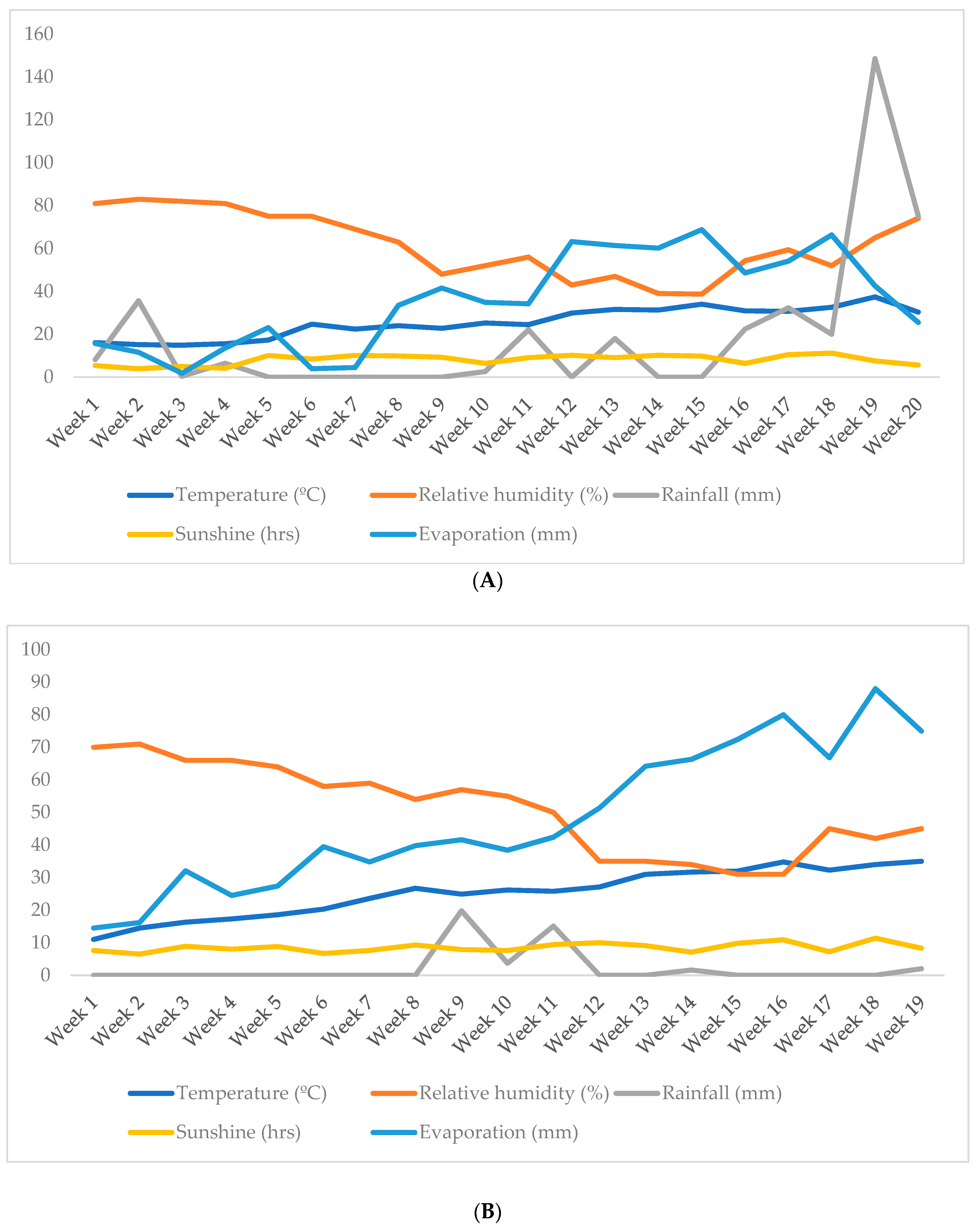
4.2. Weather Parameters and Soil Properties
4.3. Characterization of Plants and Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| A | Cytoplasmic male sterile line |
| B | Maintainer line |
| CMS | Cytoplasmic male sterile |
| GA3 | Gibberellic acid |
| GCA | General combining ability |
| HI | Harvest index |
| LAI | Leaf area index |
| R | Restorer line |
| RCBD | Randomized complete block design |
| RLWC | Relative leaf water content |
| SCA | Specific combining ability |
References
- Pal, D. Sunflower (Helianthus annuus L.) Seeds in Health and Nutrition—Chapter 130. In Nuts and Seeds in Health and Disease Prevention; Preedy, V.R., Watson, R.R., Patel, V.B., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 1097–1105. ISBN 978-0-12-375688-6. [Google Scholar]
- González-Alonso, A.; Ramírez-Tortosa, C.; Varela-López, A.; Roche, E.; Arribas, M.; Ramírez-Tortosa, M.; Giampieri, F.; Ochoa, J.; Quiles, J.; González-Alonso, A.; et al. Sunflower Oil but Not Fish Oil Resembles Positive Effects of Virgin Olive Oil on Aged Pancreas after Life-Long Coenzyme Q Addition. Int. J. Mol. Sci. 2015, 16, 23425–23445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhi, S.D.; Heesacker, A.F.; Freeman, C.A.; Argyris, J.; Bradford, K.; Knapp, S.J. The self-incompatibility locus (S) and quantitative trait loci for self-pollination and seed dormancy in sunflower. Theoret. Appl. Genet. 2005, 111, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Lai, Z.; Nakazato, T.; Salmaso, M.; Burke, J.M.; Tang, S.; Knapp, S.J.; Rieseberg, L.H. Extensive Chromosomal Repatterning and the Evolution of Sterility Barriers in Hybrid Sunflower Species. Genetics 2005, 171, 291–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatun, M.; Hossain, T.M.; Miah, M.M.; Khandoker, S.; Rashid, M.A. Profitability of sunflower cultivation in some selected sites of Bangladesh. Bangladesh J. Agric. Res. 2016, 41, 599–623. [Google Scholar] [CrossRef]
- Sheoran, P.; Sheoran, O.P.; Sardana, V. Modeling Sunflower Productivity and Profitability in Relation to Adequate and Limited Sulphur Availability under Semiarid Irrigated Conditions. Int. J. Agron. 2013, 2013, 738263. Available online: https://www.hindawi.com/journals/ija/2013/738263/ (accessed on 20 August 2018). [CrossRef]
- Dimitrijevic, A.; Horn, R. Sunflower Hybrid Breeding: From Markers to Genomic Selection. Front. Plant Sci. 2018, 8, 2238. [Google Scholar] [CrossRef] [PubMed]
- Bohra, A.; Jha, U.C.; Adhimoolam, P.; Bisht, D.; Singh, N.P. Cytoplasmic male sterility (CMS) in hybrid breeding in field crops. Plant Cell Rep. 2016, 35, 967–993. [Google Scholar] [CrossRef] [PubMed]
- Jansky, S. Breeding, Genetics, and Cultivar Development—Chapter 2. In Advances in Potato Chemistry and Technology; Singh, J., Kaur, L., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 27–62. ISBN 978-0-12-374349-7. [Google Scholar]
- Griffiths, A.J.; Miller, J.H.; Suzuki, D.T.; Lewontin, R.C.; Gelbart, W.M. Cytoplasmic male sterility. In An Introduction to Genetic Analysis, 7th ed.; W.H. Freeman and Company: San Francisco, CA, USA, 2000. [Google Scholar]
- Leclercq, P. Une sterilite male cytoplasmique chez le tournesol. Ann. Amel. Plantes 1969, 19, 99–106. [Google Scholar]
- Leclercq, P.; Philippon, J.P. Identification de gènes de restauration de fertilité sur cytoplasmes stérilisants chez le tournesol. Agronomie 1984, 4, 573–576. [Google Scholar] [CrossRef]
- Horn, R.; Gupta, K.J.; Colombo, N. Mitochondrion role in molecular basis of cytoplasmic male sterility. Mitochondrion 2014, 19 Pt B, 198–205. [Google Scholar] [CrossRef]
- Echeverria, M.M.; Salaberry, M.T.; Rodriguez, R.H. Characterization for agronomic use of cytoplasmic male-sterility in sunflower (Helianthus annuus L.) introduced from H. resinosus Small. Plant Breed. 2003, 122, 357–361. [Google Scholar] [CrossRef]
- Owens, G.L.; Baute, G.J.; Hubner, S.; Rieseberg, L.H. Genomic sequence and copy number evolution during hybrid crop development in sunflowers. Evol. Appl. 2018. [Google Scholar] [CrossRef]
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought Stress in Plants: Causes, Consequences, and Tolerance. In Drought Stress Tolerance in Plants; Springer: Cham, Switzerland, 2016; Volume 1, pp. 1–16. ISBN 978-3-319-28897-0. [Google Scholar]
- Kaya, Y.; Pekcan, V.; Cicek, N. Effects of Drought on Morphological Traits of some Sunflower Lines. Ekin J. 2016, 2, 54–68. [Google Scholar]
- Prasad, P.V.V.; Staggenborg, S.; Ristic, Z.; Ahuja, L.; Reddy, V.; Anapalli, S.; Yu, Q. Impacts of Drought and/or Heat Stress on Physiological, Developmental, Growth, and Yield Processes of Crop Plants; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2008. [Google Scholar]
- Pekcan, V.; Evci, G.; Ibrahim Yilmaz, M.; Balkan Nalçaiyi, S.; Çulha Erdal, Ş.; Çiçek, N.; Ekmekci, Y.; Kaya, Y. Drought effects on yield traits of some sunflower inbred lines. Podgorica 2015, 61, 101. [Google Scholar] [CrossRef]
- Seiler, G.J.; Qi, L.L.; Marek, L. Use of Sunflower Crop Wild Relatives for Cultivated Sunflower Improvement. Crop Sci. 2017, 57, 1083–1101. [Google Scholar] [CrossRef]
- Vukich, M.; Schulman, A.H.; Giordani, T.; Natali, L.; Kalendar, R.; Cavallini, A. Genetic variability in sunflower (Helianthus annuus L.) and in the Helianthus genus as assessed by retrotransposon-based molecular markers. Theoret. Appl. Genet. 2009, 119, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Darvishzadeh, R.; Pirzad, A.; Bernousi, I.; Mandoulakani, B.A.; Azizi, H.; Akhondi, N.; Kiani, S.P.; Sarrafi, A. Genetic properties of drought tolerance indices in sunflower. Acta Agric. Scand. B Soil Plant 2011, 61, 593–601. [Google Scholar] [CrossRef]
- Rauf, S. Breeding sunflower (Helianthus annuus L.) for drought tolerance. Commun. Biometry Crop Sci. 2008, 3, 29–44. [Google Scholar]
- Cechin, I.; Rossi, S.C.; Oliveira, V.C.; Fumis, T.F. Photosynthetic responses and proline content of mature and young leaves of sunflower plants under water deficit. Photosynthetica 2006, 44, 143–146. [Google Scholar] [CrossRef]
- Levings, C.S. The Texas cytoplasm of maize: Cytoplasmic male sterility and disease susceptibility. Science 1990, 250, 942–947. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.J.; Koeppe, D.E. Southern corn leaf blight: Susceptible and resistant mitochondria. Science 1971, 173, 67–69. [Google Scholar] [CrossRef] [PubMed]
- Reddemann, A.; Horn, R.; Reddemann, A.; Horn, R. Recombination Events Involving the atp9 Gene Are Associated with Male Sterility of CMS PET2 in Sunflower. Int. J. Mol. Sci. 2018, 19, 806. [Google Scholar] [CrossRef] [PubMed]
- Jan, C.C. Cytoplasmic Male Sterility in Two Wild Helianthus annuus L. Accessions and Their Fertility Restoration. Crop Sci. 2000, 40, 1535–1538. [Google Scholar] [CrossRef]
- Tyagi, V.; Dhillon, S. Effect of Alien Cytoplasm on Combining Ability for Earliness and Seed Yield in Sunflower under Irrigation and Drought Stress. Helia 2017, 40, 71–83. [Google Scholar] [CrossRef]
- Horn, R. Molecular diversity of male sterility inducing and male-fertile cytoplasms in the genus Helianthus. Theor. Appl. Genet. 2002, 104, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Fujino, Y.; Murata, H.; Mayama, C.; Asaoka, R. A Review on Mating Designs. Nat. Sci. 2015, 13, 98–105. [Google Scholar]
- Clark, P.J.; Kempthorne, O. An Introduction to Genetic Statistics; John Wiley & Sons, Inc.: New York, NY, USA, 1958; Volume 39, p. 313. [Google Scholar]
- Numerical Ecology, Volume 24—2nd Edition. Available online: https://0-www-elsevier-com.brum.beds.ac.uk/books/numerical-ecology/legendre/978-0-444-89249-2 (accessed on 15 July 2018).
- Khan, A.; Sovero, V.; Gemenet, D. Genome-assisted Breeding for Drought Resistance. Curr. Genom. 2016, 17, 330–342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Mittal, N.; Leamy, L.J.; Barazani, O.; Song, B. Back into the wild—Apply untapped genetic diversity of wild relatives for crop improvement. Evol. Appl. 2016, 10, 5–24. [Google Scholar] [CrossRef] [PubMed]
- Ford-Lloyd, B.V.; Schmidt, M.; Armstrong, S.J.; Barazani, O.; Engels, J.; Hadas, R.; Hammer, K.; Kell, S.P.; Kang, D.; Khoshbakht, K.; et al. Crop Wild Relatives—Undervalued, Underused and under Threat? BioScience 2011, 61, 559–565. [Google Scholar] [CrossRef]
- Budak, H.; Kantar, M.; Yucebilgili, K.K. Drought Tolerance in Modern and Wild Wheat. Sci. World J. 2013, 2013, 548246. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Dixit, S.; Ram, T.; Yadaw, R.B.; Mishra, K.K.; Mandal, N.P. Breeding high-yielding drought-tolerant rice: Genetic variations and conventional and molecular approaches. J. Exp. Bot. 2014, 65, 6265–6278. [Google Scholar] [CrossRef] [PubMed]
- Škorić, D. Sunflower Breeding for Resistance to Abiotic and Biotic Stresses. In Abiotic and Biotic Stress in Plants—Recent Advances and Future Perspectives; InTech: Rijeka, Croatia, 2016. [Google Scholar] [Green Version]
- Seiler, G.J. Wild annual Helianthus anomalus and H. deserticola for improving oil content and quality in sunflower. Ind. Crops Prod. 2007, 25, 95–100. [Google Scholar] [CrossRef]
- Hussain, M.M.; Rauf, S.; Riaz, M.A.; Al-Khayri, J.M.; Monneveux, P. Determination of drought tolerance related traits in Helianthus argophyllus, Helianthus annuus, and their hybrids. Breed. Sci. 2017, 67, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Joseph, B.; Corwin, J.A.; Züst, T.; Li, B.; Iravani, M.; Schaepman-Strub, G.; Turnbull, L.A.; Kliebenstein, D.J. Hierarchical Nuclear and Cytoplasmic Genetic Architectures for Plant Growth and Defense within Arabidopsis. Plant Cell 2013, 25, 1929–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Lu, G.; Long, W.; Zou, X.; Li, F.; Nishio, T. Recent progress in drought and salt tolerance studies in Brassica crops. Breed. Sci. 2014, 64, 60–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanetomo, R.; Gebhardt, C. Cytoplasmic genome types of European potatoes and their effects on complex agronomic traits. BMC Plant Biol. 2015, 15, 162. [Google Scholar] [CrossRef] [PubMed]
- Beavis, W.D.; Pollak, E.; Frey, K.J. A theoretical model for quantitatively inherited traits influenced by nuclear-cytoplasmic interactions. Theoret. Appl. Genet. 1987, 74, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Xiang, H.; Tian, J.; Yin, J.; Pinkert, C.A.; Li, Q.; Zhao, X. Mitochondrial Haplotypes Influence Metabolic Traits in Porcine Transmitochondrial Cybrids. Sci. Rep. 2015, 5, 13118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushik, P.; Dhaliwal, M.S. Diallel Analysis for Morphological and Biochemical Traits in Tomato Cultivated under the Influence of Tomato Leaf Curl Virus. Agronomy 2018, 8, 153. [Google Scholar] [CrossRef]
- Putt, E.D. Heterosis, Combining Ability, and Predicted Synthetics from a Diallel Cross in Sunflowers (Helianthus annuus L.). Can. J. Plant Sci. 1966, 46, 59–67. [Google Scholar] [CrossRef]
- Giriraj, K.; Hiremath, S.R.; Seenappa, K. Combining ability of converted male sterile lines of sunflower (Helianthus annuus L.). Indian J. Genet. Plant Breed. 1987, 47, 315–317. [Google Scholar]
- Hladni, N.; Jocić, S.; Miklič, V.; Mijić, A.; Saftić-Panković, D.; Škorić, D. Effect of morphological and physiological traits on seed yield and oil content in sunflower. Helia 2014, 33, 101–116. [Google Scholar] [CrossRef]
- Dudhe, M.Y.; Moon, M.K.; Lande, S.S. Study of gene action for restorer lines in sunflower. Helia 2014, 34, 159–164. [Google Scholar] [CrossRef]
- Golabadi, M.; Golkar, P.; Shahsavari, M.R. Genetic analysis of agro-morphological traits in promising hybrids of sunflower (Helianthus annuus L.). Acta Agric. Slov. 2015, 105, 249–260. [Google Scholar] [CrossRef]
- Jocić, S.; Cvejić, S.; Ciric, M.; Hladni, N.; Miladinović, D.; Miklic, V. Estimation of combining abilities in sunflower (Helianthus annuus L.). In Proceedings of the 18th International Sunflower Conference, Mar del Plata, Argentina, 27 February–1 March 2012; pp. 657–662. [Google Scholar]
- Salem, A.H.; Ali, M.A. Combining ability for sunflower yield contributing characters and oil content over different water supply environments. J. Am. Sci. 2012, 8, 230–237. [Google Scholar]
- Kaya, Y.; Atakisi, I.K. Combining ability analysis of some yield characters of sunflower (Helianthus annuus L.). Helia 2004, 41, 75–84. [Google Scholar] [CrossRef]
- Andarkhor, S.A.; Mastibege, N.; Rameeh, V. Combining ability of agronomic traits in Sunflower (Helianthus annuus L.) using line X tester analysis. Int. J. Biol. 2011, 4, 89–95. [Google Scholar] [CrossRef]
- Mirarab, M.; Ahmadikhah, A. Study on genetics of some important phonological traits in rice using line × tester analysis. Ann. Biol. Res. 2010, 4, 119–125. [Google Scholar]
- Kang, S.A.; Khan, F.A.; Ahsan, M.Z.; Chatha, W.S.; Saeed, F. Estimation of combining ability for the development of hybrid genotypes in sunflower (Helianthus annuus L.). J. Biol. Agric. Healthc. 2013, 39, 68–74. [Google Scholar]
- Memon, S.; Baloch, M.J.; Baloch, G.M.; Jatoi, W.A. Combining ability through line × tester analysis for phenological, seed yield, and oil traits in sunflower (Helianthus annuus L.). Euphytica 2015, 204, 199–209. [Google Scholar] [CrossRef]
- Kadkol, G.P.; Anand, I.J.; Sharma, R.P. Combining ability and heterosis in sunflower. Indian J. Genet. Plant Breed. 1984, 44, 447–451. [Google Scholar]
- Andarkhor, S.A.; Rameeh, V.; Alitabar, R.A. Estimation of genetic parameters for yield components and seed yield in sunflower using line x tester analysis. Afr. J. Biotechnol. 2013, 12, 3978–3983. [Google Scholar]
- Karasu, A.; Mehmet, O.Z.; Sincik, M.; Goksoy, A.T.; Turan, Z.M. Combining ability and heterosis for yield and yield components in sunflower. Not. Bot. Horti Agrobot. Cluj Napoca 2010, 38, 259–264. [Google Scholar]
- Pace, B.A.; Alexander, H.M.; Emry, J.D.; Mercer, K.L. Seed fates in crop–wild hybrid sunflower: Crop allele and maternal effects. Evol. Appl. 2015, 8, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Seetharam, A.; Kumari, P.K. Induction of male sterility by gibberellic acid in sunflower. Indian J. Genet. Plant Breed. 1975, 35, 136–138. [Google Scholar]
- Baydar, H.; Gökmen, O.Y.; Friedt, W. Hybrid seed production in safflower (Carthamus tinctorius) following the induction of male sterility by gibberellic acid. Plant Breed. 2003, 122, 459–461. [Google Scholar] [CrossRef]
- Razzaq, H.; Tahir, N.; Hammad, M.; Ahmad Sadaqat, H.; Sadia, B. Screening of sunflower (Helianthus annus L.) accessions under drought stress conditions, an experimental assay. J. Soil Sci. Plant Nutr. 2017, 17, 662–671. [Google Scholar] [CrossRef]
- Bandyopadhyay, K.; Aggarwal, P.; Chakraborty, D.; Pradhan, S.; Narayan Garg, R.; Singh, R. Practical Manual on Measurement of Soil Physical Properties Practical; Indian Agricultural Research Institute: New Delhi, India, 2012; pp. 8–25. [Google Scholar]
- Maldaner, I.C.; Heldwein, A.B.; Loose, L.H.; Lucas, D.D.P.; Guse, F.I.; Bortoluzzi, M.P. Modelos de determinação não-destrutiva da área foliar em girassol. Cienc. Rural 2009, 39, 1356–1361. [Google Scholar] [CrossRef] [Green Version]
- Smart, R.E.; Bingham, G.E. Rapid Estimates of Relative Water Content. Plant Physiol. 1974, 53, 258–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, Q.; Huang, W.; Jarvis, P. Use of a SPAD-502 meter to measure leaf chlorophyll concentration in Arabidopsis thaliana. Photosynth. Res. 2011, 107, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Wnuk, A.; Górny, A.; Bocianowski, J.; Kozak, M. Visualizing harvest index in crops. Commun. Biometry Crop Sci. 2013, 8, 48–59. [Google Scholar]
- Robertson, J.A.; Morrison, W.H. Analysis of oil content of sunflower seed by wide-line NMR. J. Am. Oil Chem. Soc. 1979, 56, 961–964. [Google Scholar] [CrossRef]
- Esteves, F. R Script for UPGMA Analysis. Available online: https://www.researchgate.net/publication/303944638_R_script_for_Principal_Component_Analysis_PCA (accessed on 15 July 2018).
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Corrplot: Visualization of a Correlation Matrix. 2017. Available online: http://cran.r-project.org/package=corrplot (accessed on 2 August 2018).
- Peterson, B.G.; Carl, P.; Boudt, K.; Bennett, R.; Ulrich, J.; Zivot, E.; Cornilly, D.; Hung, E.; Lestel, M.; Balkissoon, K.; et al. Performance Analytics: Econometric Tools for Performance and Risk Analysis. 2018. Available online: https://github.com/braverock/PerformanceAnalytics (accessed on 2 August 2018).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptors | Parent (n = 19) | Parent Stress | Hybrid (n = 60) | Hybrid Stress | F-Ratio | Probability |
|---|---|---|---|---|---|---|
| Mean a | Mean a | Mean a | Mean a | |||
| (Range) | (Range) | (Range) | (Range) | |||
| Number of leaves | 22.31 b | 19.50 a | 24.65 b | 24.10 b | 9.61 | <0.001 |
| (20.55–24.05) | (17.75–21.25) | (23.66–25.64) | (23.11–25.08) | |||
| Leaf area (m2) | 0.62 b | 0.57 ab | 0.53 a | 0.45 ab | 3.98 | 0.0092 |
| (0.52–0.72) | (0.47–0.67) | (0.47–0.58) | (0.39–0.50) | |||
| Specific leaf weight (g) | 1.48 c | 1.38 bc | 1.15 ab | 1.00 a | 5.86 | 0.008 |
| (1.25–1.71) | (1.16–1.62) | (1.02–1.28) | (0.87–1.13) | |||
| Leaf area index | 3.41 b | 2.98 ab | 2.92 ab | 2.45 a | 3.82 | 0.0113 |
| (2.88–3.95) | (2.45–3.15) | (2.62–3.22) | (2.15–2.75) | |||
| Leaf water potential (MPa) | −2.38 b | −2.83 a | −2.23 b | −2.93 a | 48.7 | <0.001 |
| (−2.32–(−2.15)) | (−3.01–(−2.84)) | (−2.53–(−2.23)) | (−2.98–(−2.68)) | |||
| Relative leaf water content (%) | 68.52 c | 55.10 a | 65.97 c | 60.90 b | 10.29 | <0.001 |
| (64.40–72.65) | (50.97–59.23) | (63.65–68.30) | (58.56–63.21) | |||
| Photosynthetic efficiency | 36.60 a | 36.41 a | 35.58 a | 34.36 a | 2.76 | 0.0439 |
| (34.92–38.25) | (34.74–38.08) | (34.65–36.52) | (33.43–35.30) | |||
| Proline content (mg/g of dw) | 0.47 a | 1.58 b | 0.46 a | 1.51 b | 284.68 | <0.001 |
| (0.36–0.57) | (1.48–1.68) | (0.40–0.52) | (1.45–1.57) | |||
| Biological yield | 310.05 b | 204.57 a | 329.97 b | 323.65 b | 38.67 | <0.001 |
| (82.01–628.33) | (52.17–522.33) | (139.17–758.33) | (136.70–678.02) | |||
| Harvest index | 10.68 a | 17.81 b | 19.66 b | 17.97 b | 9.63 | <0.001 |
| (7.80–13.57) | (14.93–20.70) | (18.04–21.28) | (18.03–19.60) | |||
| Seed yield/Plant (g) | 25.88 a | 25.07 a | 52.90 c | 40.81 b | 81.83 | <0.001 |
| (22.05–29.72) | (21.23–28.90) | (50.74–55.06) | (38.66–42.97) | |||
| Oil content (%) | 29.38 b | 27.30 a | 31.23 c | 30.61 bc | 12.47 | <0.001 |
| (28.22–30.54) | (26.13–28.45) | (30.57–31.88) | (29.96–31.26) |
| Source of Variation | Treatments | Years | Replications/Years | Females | Males | Female (F) × Male (M) | Female × Years | Male × Years | F × M × Years | Error | σ² Female x Males (SCA) | σ² GCA | σ² GCA/σ² SCA | Contribution of Lines | Contribution Testers | Contribution of Lines x Testers | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Degree of freedom | 78 | 1 | 4 | 14 | 3 | 42 | 14 | 3 | 42 | 236 | |||||||
| Number of leaves | N a | 30.52 ** | 105.31 ** | 23.87 ** | 257.30 ** | 74.06 ** | 36.16 ** | 57.45 ** | 50.58 ** | 22.18 | 0.78 | 4.66 | 1.71 | 0.37 | 67.42 | 4.16 | 28.42 |
| S b | 34.63 ** | 1985.12 ** | 94.54 ** | 255.88 ** | 39.75 ** | 35.94 ** | 77.20 ** | 37.75 ** | 19.82 ** | 1.76 | 5.38 | 1.3 | 0.24 | 68.74 | 2.29 | 28.97 | |
| Leaf area | N | 0.10 ** | 1.85 ** | 0.01 ** | 0.28 ** | 0.15 ** | 0.16 ** | 0.06 ** | 0.02 ** | 0.03 | 0.002 | 0.04 | 0.01 | 0.02 | 35.83 | 4.18 | 59.99 |
| S | 0.10 ** | 2.26 ** | 0.01 ** | 0.22 ** | 0.12 ** | 0.13 ** | 0.04 ** | 0.09 ** | 0.05 ** | 0.002 | 0.03 | 0.01 | 0.02 | 34.89 | 3.98 | 61.13 | |
| Specific leaf weight | N | 0.62 ** | 0.55 ** | 0.17 ** | 0.83 ** | 0.67 ** | 0.68 ** | 0.46 ** | 0.20 ** | 0.16 | 0.01 | 0.17 | 0.01 | 0.01 | 27.52 | 4.71 | 67.77 |
| S | 0.52 ** | 0.94 ** | 0.26 ** | 0.74 ** | 0.64 ** | 0.47 ** | 0.40 ** | 0.14 ** | 0.19 ** | 0.01 | 0.09 | 0.01 | 0.03 | 32.47 | 6.04 | 61.49 | |
| Leaf area index | N | 3.29 ** | 57.12 ** | 0.52 ** | 8.59 ** | 4.24 ** | 4.79 ** | 1.67 ** | 0.95 ** | 0.82 | 0.05 | 1.32 | 0.02 | 0.02 | 36 | 3.81 | 60.19 |
| S | 2.79 ** | 47.22 ** | 0.25 ** | 6.36 ** | 3.62 ** | 3.78 ** | 1.55 ** | 0.96 ** | 0.81 ** | 0.06 | 0.99 | 0.01 | 0.01 | 34.44 | 4.2 | 61.37 | |
| Leaf water potential | N | 0.17 ** | 0.14 ** | 0.17 ** | 0.52 * | 0.78 ** | 0.46 ** | 0.05 ** | 0.04 | 0.05 | 0.02 | 0.14 | 0.01 | 0.02 | 24.94 | 8.08 | 66.99 |
| S | 0.28 ** | 0.07 ** | 0.34 ** | 1.01 ** | 1.17 ** | 0.61 ** | 0.31 ** | 1.77 ** | 0.64 ** | 0.005 | 0.01 | 0.03 | 3 | 32.72 | 8.1 | 59.18 | |
| Relative leaf water content | N | 168.79 ** | 393.38 ** | 22.93 ** | 742.36 ** | 311.75 ** | 340.78 ** | 29.29 ** | 22.30 ** | 29.56 | 1.76 | 103.74 | 3.33 | 0.03 | 40.53 | 3.65 | 55.82 |
| S | 208.61 ** | 652.04 ** | 16.52 ** | 716.68 ** | 367.48 ** | 595.45 ** | 17.80 ** | 6.86 * | 29.59 ** | 2.3 | 188.62 | 0.63 | 0.003 | 27.76 | 3.05 | 69.19 | |
| Photosynthetic efficiency | N | 32.40 ** | 37.13 ** | 45.97 ** | 184.11 ** | 236.07 ** | 75.06 ** | 43.92 ** | 44.96 ** | 47.97 ** | 1.92 | 9.03 | 2.43 | 0.27 | 40.03 | 11 | 48.97 |
| S | 24.87 ** | 554.63 ** | 191.26 ** | 82.64 ** | 147.14 ** | 26.99 ** | 30.69 ** | 51.69 ** | 32.93 ** | 2.01 | 1.98 | 1.39 | 0.7 | 42.35 | 16.16 | 41.5 | |
| Proline content | N | 0.01 | 1.74 ** | 0.09 ** | 0.07 ** | 0.06 ** | 0.05 ** | 0.06 ** | 0.06 ** | 0.04 ** | 0.001 | 0.01 | 0.01 | 0.02 | 30.73 | 6.32 | 62.94 |
| S | 0.2 | 0.08 ** | 0.01 * | 0.74 ** | 0.35 ** | 0.50 ** | 0.71 ** | 0.30 ** | 0.48 ** | 0.002 | 0.01 | 0.01 | 0.03 | 31.75 | 3.20 | 65.05 | |
| Biological yield | N | 32,121.15 ** | 2,554,378.00 ** | 717.17 | 53,416.76 ** | 64,517.33 ** | 62,683.68 ** | 61,534.31 ** | 84,260.41 ** | 30,359.24 ** | 411.59 | 10,774.82 | 811.48 | 0.08 | 20.92 | 5.42 | 73.66 |
| S | 24,228.56 ** | 1,858,552.00 ** | 536.71 | 62,167.62 ** | 150,506.50 ** | 53,274.99 ** | 43,248.35 ** | 142,227.60 ** | 43,642.73 ** | 826.16 | 3210.75 | 69.59 | 0.02 | 24.45 | 12.69 | 62.86 | |
| Harvest index | N | 85.23 ** | 5505.00 ** | 22.09 ** | 56.11 ** | 320.93 ** | 160.21 ** | 153.45 ** | 166.44 ** | 108.69 ** | 2.94 | 17.17 | 0.40 | 0.02 | 9.27 | 11.36 | 79.38 |
| S | 110.82 ** | 7137.38 ** | 6.76 | 323.99 ** | 640.69 ** | 122.05 ** | 206.76 ** | 274.21 | 101.06 ** | 5.56 | 7.00 | 3.87 | 0.55 | 39.16 | 16.59 | 44.25 | |
| Seed yield | N | 419.17 ** | 2961.48 ** | 88.81 ** | 543.29 ** | 976.08 ** | 379.31 ** | 145.52 ** | 343.09 ** | 149.26 ** | 13.03 | 76.68 | 5.00 | 0.07 | 28.74 | 11.06 | 60.2 |
| S | 239.13 ** | 372.00 ** | 30.93 | 841.16 ** | 158.49 ** | 338.09 ** | 189.12 ** | 693.05 ** | 130.24 ** | 13.3 | 69.28 | 2.61 | 0.04 | 44.52 | 1.80 | 53.68 | |
| Oil content | N | 8.68 * | 456.52 ** | 1.14 ** | 27.63 ** | 18.94 ** | 16.63 ** | 17.34 ** | 3.20 ** | 17.61 ** | 0.14 | 0.33 | 0.24 | 0.73 | 33.86 | 4.98 | 61.16 |
| S | 15.43 ** | 39.43 ** | 0.54 ** | 79.32 ** | 12.92 ** | 32.99 ** | 41.69 ** | 15.71 ** | 39.02 ** | 0.16 | 2.01 | 0.41 | 0.2 | 43.81 | 1.53 | 54.66 |
| Parents | Number of Leaves | Leaf Area | Specific Leaf Weight | Leaf Area Index | Leaf Water Potential | Relative Leaf Water Content | Photosynthetic Efficiency | Proline Content | Biological Yield | Harvest Index | Seed Yield | Oil Content |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| GCA Under Non-stressed Environment | ||||||||||||
| CMS-XA | −4.43 | −0.11 | −0.06 | −0.61 | 0.001 | −0.75 | 0.79 ** | 0.10 ** | 8.76 * | 0.65 * | −1.3 | 1.40 ** |
| CMS-E002-91A | −2.35 | −0.12 | −0.19 | −0.67 | 0.07 ** | 5.09 ** | −0.46 | 0.07 ** | 26.47 ** | −1.3 | 1.75 ** | −1.45 |
| CMS-PKU-2A | −1.47 | 0.13 ** | 0.37 ** | 0.73 ** | −0.1 | 2.44 ** | −1.3 | −0.07 | −23.55 | 0.14 | 0.16 | −0.21 |
| CMS-ARG-2A | −3.45 | −0.02 | 0.09 ** | −0.11 | −0.11 | 2.06 ** | −2.01 | 0.02* | −40.94 | −1.42 | −2.90 | 1.29 ** |
| CMS-ARG-3A | −3.94 | −0.12 | −0.03 | −0.64 | 0.25 ** | 6.94 ** | −5.98 | −0.03 | 52.69 ** | 1.11 ** | 5.17 ** | −0.24 |
| CMS-ARG-6A | 2.33 ** | −0.03 | −0.12 | −0.31 | −0.08 | 1.50 ** | 0.77 ** | 0.03 ** | −30.93 | 2.65 ** | 7.21 ** | −0.37 |
| CMS-DV-10A | 0.12 | −0.03 | −0.04 | −0.12 | −0.04 | −3.09 | −3.09 | −0.08 | 8.97 * | −0.38 | 0.21 | −0.75 |
| CMS-PHIR-27A | −0.34 | −0.13 | −0.28 | −0.7 | 0.21 ** | 5.03 ** | 1.62 ** | −0.02 | −5.51 | 1.91 ** | 1.00 | −0.81 |
| CMS-PRUN-29A | −0.41 | 0.16 ** | 0.32 ** | 0.89 ** | −0.22 | −5.61 | 3.57 ** | 0.08 ** | 48.11 ** | 0.33 | 1.07 | 1.94 ** |
| CMS-40A | 2.82 ** | −0.08 | −0.24 | −0.4 | 0.04 * | −0.87 | 0.64 ** | −0.02 | −21.66 | 1.00 ** | 3.84 ** | −0.25 |
| CMS-42A | 1.86 ** | 0.14 ** | 0.18 ** | 0.82 ** | 0.14 ** | −8.02 | −2.12 | −0.06 | −9.99 | −0.52 | 3.18 ** | 0.66 ** |
| CMS-234A | 1.58 ** | 0.16 ** | −0.01 | 0.88 ** | 0.20 ** | −8.97 | 5.56 ** | 0.01 | 111.49 ** | −2.2 | 3.39 ** | 0.49 ** |
| CMS-38A | 1.29 ** | 0.09 ** | 0.07 ** | 0.43 ** | −0.18 | −7.76 | 1.25 ** | −0.01 | −81.18 | 0.05 | −7.13 | −0.65 |
| NC-41B (C) | −1.47 | −0.04 | −0.01 | −0.22 | −0.1 | 5.01 ** | 1.35 ** | −0.02 | 5.95 | 1.04 ** | −5.24 | −1.85 |
| 42B | 0.86 ** | 0.001 | −0.06 | 0.01 | −0.07 | 6.98 ** | −0.6 | 0.02 * | −48.68 | −3.05 | −10.4 | 0.81 ** |
| RCR-8297 | 0.26 ** | −0.02 | −0.03 | −0.06 | 0.05 ** | −0.19 | 0.93 ** | 0.001 | −18.32 | 2.54 ** | 3.15 ** | −0.27 |
| P69R | 1.09 ** | 0.03 ** | 0.01 | 0.16 ** | −0.14 | 2.21 ** | 1.67 ** | 0.04 ** | 23.20 ** | −0.85 | −3.5 | −0.14 |
| P124R | −0.29 | −0.05 | −0.09 | −0.28 | 0.02 * | −2.33 | −1.95 | −0.02 | −27.59 | 0.17 | −2.1 | −0.28 |
| P100R | −1.06 | 0.04 ** | 0.11 ** | 0.19 ** | 0.07 ** | 0.30 ** | −0.64 | −0.02 | 22.71 ** | −1.86 | 2.44 ** | 0.68 ** |
| GCA Under Stressed Environment | ||||||||||||
| CMS-XA | −1.36 | −0.08 | −0.12 | −0.37 | −0.1 | 6.28 ** | −0.75 | 0.32 ** | 15.09 ** | 2.34 ** | 6.60 ** | 0.22 ** |
| CMS-E002-91A | 0.19 | 0.04 ** | 0.08 ** | 0.32 ** | 0.15 ** | −7.82 | −0.07 | 0.24 ** | −10.51 * | 1.93 ** | 1.82 ** | 0.36 ** |
| CMS-PKU-2A | −0.98 | 0.10 ** | 0.27 ** | 0.64 ** | 0.12 ** | 4.08 ** | −1.42 | −0.22 | −49.58 | 6.01 ** | −0.8 | 1.86 ** |
| CMS-ARG-2A | −2.78 | −0.07 | −0.02 | −0.29 | 0.41 ** | 5.18 ** | 0.19 | 0.08 ** | −57.42 | 1.54 ** | −3.82 | 1.58 ** |
| CMS-ARG-3A | −1.79 | −0.17 | −0.31 | −0.85 | −0.14 | −2.72 | −1.79 | −0.06 | −13.07 | 1.59 ** | 4.80 ** | 3.23 ** |
| CMS-ARG-6A | 1.99 ** | 0.03 ** | 0.001 | −0.05 | −0.22 | −4.9 | 2.18 ** | −0.09 | 14.80 ** | 0.41 | 3.78 ** | −0.45 |
| CMS-DV-10A | −3.00 | −0.06 | 0.03 | −0.25 | −0.26 | −7.56 | −2.33 | −0.22 | −58.69 | 3.27 ** | −0.04 | 0.68 ** |
| CMS-PHIR-27A | −2.94 | −0.13 | −0.2 | −0.63 | −0.22 | 2.62 ** | −2.27 | −0.04 | −45.97 | 1.88 ** | −0.79 | −1.22 |
| CMS-PRUN-29A | −2.12 | 0.06 ** | 0.22 ** | 0.41 ** | −0.03 | −0.61 | −0.09 | 0.28 ** | 31.44 ** | 3.43 ** | 8.98 ** | 1.15 ** |
| CMS-40A | 3.92 ** | −0.02 | −0.11 | −0.19 | −0.16 | −4.99 | 0.80 ** | −0.19 | 74.96 ** | −2.94 | 0.69 | −0.52 |
| CMS-42A | 1.63 ** | 0.18 ** | 0.29 ** | 1.05 ** | −0.18 | 2.42 ** | −0.82 | −0.18 | −11.28 | −1.72 | 0.53 | −0.58 |
| CMS-234A | 1.53 ** | 0.09 ** | −0.1 | 0.59 ** | 0.20 ** | −2.04 | 4.68 ** | 0.03 ** | 93.32 ** | −2.84 | 3.44 ** | 0.27 ** |
| CMS-38A | 1.42 ** | 0.02* | −0.13 | −0.23 | 0.22 ** | −2.17 | 0.96 ** | 0.02 * | −61.22 | −1.59 | −7.12 | −0.01 |
| NC-41B (C) | −4.81 | −0.06 | 0.15 ** | −0.27 | 0.21 ** | 11.77 ** | 1.50 ** | −0.03 | 7.63 | −7.67 | −15.49 | −4.71 |
| 42B | 3.10 ** | 0.08 ** | −0.07 | 0.11 ** | 0.01 | 0.47 | −0.78 | 0.06 ** | 70.51 ** | −5.63 | −2.59 | −1.85 |
| RCR-8297 | −0.27 | 0.001 | 0.04 ** | 0.08 ** | 0.01 | 1.58 ** | 1.34 ** | 0.03 ** | −28.1 | 2.88 ** | 1.70 ** | 0.34 ** |
| P69R | 0.96 ** | −0.01 | −0.06 | −0.03 | −0.05 | −2.96 | 0.75 ** | 0.07 ** | 60.37 ** | −3.15 | −1.05 | 0.32 ** |
| P124R | −0.13 | −0.04 | −0.08 | −0.26 ** | 0.16 ** | 0.50 ** | −1.46 | −0.03 | −21.67 | 1.38 ** | −1.06 | −0.35 |
| P100R | −0.56 | 0.05 ** | 0.10 ** | 0.21 ** | −0.11 | 0.88 ** | −0.64 | −0.07 | −10.6 | −1.11 | 0.41 | −0.31 |
| Traits | Crosses | N | S |
|---|---|---|---|
| Number of leaves | CMS-38A × RCR-8297 | 4.65 ** | 1.79 ** |
| CMS-ARG-6A × P124R | 3.08 ** | 2.65 ** | |
| CMS-40A × P124R | 2.22 ** | 1.01 ** | |
| Leaf area | CMS-ARG-2A × RCR-8297 | 0.42 ** | 0.38 ** |
| CMS-234A × P100R | 0.34 ** | 0.31 ** | |
| CMS-XA × P69R | 0.23 ** | 0.03 ** | |
| Specific leaf weight | CMS-ARG-2A × RCR-8297 | 0.88 ** | 0.82 ** |
| CMS-XA × P69R | 0.67 ** | 0.13 ** | |
| CMS-234A × P100R | 0.44 ** | 0.45 ** | |
| Leaf area index | CMS-ARG-2A × RCR-8297 | 2.25 ** | 2.01 ** |
| CMS-234A × P100R | 1.90 ** | 1.79 ** | |
| CMS-38A × RCR-8297 | 1.09 ** | 1.13 ** | |
| Leaf water potential | CMS-PRUN-29A × RCR-8297 | 0.45 ** | 0.13 ** |
| CMS-38A × RCR-8297 | 0.40 ** | 0.28 ** | |
| 42B × P69R | 0.33 ** | 0.33 ** | |
| Relative leaf water content | CMS-234A × RCR-8297 | 5.94 ** | 17.21 ** |
| CMS-E002-91A × P69R | 5.77 ** | 7.54 ** | |
| CMS-40A × P69R | 5.70 ** | 7.52 ** | |
| Photosynthetic efficiency | CMS-PHIR-27A × P100R | 4.21 ** | 1.18 ** |
| CMS-38A × RCR-8297 | 3.95 ** | 7.14 ** | |
| CMS-E002-91A × P100R | 1.97 ** | 2.85 ** | |
| Proline content | 42B × RCR-8297 | 0.20 ** | 0.62 ** |
| CMS-XA × P69R | 0.18 ** | 0.60 ** | |
| CMS-PRUN-29A × P124R | 0.14 ** | 0.38 ** | |
| Biological yield | CMS-38A × RCR-8297 | 98.05 ** | 58.42 ** |
| CMS-XA × P124R | 84.12 ** | 58.67 ** | |
| CMS-ARG-6A × P124R | 67.40 ** | 184.79 ** | |
| Harvest index | CMS-PHIR-27A × RCR-8297 | 9.10 ** | 2.47 ** |
| 42B × P100R | 5.39 ** | 2.83 ** | |
| CMS-ARG-3A × P124R | 5.18 ** | 4.42 ** | |
| Seed yield | CMS-ARG-2A × P100R | 9.97 ** | 10.70 ** |
| CMS-PKU-2A × P124R | 8.31 ** | 7.46 ** | |
| 42B × P69R | 8.14 ** | 11.60 ** | |
| Oil content | CMS-E002-91A × P124R | 2.11 ** | 1.62 ** |
| NC-41B (C) × RCR-8297 | 2.08 ** | 3.32 ** | |
| CMS-PRUN-29A × P100R | 2.03 ** | 1.77 ** |
| A/B/R Lines Accessions | Species | Hybrids | |||
|---|---|---|---|---|---|
| RCR-8297 | P69R | P124R | P100R | ||
| A Lines (Alloplasmic) | |||||
| CMS-XA | Unknown | CMS-XA × RCR-8297 | CMS-XA × P69R | CMS-XA × P124R | CMS-XA × P100R |
| CMS-E002-91A | H. annuus L. | CMS-E002-91A × RCR-8297 | CMS-E002-91A × P69R | CMS-E002-91A × P124R | CMS-E002-91A × P100R |
| CMS-PKU-2A | H. annuus | CMS-PKU-2A × RCR-8297 | CMS-PKU-2A × P69R | CMS-PKU-2A × P124R | CMS-PKU-2A × P100R |
| CMS-ARG-2A | H. argophyllus Torr. & A.Gray | ARG-2A × RCR-8297 | CMS-ARG-2A × P69R | CMS-ARG-2A × P124R | CMS-ARG-2A × P100R |
| CMS-ARG-3A | H. argophyllus | CMS-ARG-3A × RCR-8297 | CMS-ARG-3A × P69R | CMS-ARG-3A × P124R | CMS-ARG-3A × P100R |
| CMS-ARG-6A | H. argophyllus | CMS-ARG-6A × RCR-8297 | CMS-ARG-6A × P69R | CMS-ARG-6A × P124R | CMS-ARG-6A × P100R |
| CMS-DV-10A | H. debilis ssp. Vestitus Nutt. | CMS-DV-10A × RCR-8297 | CMS-DV-10A × P69R | CMS-DV-10A × P124R | CMS-DV-10A × P100R |
| CMS-PHIR-27A | H. praecox ssp. Hirtus Engelm. & A.Gray | CMS-PHIR-27A × RCR-8297 | CMS-PHIR-27A × P69R | CMS-PHIR-27A × P124R | CMS-PHIR-27A × P100R |
| CMS-PRUN-29A | H. praecox ssp. Runyonii Engelm. & A.Gray | CMS-PRUN-29A × RCR-8297 | CMS-PRUN-29A × P69R | CMS-PRUN-29A × P124R | CMS-PRUN-29A × P100R |
| A Lines (Euplasmic) | |||||
| CMS-40A | H. petiolaris (conventional) Siebold & Zucc. | CMS-40A × RCR-8297 | CMS-40A × P69R | CMS-40A × P124R | CMS-40A × P100R |
| CMS-42A | H. petiolaris (conventional) | CMS-42A × RCR-8297 | CMS-42A × P69R | CMS-42A × P124R | CMS-42A × P100R |
| CMS-234A | H. petiolaris (conventional) | CMS-234A × RCR-8297 | CMS-234A × P69R | CMS-234A × P124R | CMS-234A × P100R |
| CMS-38A | H. petiolaris (conventional) | CMS-38A × RCR-8297 | CMS-38A × P69R | CMS-38A × P124R | CMS-38A × P100R |
| B Lines (Maintainer) | |||||
| NC-41B | H. petiolaris (conventional) | NC-41B × RCR-8297 | NC-41B × P69R | NC-41B × P124R | NC-41B × P100R |
| 42B | H. petiolaris (conventional) | 42B × RCR-8297 | 42B × P69R | 42B × P124R | 42B × P100R |
| R Lines (Restorer) | |||||
| RCR-8297 | H. annuus | ||||
| P69R | H. annuus | ||||
| P124R | H. annuus | ||||
| P100R | H. annuus | ||||
| Soil Property | Value |
|---|---|
| Sand (per cent) | 82.2 |
| Silt (per cent) | 7.1 |
| Clay (per cent) | 10.7 |
| Textural class | loamy sand |
| Soil temperature (mean) | 13 °C |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tyagi, V.; Dhillon, S.K.; Kaushik, P.; Kaur, G. Characterization for Drought Tolerance and Physiological Efficiency in Novel Cytoplasmic Male Sterile Sources of Sunflower (Helianthus annuus L.). Agronomy 2018, 8, 232. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8100232
Tyagi V, Dhillon SK, Kaushik P, Kaur G. Characterization for Drought Tolerance and Physiological Efficiency in Novel Cytoplasmic Male Sterile Sources of Sunflower (Helianthus annuus L.). Agronomy. 2018; 8(10):232. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8100232
Chicago/Turabian StyleTyagi, Vikrant, Satwinder Kaur Dhillon, Prashant Kaushik, and Gurpreet Kaur. 2018. "Characterization for Drought Tolerance and Physiological Efficiency in Novel Cytoplasmic Male Sterile Sources of Sunflower (Helianthus annuus L.)" Agronomy 8, no. 10: 232. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8100232