Hydraulic Redistribution in Slender Wheatgrass (Elymus trachycaulus Link Malte) and Yellow Sweet Clover (Melilotus officinalis L.): Potential Benefits for Land Reclamation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
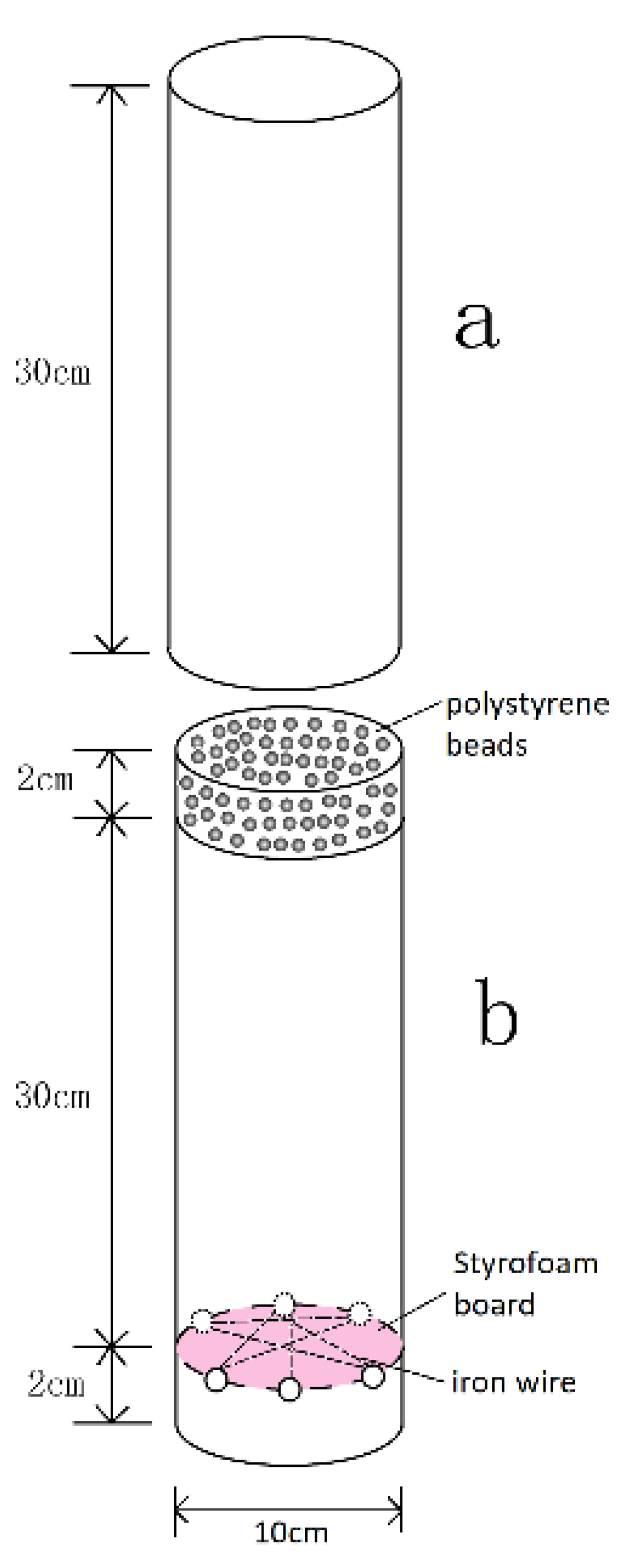
2.1. Split-Root Setup and Plant Material
2.2. Experimental Design and Deuterium (D)-Enriched Water Treatments
2.3. Hydrogen Isotope Analysis
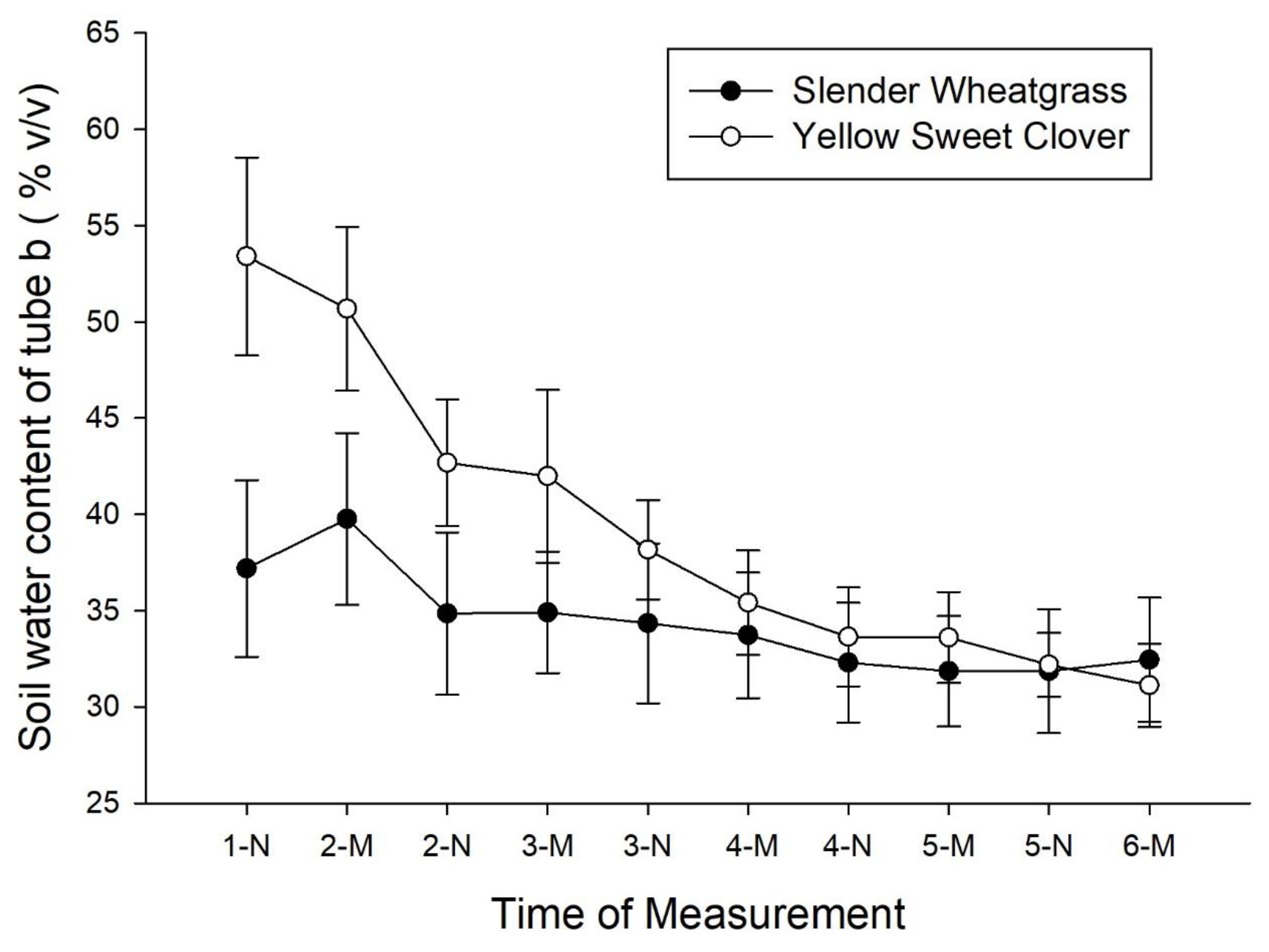
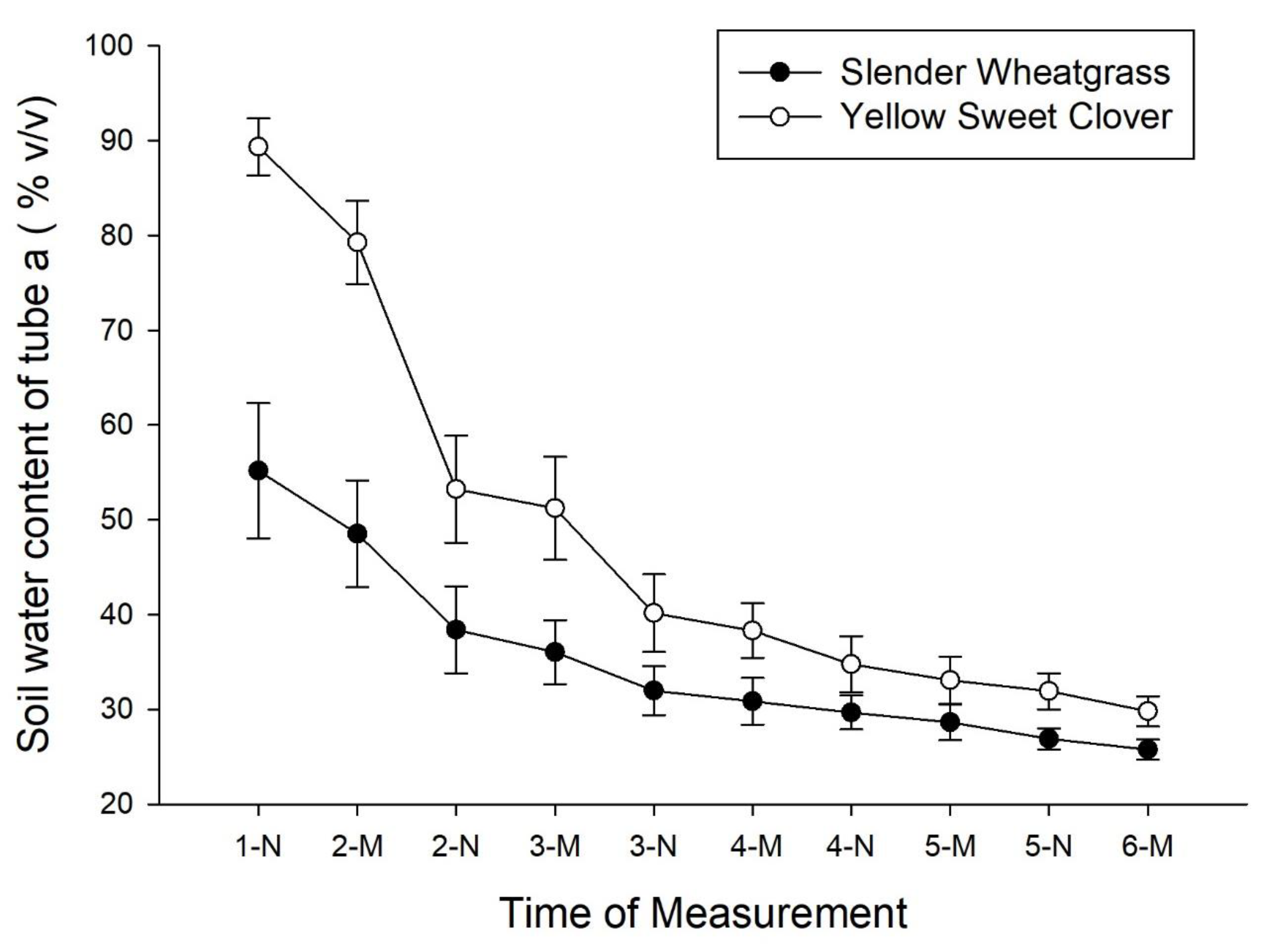
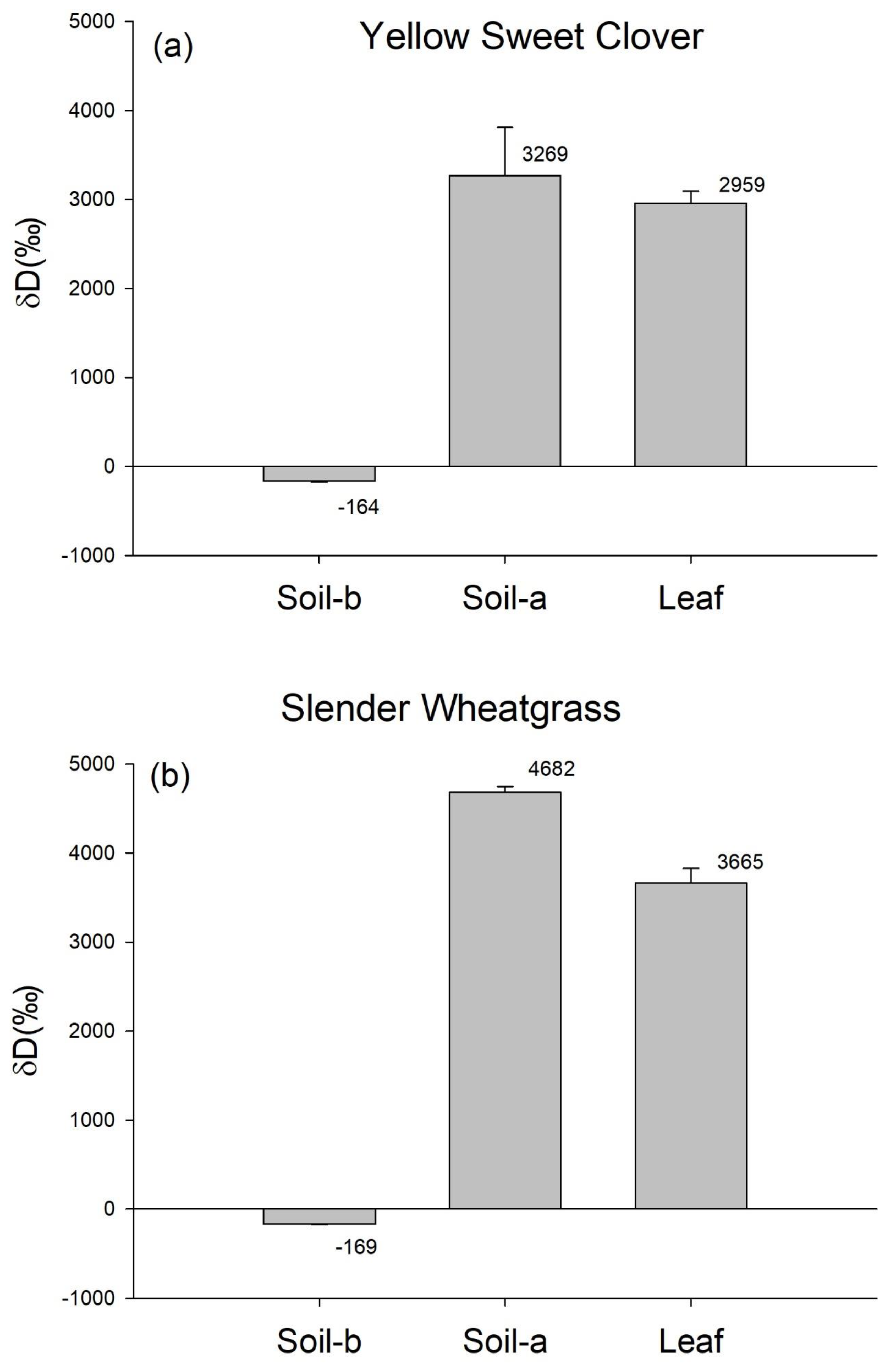
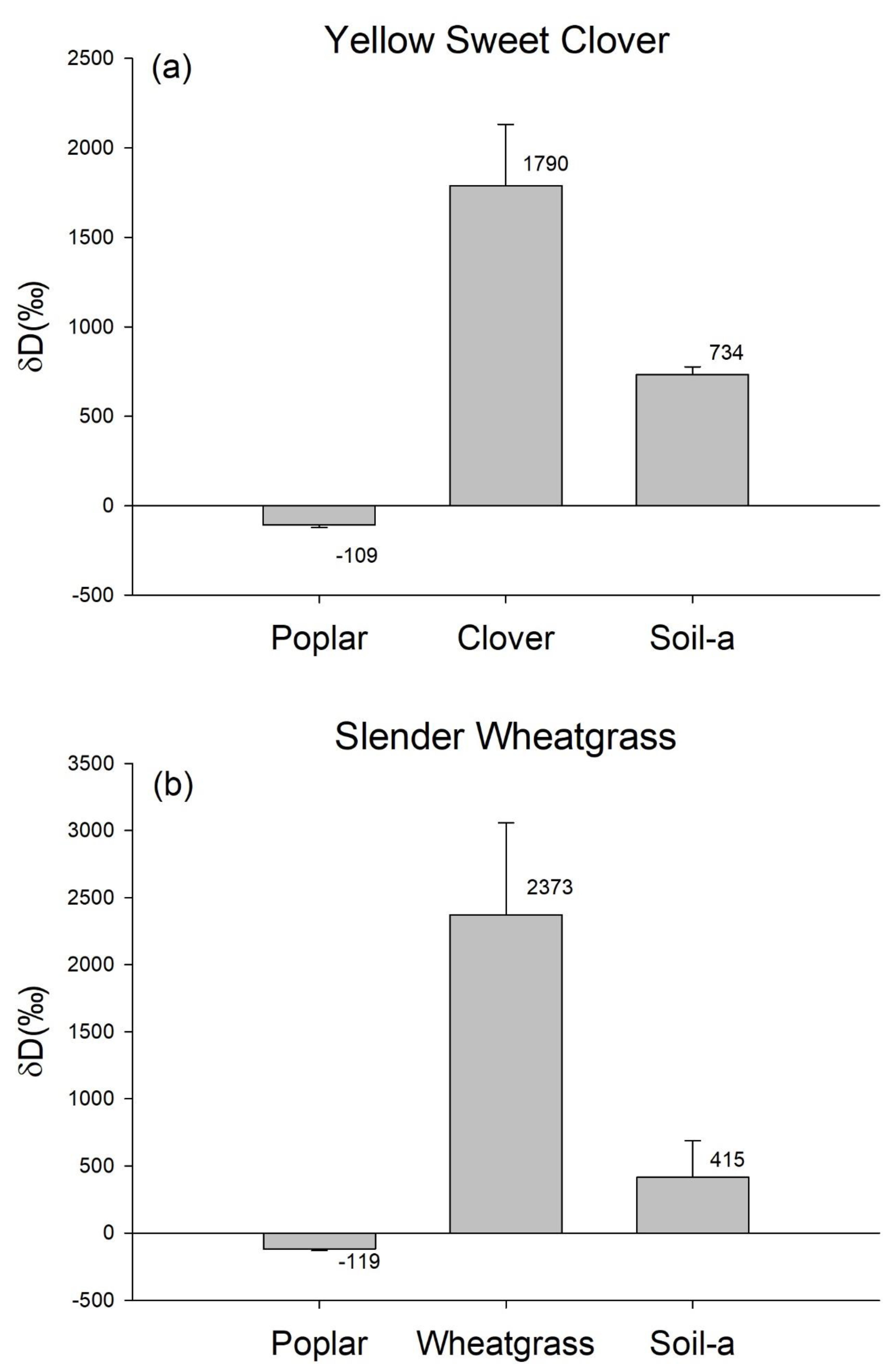
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Prieto, I.; Armas, C.; Pugnaire, F.I. Water release through plant roots: New insights into its consequences at the plant and ecosystem level. New Phytol. 2012, 193, 830–841. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, M.M.; Dawson, T.E.; Richards, J.H. Hydraulic lift: Consequences of water efflux from the roots of plants. Oecologia 1998, 113, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Ryel, R.J.; Caldwell, M.; Leffler, A.; Yoder, C. Rapid soil moisture recharge to depth by roots in a stand of Artemisia tridentata. Ecology 2003, 84, 757–764. [Google Scholar] [CrossRef]
- Burgess, S.S.; Adams, M.A.; Turner, N.C.; Ong, C.K. The redistribution of soil water by tree root systems. Oecologia 1998, 115, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Bauerle, T.L.; Richards, J.H.; Smart, D.R.; Eissenstat, D.M. Importance of internal hydraulic redistribution for prolonging the lifespan of roots in dry soil. Plant Cell Environ. 2008, 31, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Domec, J.C.; Scholz, F.; Bucci, S.; Meinzer, F.; Goldstein, G.; Villalobos-Vega, R. Diurnal and seasonal variation in root xylem embolism in neotropical savanna woody species: Impact on stomatal control of plant water status. Plant Cell Environ. 2006, 29, 26–35. [Google Scholar] [CrossRef]
- Querejeta, J.; Egerton-Warburton, L.M.; Allen, M.F. Direct nocturnal water transfer from oaks to their mycorrhizal symbionts during severe soil drying. Oecologia 2003, 134, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, M.M.; Richards, J.H. Hydraulic lift: Water efflux from upper roots improves effectiveness of water uptake by deep roots. Oecologia 1989, 79, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, F.; Dawson, T.; Prins, H.; Berendse, F.; Kroon, H. Below-ground competition between trees and grasses may overwhelm the facilitative effects of hydraulic lift. Ecol. Lett. 2004, 7, 623–631. [Google Scholar] [CrossRef]
- Brooks, J.R.; Meinzer, F.C.; Warren, J.M.; Domec, J.C.; Coulombe, R. Hydraulic redistribution in a Douglas-fir forest: Lessons from system manipulations. Plant Cell Environ. 2006, 29, 138–150. [Google Scholar] [CrossRef]
- Priyadarshini, K.V.R.; Prins, H.H.T.; de Bie, S.; Heitkönig, I.M.A.; Woodborne, S.; Gort, G.; Kirkman, K.; Ludwig, F.; Dawson, T.E.; De Kroon, H. Seasonality of hydraulic redistribution by trees to grasses and changes in their water-source use that change tree-grass interactions. Ecohydrology 2015, 9, 218–228. [Google Scholar] [CrossRef]
- Sekiya, N.; Yano, K. Do pigeon pea and sesbania supply groundwater to intercropped maize through hydraulic lift?—Hydrogen stable isotope investigation of xylem waters. Field Crop. Res. 2004, 86, 167–173. [Google Scholar] [CrossRef]
- Sekiya, N.; Araki, H.; Yano, K. Applying hydraulic lift in an agroecosystem: Forage plants with shoots removed supply water to neighboring vegetable crops. Plant Soil 2011, 341, 39–50. [Google Scholar] [CrossRef]
- Kizito, F.; Sene, M.; Dragila, M.; Lufafa, A.; Diedhiou, I.; Dossa, E.; Cuenca, R.; Selker, J.; Dick, R. Soil water balance of annual crop–native shrub systems in Senegal’s Peanut Basin: The missing link. Agric. Water Manag. 2007, 90, 137–148. [Google Scholar] [CrossRef]
- Pang, J.; Wang, Y.; Lambers, H.; Tibbett, M.; Siddique, K.H.; Ryan, M.H. Commensalism in an agroecosystem: Hydraulic redistribution by deep-rooted legumes improves survival of a droughted shallow-rooted legume companion. Physiol. Plant. 2013, 149, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Pokharel, P.; Chang, S. Exponential fertilization promotes seedling growth by increasing nitrogen retranslocation in trembling aspen planted for oil sands reclamation. For. Ecol. Manag. 2016, 372, 35–43. [Google Scholar] [CrossRef]
- Zhang, W.; Zwiazek, J.J. Responses of Reclamation Plants to High Root Zone pH: Effects of Phosphorus and Calcium Availability. J. Environ. Qual. 2016, 45, 1652–1662. [Google Scholar] [CrossRef]
- Abouguendia, Z.M.; Redmann, R.E. Germination and Early Seedling Growth of Four Conifers on Acidic and Alkaline Substrates. For. Sci. 1979, 25, 358–360. [Google Scholar]
- Fung, M.Y.P.; Macyk, T.M. Reclamation of oil sands mining areas. In Reclamation of Drastically Disturbed Lands; Barnhisel, R.I., Darmody, R.G., Daniels, W.L., Eds.; American Society of Agronomy, Crop Science Society of America, Soil Science Society of America: Madison, WI, USA, 2000; pp. 755–774. [Google Scholar]
- Quoreshi, A.M. The use of mycorrhizal biotechnology in restoration of disturbed ecosystem. In Mycorrhizae: Sustainable Agriculture and Forestry; Springer: New York, NY, USA, 2008; pp. 303–320. [Google Scholar]
- Zhang, W.; Calvo-Polanco, M.; Chen, Z.C.; Zwiazek, J.J. Growth and physiological responses of trembling aspen (Populus tremuloides), white spruce (Picea glauca) and tamarack (Larix laricina) seedlings to root zone pH. Plant Soil 2013, 373, 775–786. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Sanchez-Romera, B.; Aroca, R. Mild salt stress conditions induce different responses in root hydraulic conductivity of phaseolus vulgaris over-time. PLoS ONE. 2014, 9, e90631. [Google Scholar] [CrossRef]
- He, H.; Dyck, M.F.; Horton, R.; Ren, T.; Bristow, K.L.; Lv, J.; Si, B. Development and application of the heat pulse method for soil physical measurements. Rev. Geophys. 2018, 56. [Google Scholar] [CrossRef]
- West, A.G.; Patrickson, S.J.; Ehleringer, J.R. Water extraction times for plant and soil materials used in stable isotope analysis. Rapid Commun. Mass Spectrom. 2006, 20, 1317–1321. [Google Scholar]
- Ehleringer, J.; Dawson, T. Water uptake by plants: Perspectives from stable isotope composition. Plant Cell Environ. 1992, 15, 1073–1082. [Google Scholar] [CrossRef]
- Baker, J.; Allmaras, R. System for automating and multiplexing soil moisture measurement by time-domain reflectometry. Soil Sci. Soc. Am. J. 1990, 54, 1–6. [Google Scholar] [CrossRef]
- Ludwig, F.; Dawson, T.E.; Kroon, H.; Berendse, F.; Prins, H.H.T. Hydraulic lift in Acacia tortilis trees on an East African savanna. Oecologia 2003, 134, 293–300. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.-Q.; Zwiazek, J.J. Hydraulic Redistribution in Slender Wheatgrass (Elymus trachycaulus Link Malte) and Yellow Sweet Clover (Melilotus officinalis L.): Potential Benefits for Land Reclamation. Agronomy 2018, 8, 308. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8120308
Zhang W-Q, Zwiazek JJ. Hydraulic Redistribution in Slender Wheatgrass (Elymus trachycaulus Link Malte) and Yellow Sweet Clover (Melilotus officinalis L.): Potential Benefits for Land Reclamation. Agronomy. 2018; 8(12):308. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8120308
Chicago/Turabian StyleZhang, Wen-Qing, and Janusz J. Zwiazek. 2018. "Hydraulic Redistribution in Slender Wheatgrass (Elymus trachycaulus Link Malte) and Yellow Sweet Clover (Melilotus officinalis L.): Potential Benefits for Land Reclamation" Agronomy 8, no. 12: 308. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy8120308