Didymella pinodes Affects N and P Uptakes and Their Efficiencies in a Tripartite Mutualism of Pea



1
Department of Crop Sciences, University of Natural Resources and Life Sciences, Gregor Mendel Strasse 33, Vienna 1180, Austria
2
Department of Ecogenomics and Systems Biology, University of Vienna, Althanstraße 14, Vienna 1090, Austria
*
Author to whom correspondence should be addressed.
Agronomy 2019, 9(2), 52; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9020052
Submission received: 13 November 2018
/
Revised: 16 January 2019
/
Accepted: 23 January 2019
/
Published: 26 January 2019
(This article belongs to the Special Issue Symbiotic Associations of Plants with Beneficial Microbes: Perspectives and Challenges for Agronomical Applications)
Abstract
:In pea (Pisum sativum L.) production, Didymella pinodes (Berk. & A. Bloxam) Petr. is the most damaging aerial pathogen globally. In two completely randomized pot experiments with four replicates, we studied the effects of D. pinodes infection interaction with three symbiotic treatments (Rhizobium leguminosarum biovar viciae, arbuscular mycorrhizal fungi (AMF) and co-inoculation of both) and a non-symbiotic control on one or two pea cultivars. Grain yield and yield components of pea, uptakes and physiological efficiencies of N and P and nitrogen fixation were recorded. The results show that there were significant interaction effects among treatments. Therefore, productivity of crops and their uptakes and efficiencies of N and P are dependent on plant health conditions, effectiveness of microbial symbionts and response of pea genotypes. For cv. Protecta inoculated with both symbionts, pathogen infection compared to healthy plants significantly enhanced P acquisition. Overall, plants inoculated with rhizobia alone had higher grain yield by 20–30% and nitrogen fixation by 20–25% than in dual symbiosis independent of plant health conditions. In conclusion, aerial pathogen, pea genotypes and microbial symbionts interactions modified N and P uptake and their efficiencies, which can lead to improving final grain yield quantity and quality in a sustainable farming system.
1. Introduction
Ascochyta blight, caused by Didimella pinodes (syn. Mycosphaerella pinodes) (Berk. & A. Bloxam) Vestergr, is one of the most economically damaging fungal pathogens in pea cultivation across the globe characterized by rapid and widespread death of plant tissue. It causes up to 50% [1] and in severe cases up to 70% pea yield losses through reduction of photosynthetic efficiency [2]. In addition to such pathogen attacks, production of pea can also be co-limited by the availability of nutrients when growings in nitrogen (N) and/or phosphorus (P) deficient soils. In such soils, pea is often considered as an efficient nitrogen-fixing crop that can accumulate more than 300 kg N ha−1 in the aboveground parts in symbiotic associations with rhizobia [3,4]. Furthermore, this legume crop can form mutualistic associations with arbuscular mycorrhiza (AM) fungi, which play vital roles in influencing the availability of plant nutrients with low mobility in soil solution such as P [5], uptake of water [6] and N [7] and promotion of plant growth due to improved uptake of nutrients [8].
The productivity of pea genotypes depends on the compatibility and effectiveness of the AM fungi and the Rhizobium populations in the rhizosphere. Several previous reports claim synergistic effects of co-inoculation of rhizobia and AM fungi on legume growth, nutrient uptake, nodulation and N2-fixation [9,10,11,12,13,14]. Other researchers presented a negative effect of mycorrhiza on nodule development and plant growth [15,16,17]. Recently, it has been underlined that microbial symbionts can enhance green area growth and trigger systemic induced resistance of plants against stress [18,19]. Thus, symbionts may also qualify the host plants to defend themselves or resist against a broad range of abiotic and biotic stresses such as pathogens or herbivore insects [20,21], particularly root pathogens mostly due to direct competition for root space and resources [22]. We also previously reported that these two symbionts can influence the leaf metabolome and proteome differently after infection of pea plants by D. pinodes [23,24], but evidence is lacking whether this association contributes similarly or differently to nitrogen or phosphorous nutrition in a legume plant.
Thus far, yield losses caused by this ascochyta blight are controlled through improved genetic resistance and best management practices such as crop rotation and fungicide applications. It is clear that extensive fungicide use increases risks for environmental pollution and development of fungicide resistant strains. For instance, the loss of disease control and resistance to strobilurin fungicides is documented in Cercospora sojina K. Hara, the causal agent of Frogeye leaf spot in soybean [25]. The overall role and mechanisms of plant nutrient–pathogen interactions is still not well understood. Furthermore, the importance of symbiotic association between rhizobia and AM fungi and among other beneficial microorganisms in view of crop resilience against aerial fungal pathogen stresses has not been studied often.
Indeed, to reduce fertilizer usage and extensive pesticide applications and frequent breakdown of major resistance genes in crop species, there is a growing demand to integrate host plant resistance and the use of beneficial microbes. Thus, we aimed to develop an integrated D. pinodes control approach through evaluation of a resistant and a susceptible pea cultivar with and without microbial symbionts under attack by D. pinodes. Here, we argue that with or without such aerial pathogen attack, the association of these two symbionts cannot separately affect the concentration and the efficiencies of NP nutrition and productivity of pea plant. The main objective of this investigation was to assess the effects of single and double inoculation of pea with Rhizobium leguminosarum bv. viceae and the AM fungus Funneliformis mosseae (T.H. Nicolson & Gerd.) C. Walker & A. Schüssler on enhanced host plant defense or stress resistance capacity by determining grain yield and yield components, N and P uptake and nutrient efficiency traits. This work provides new understanding and insights into the use of these widespread microbes and their potential benefits to increase vegetal protein supply and security in a sustainable agricultural system.
2. Materials and Methods
2.1. Experimental Design
In an effort to investigate whether a single or double inoculation of microbial symbionts affect the resistance, final yield, yield components, N and P uptakes and physiological efficiencies of pea, twice and independently, a factorial experimental design with one field pea genotype (Messire) and two different ones (Messire and Protecta) was established. In both experiments, the plants were treated with or without pathogen infections as well as with five different NP sources including an unfertilized control that was merely used as a baseline to compare the other four NP groups and assess their effect, arbuscular mycorrhiza fungus Funneliformis mosseae (M), Rhizobium leguminosarum bv. viceae (R), dual inoculation with F. mosseae and R. leguminosarum (MR) and synthetic mineral nitrogen and phosphorus fertilization (SF). The experiments were conducted in a completely randomized design with four replicates under controlled greenhouse conditions, as described previously by Desalegn et al. [23] and Turetsheck et al. [24].
2.2. Experimental Soil, Symbiont and Nutrient Management
Soil was collected from the 0–25 cm horizon of an experimental field in Tulln, Austria. It was air-dried, sieved to pass a 2 mm sieve, mixed with expanded clay and silica sand (1:1:1 w/w/w), and then sterilized at 121 °C in an autoclavable plastic bag for 20 min. Three-liter black plastic pots, which had been autoclaved prior to planting, were filled with 2 kg growing substrate mix obtained as described above and moistened with 400 mL sterilized deionized water. To maintain the moisture content at optimum levels, pots were irrigated every other day with sterilized deionized water until a drop came out from their bottom. The soil material was low in both plant available N and P [23]. In such soils, a starter dose of N and P can stimulate seedling growth and early nodulation such that both N2-fixation and eventual yield are enhanced. After planting, therefore, each pot received starter doses of N (21 kg N ha−1) as a combination of CaNO3 and KNO3, as described by Hoffmann et al. [26], and P (20 kg P ha−1) from K2HPO4 during the first week. According to Broughton and Dilworth [27], N- and P-free nutrient solutions were prepared. After preparing each of the above-mentioned N and P fertilizers in a deionized and sterile water in separate bottles, additional 130 kg N ha−1 for M and SF treatment groups and 20 kg P ha−1 for R and SF pots were applied at three successive times (Weeks 3, 5 and 7) during vegetative growth. Once a week, 10 mL of additional N- or P-free nutrient solution was added to the surface of all treatments.
2.3. Biological Materials
The biological materials used in this study were the same as those evaluated for mycorrhizal and rhizobial status by Desalegn et al. [23] and Turetschek et al. [24]. Uniform and visibly healthy seeds were selected and washed with 70% ethanol for 30 s followed by surface sterilization with 12% commercial bleach for 5 min. They were subsequently rinsed six times with exchanged sterilized distilled water, and pre-germinated in perlite (previously autoclaved for 20 min at 121 °C).
Five three-day-old pre-germinated seeds with 4 mm long initial radicle, obtained as described above, were planted per pot. A commercial inoculant including a R. leguminosarum bv. viceae strain selected for maximum N2-fixation potential and Vaminoc, which contains a F. mosseae fungus species, from Becker Underwood Ltd. (www.beckerunderwood.com, now BASF, New York, NY, USA) Ludwigshafen, Germany, were applied according to the company recommendations and procedures. To avoid potential cross-contamination of microbial symbionts among treatments, planting was started with both non-inoculated groups (control and NP treatment).
Pathogen inoculum preparation and infection were performed as described by Desalegn et al. [23]. The fungal pathogen isolate of D. pinodes was multiplied on a Potato Dextrose Agar (PDA) medium amended with Ampicillin (100 μg per mL) and Chloramphenicol (8 μg per mL) at 22 °C and 12 h photoperiod [28]. Spore suspensions were prepared by flooding the surface of 10-day-old cultures with sterile deionized water, then gently scraping the colony with a flathead sterile spatula and finally filtering through two layers of sterile cheesecloth.
After adjusting the concentration of conidia in the suspension to 3 × 105 spores/mL, four-week-old plants were infected with this suspension containing Tween-20 (120 μL per 100 mL of suspension). The non-inoculated plants were also sprayed with a similar volume of sterile deionized water and Tween-20. Before spraying, the two main treatments (healthy and diseased plant groups) were separately covered by a transparent plastic film: (i) to avoid cross-contamination; and (ii) to facilitate the pathogen infection during the first 48 h. Additionally, deionized sterile water was sprayed several times during that period to maintain high relative humidity (95–100%). The two main groups were kept separately in a glasshouse on benches at 21 ± 2 °C with a 12 h photoperiod and 60–70% relative humidity, while their bench and the position of each pot were changed every week.
2.4. Plant Harvest and Nutrient Measurements
At pea maturity with 80 days for cv. Messire and 86 days for cv. Protecta after planting, plants were cut at ground level, weighed and their pods and residual shoot parts separated, number of seeds and pods plant-1 were counted and weighed, and dry matter (DM) of all fractions was determined after drying them separately at 40 °C in a drying oven for five days.
Seed samples were ground using a stone mill. Subsequently, the nitrogen (N) concentrations were measured using a CNS-2000 high temperature combustion elemental analyzer (Leco Corporation, Saint Joseph, MI, USA). In pot experiments, it should be possible to control all inputs and outputs of N, so the N balance method can provide a useful, integrated estimate of N2-fixation (N-fixed) using the following equation:
Amount of fixed N = N yield in N fixing plant − N yield in non-rhizobial (control) plant
Total P was determined after samples digested with aquaregia (100 mg sample with 4.5 mL 37% HCl + 1.5 mL 65% HNO3). Digestions of samples were performed in a Multiwave 3000 microwave system (Anton Paar GmbH, Graz, Austria) using Teflon liners under the following conditions: 1400 W (for 16 tubes), 10 min ramp time, 40 min hold time and 10 min cooling time. Pressure and temperature were regulated with threshold values of 40 bar and 210 °C, respectively, above which heating was suspended. In the digests, P concentration was measured by ICP-OES (Optima 8300, Perkin Elmer, Waltham, MA, USA) at 213.617 nm with matrix-matched standards. To compare the effectiveness of AMF on P supply, the following equation was also used.
Amount of PU = P yield in mycorrhizal plant − P yield in non-mycorrhizal (control) plant
To assess the ability of plants to transform nutrients acquired from N and P sources into economic yield under diseased and healthy conditions, the physiological efficiency (pe) of N (peN) and of P (peP) were calculated according to Delogu et al. [29] using the following equation:
where Y is the grain yield of crop with NP sources (i.e., M, R, MR or SF); Y0 is the grain yield of control crop with no N or P nutrient applied and non-symbiont; U is the N or P uptake in grain of crop with NP sources; U0 is the N or P uptake in grain of control crop with no N or P nutrient applied and non-symbiont; and physiological efficiency values are ratios on a mass basis and are therefore unitless.
Physiological efficiency (pe) = (Y − Y0)/(U − U0)
2.5. Statistical Analyses
Two-way ANOVA or three-way ANOVA was used to examine the main and interaction effects of plant pathogen, nutrient (NP) source and Pisum sativum genotype on grain yield and yield components as well as NP uptake and physiological efficiency. To study the association of these traits, Pearson’s linear correlation coefficients were calculated. Besides, data were analyzed with one-way ANOVA tests to estimate the effects of symbionts on nitrogen fixation and P acquisition. Differences between treatments were compared with Tukey’s multiple range test and statistical significances were defined at p ≤ 0.05, p ≤ 0.01, and p ≤ 0.001.
3. Results
As we examined the interaction effects of NP regimes and pathogen infections as well as cultivars of Pisum sativum on grain yield, yield components, NP uptakes and the physiological efficiencies of N and P, a wide range of main effects and interactions were noted among treatment factors. Our result section focuses on the main interactions with regard to impact and relevance for our research questions.
3.1. Effects of N and P Regime, Pathogen Infection and Genotype on Pea Productivity
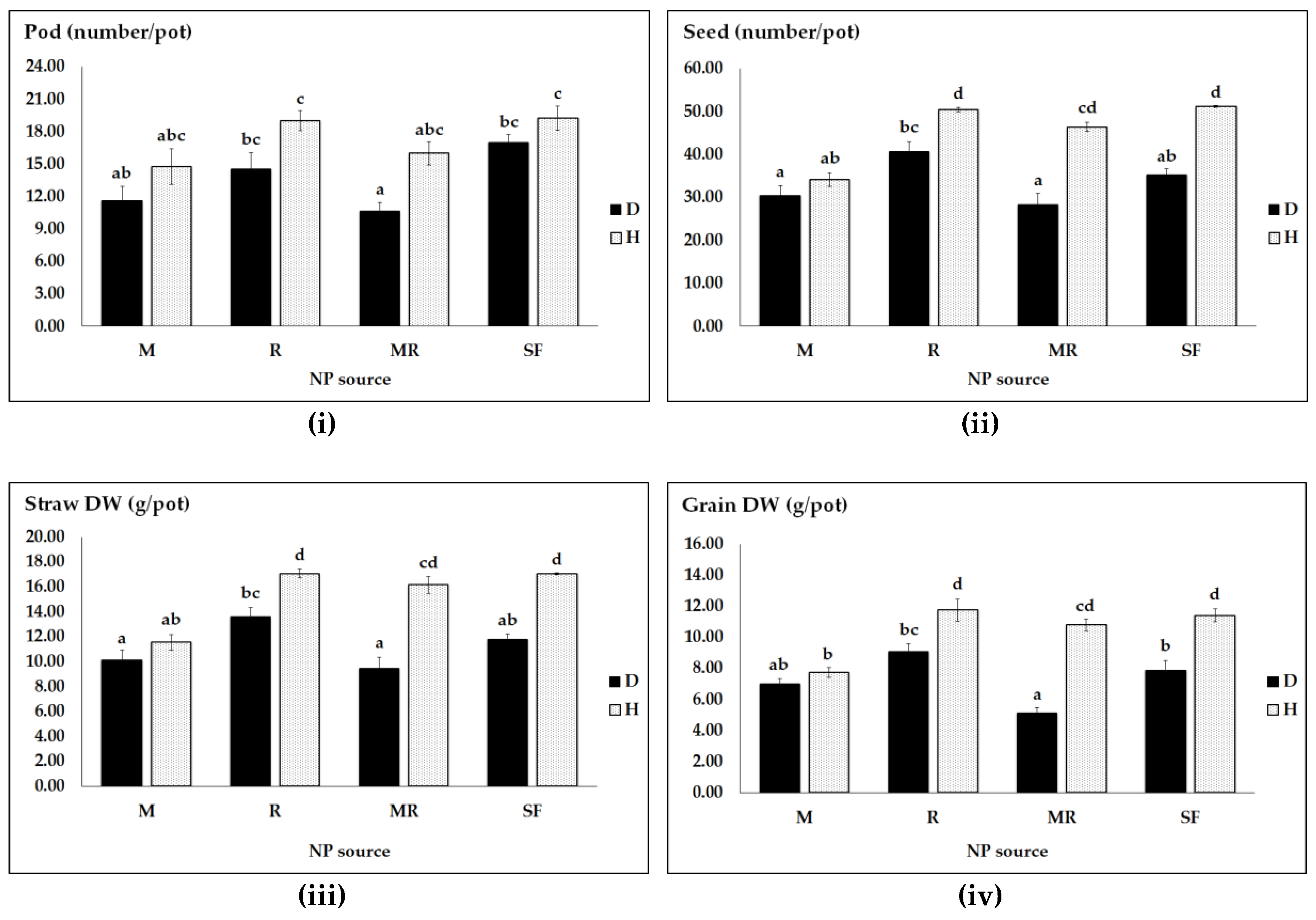
In this study, two-way ANOVA (Table 1 and Table 3) and three-way ANOVA (Table 2 and Table 5) showed that interaction effects were statistically significant. In the first experiment, pathogen infection X NP source interactions had significant effects on pea productivity (i.e., seed number, grain and straw accumulation), but not on pod number (Table 1). However, for pathogen infected pea plants, both single M and single R had virtually similar impacts on pea productivity (Figure 1i,iii,iv), but not on seed number (Figure 1ii). In healthy plants, a combination of the two symbionts (MR) led to significantly higher seed numbers and straw accumulation as well as grain yield than a single M inoculation (Figure 1ii–iv).
However, under healthy plant growth conditions, Figure 1 shows that the dual MR inoculation caused significantly lower results of seed number, grain yield and straw production than a single R inoculation. Overall, under both health conditions, only rhizobia inoculation significantly increased the number of seeds and pods as well as straw accumulation and seed yield as compared to the AMF inoculated plants.
Furthermore, for the main factor pathogen, the average pod number and seed number, straw accumulation and grain yield were significantly higher by 30%, 35%, 40% and 45% in healthy plants than in the diseased pea plants, respectively (Figure 1).
Table 2 shows that the threefold (2 pathogens × 2 cultivars × 4 NP sources) interaction effects significantly affected grain yield and yield components in the second experiment.
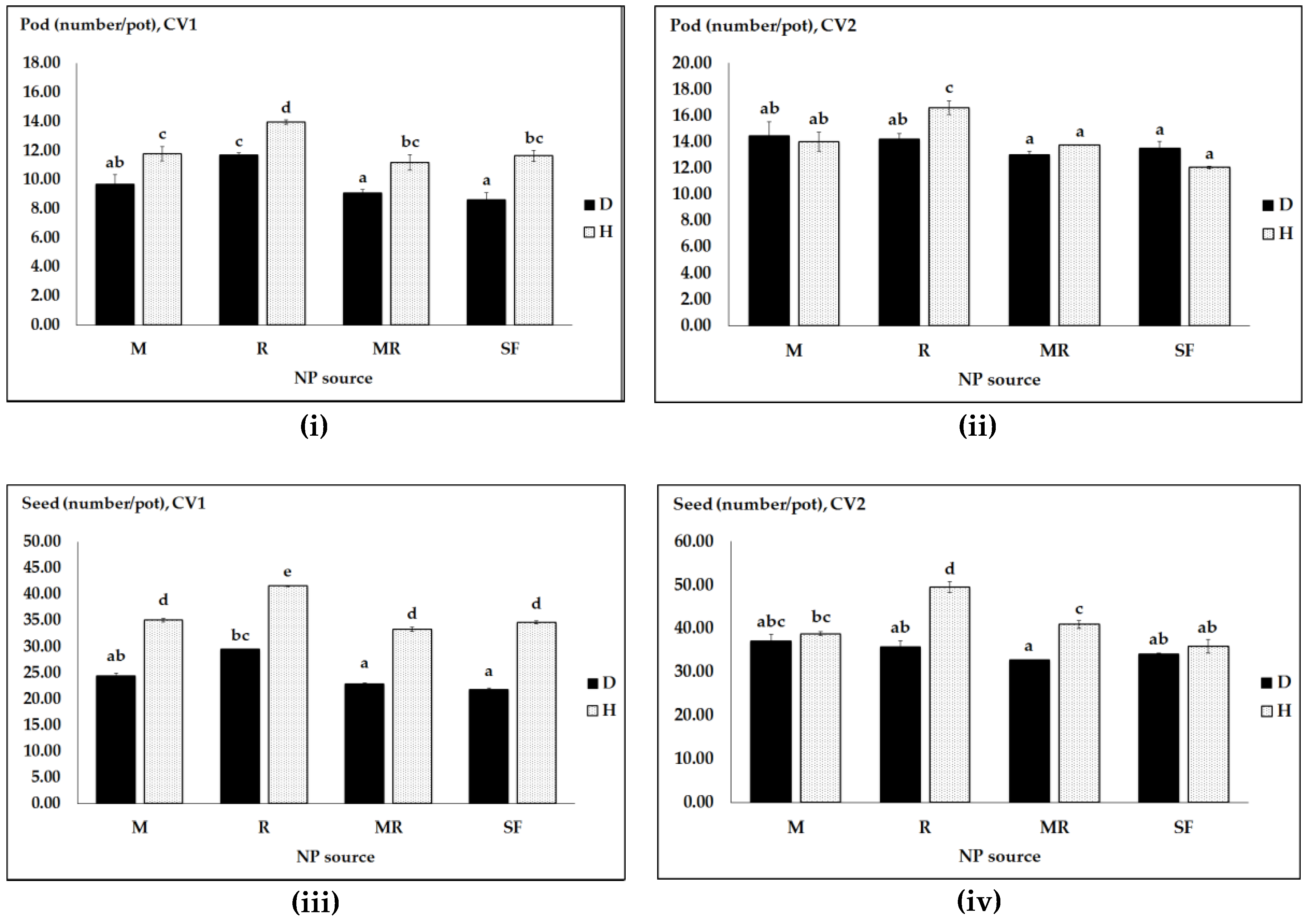
Pea plants inoculated with rhizobia showed higher seed number and grain yield than compared to the other NP regimes (M, MR and synthetic NP fertilizers) in the healthy cv. Protecta that is a partially resistant genotype (Figure 2iv,viii). However, in this cultivar for M and synthetic fertilizers treatments, seed numbers and grain yield were virtually identical for both diseased and healthy plants. In general, these data show that pathogen infected and healthy plants were performing equally with M and synthetic fertilizers, while R favorably affected the healthy plants compared to the diseased plants.
In cv. Messire, for each of the NP sources, pea productivity was consistently lower in the pathogen infected plants as compared to healthy plants (Figure 2i,iii,v,vii). Only if Protecta had been inoculated with single M and fertilized with SF, it had virtually equal SN in both healthy and D. pinodes infected plants (Figure 2iv).
In CV1, for the main factor pathogen, means of pod number, seed number, straw accumulation and grain yield were also significantly lower by 25%, 45%, 31% and 42% in the diseased plants than in healthy ones (Figure 2i,iii,v,vii), but such high levels of pathogen infection effects on reduction of CV2 productivity were not shown (Figure 2ii,iv,vi,viii). Similarly, for the main factor pea genotype, overall increases of crop productivity of CV2 over CV1 were more than 25%.
3.2. Effects of N and P Sources, Pathogen Infection and Genotype on N and P Uptake and Efficiency
In addition, with regard to NP uptakes and their physiological efficiencies, Table 3 and Table 5 show that there were also significant interaction effects between pathogen infection and NP sources and among cultivars, pathogen infection and NP sources, respectively. For pathogen infected pea plants compared to healthy ones, both N uptake and P uptake were much lower in a single M treatment than in R, and NP synthetic fertilizers (Table 4). Additionally, for single M inoculated plants, the mean NP uptakes were nearly equal in both healthy and diseased plants, but not in the other three NP sources.
Significant interaction effects on the physiological N-efficiency (peN) were revealed between the two rhizobia (R and MR). There were also significant interaction effects on P-efficiency (peP) between the two AMF (M and MR) treatments. For diseased plants, the peN was significantly higher in a single R than a dual MR, but, for healthy plants, both R and MR had virtually equal peN. In pathogen-infected plants, a single M increased significantly the peP compared to healthy ones. In contrast, the result of peP with a dual MR was much higher in healthy plants than diseased ones (Table 4).
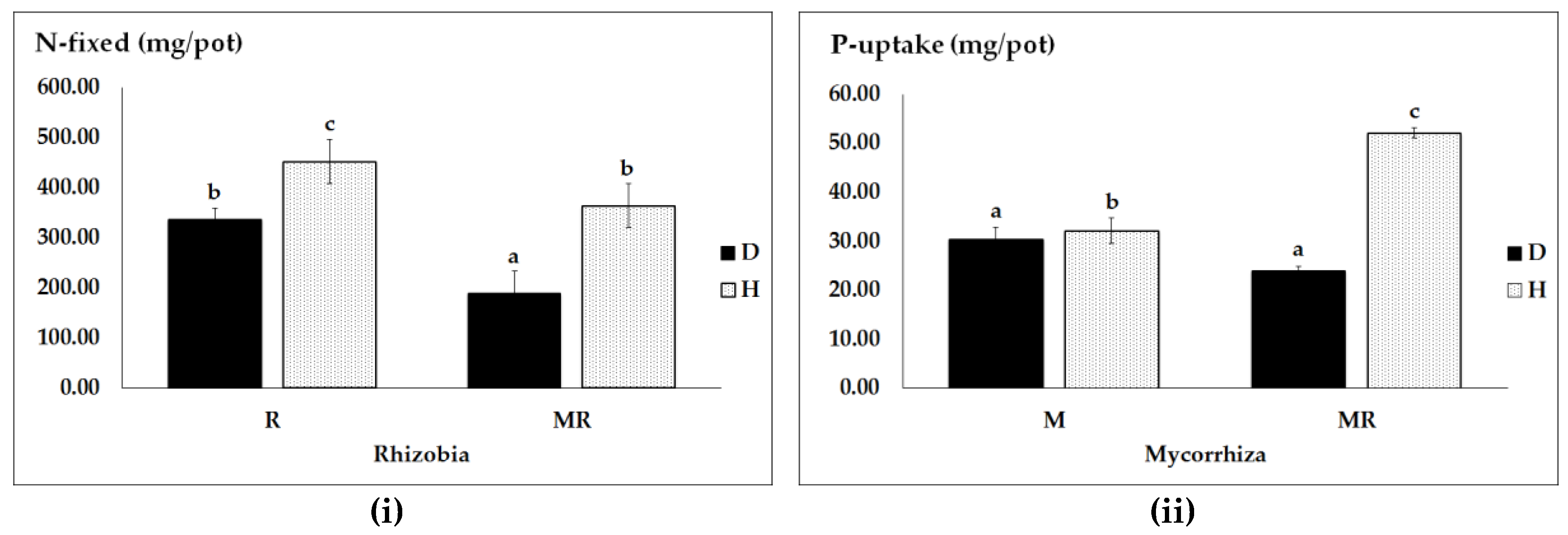
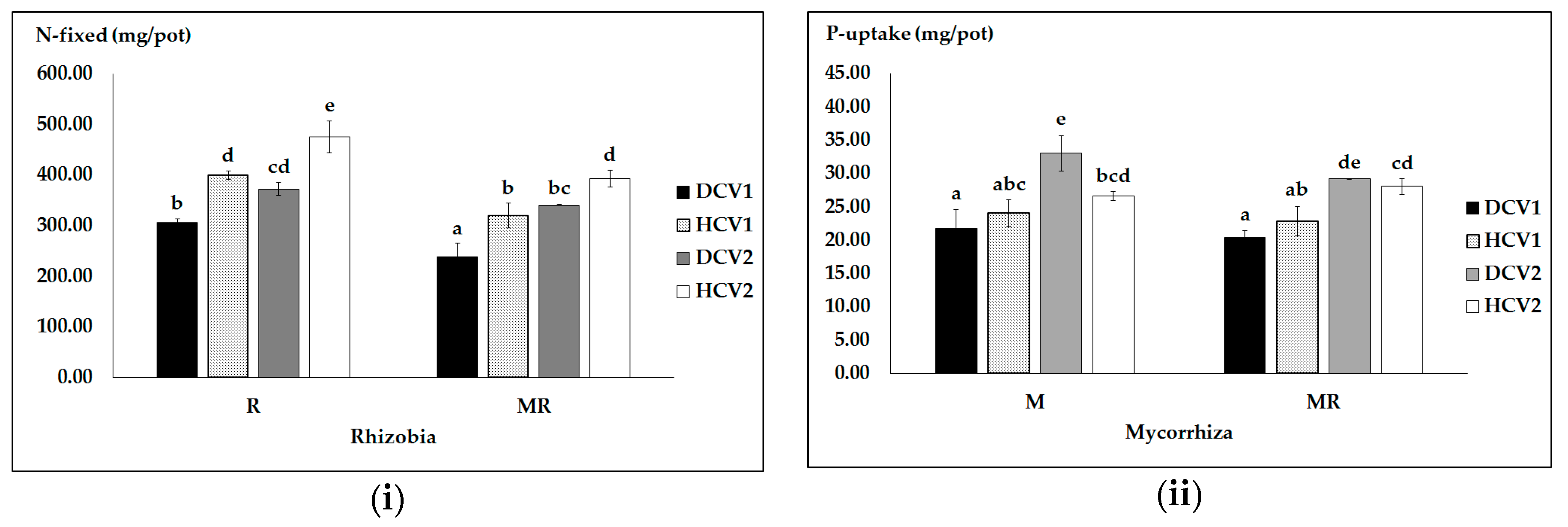
Furthermore, we found significant effects in assessments of interaction effects of pathogen X rhizobial bacteria (with or without AMF) on atmospheric nitrogen fixation as well as pathogen × AMF (with and without rhizobia) on phosphorus acquisition. In diseased pea plants inoculated with dual MR, nitrogen fixation was significantly lower than in healthy plants inoculated with either R or MR (Figure 3i). Interestingly, pathogen infection also reduced nodule DW (Figure S1). However, pathogen infected pea plants inoculated with R only had an almost equal amount of fixed nitrogen as healthy plants inoculated with a dual MR. Maximum nitrogen fixation in diseased plants was observed in pea plants inoculated with single R compared with dual MR.
In addition to nitrogen fixation, the interaction effect of pathogen × AMF (with and without rhizobia) on phosphorous acquisition was also significant. In healthy pea plants inoculated with dual MR, we found the highest P uptake (Figure 3ii). This might be due to a high demand for phosphorous to enhance the nitrogen fixation process in a dual MR treatment. Biotic stress significantly reduced fungal colonization compared to healthy plants (Figure S1). In summary, the effects of AMF on P acquisition are dependent on plant health conditions and its combination with rhizobia.
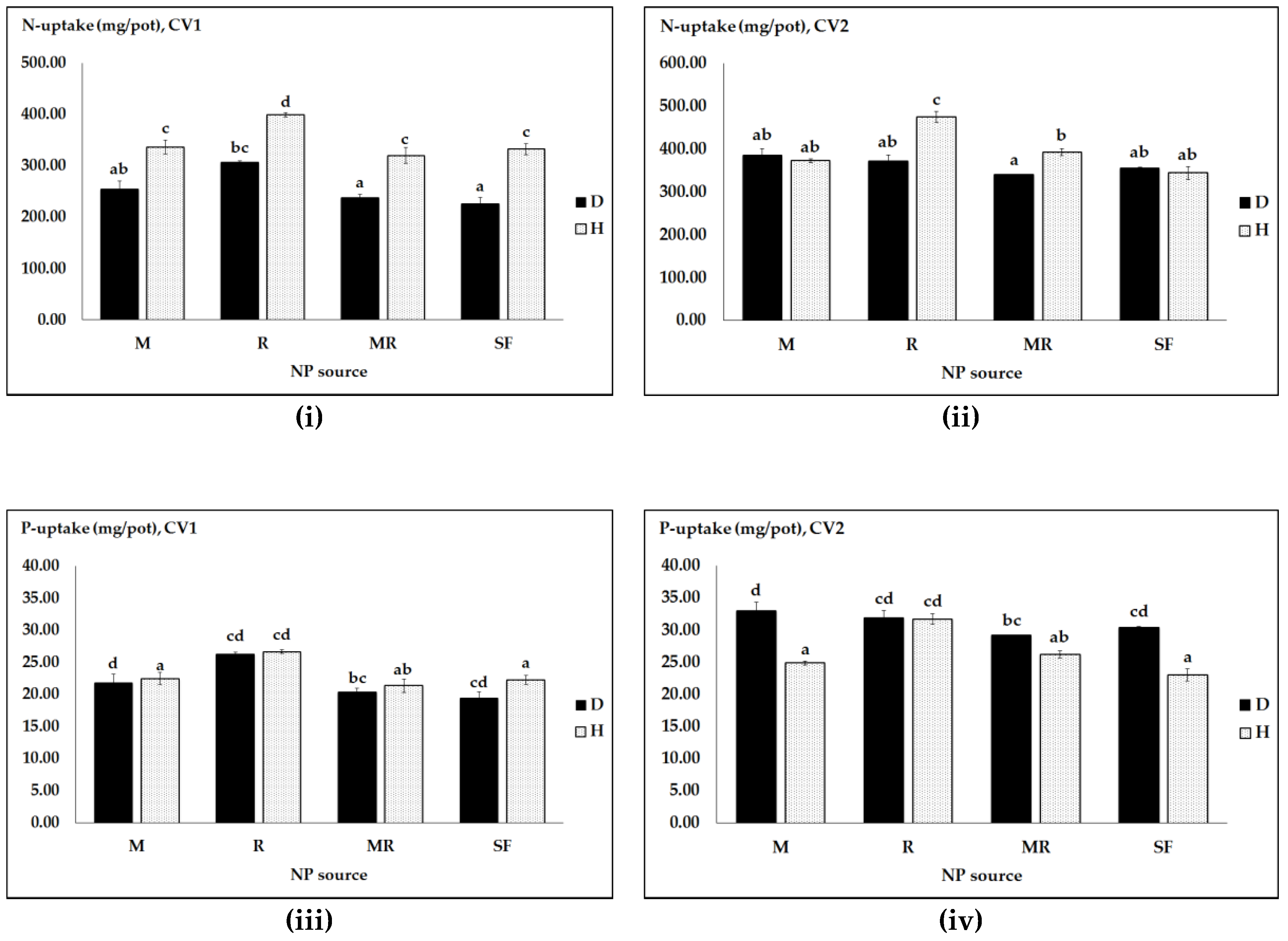
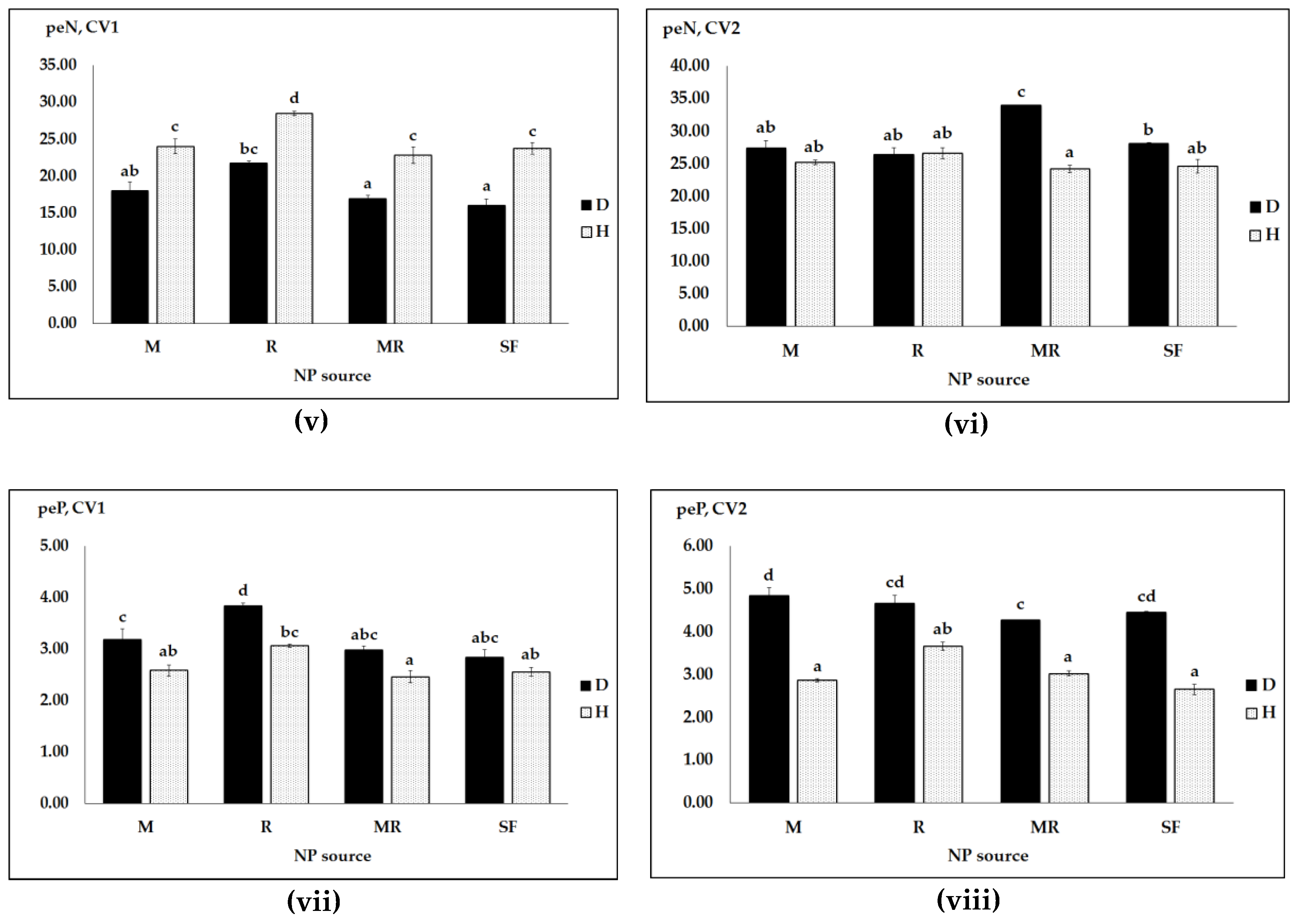
In the second experiment, the three-way ANOVA interactions were also significant for N and P uptake, and physiological N and P efficiencies (Table 5). These threefold interaction effects were largely associated with efficacy of rhizobia bacteria for N uptake and peN and of a single M inoculation for P uptake and peP in cv. Protecta (CV2) under healthy and diseased conditions. N uptake and peN were significantly higher in both healthy and pathogen infected Protecta plants inoculated with rhizobia (either R or MR) than with single M and/or SF. Additionally, both healthy and pathogen infected Protecta plants treated with single M or SF had virtually equal N uptake or peN (Figure 4ii,vi). In cv. Messire (CV1), the overall increase of N uptake and peN was much higher with single R than with single M under both healthy and pathogen-infected conditions (Figure 4i,v).
P uptake values were significantly increased with single M for pathogen infected Protecta plants compared to healthy ones (Figure 4iv). Much stronger effects of M applied alone as well as synthetic NP fertilizers on peP, especially in Protecta, were shown in pathogen infected pea plants than in healthy plants (Figure 4viii). As shown in Table 5, there were also significant differences in main effects and two-way interactions, except for the interaction of cultivar × NP regime.
For pathogen × rhizobia bacteria (with and without AMF) X cultivar on N2-fixation efficiency as well as pathogen × AMF (with and without rhizobia) × cultivar on P acquisition efficiency, no significant interactions effects were revealed. However, the second-degree interaction effects of pathogen × NP source on N2-fixation or its efficiency and on P acquisition or its efficiency were significant. The overall mean results of N2-fixation with single R were 15% higher than with dual MR, from healthy plants compared to pathogen infected ones 20% higher and from Protecta compared with Messire 25% (Figure 5i). Both cultivars showed lower nodule dry weight in pathogen infected compared to non-infected plants (Figure S2). Additionally, there were significant increases of P efficiency in pathogen infected Protecta plants inoculated with M or dual MR (Figure 5ii), but not in Messire. Further, the two pea cultivars were different in root mycorrhizal colonization (Figure S2).
In the first experiment, grain yield had significant positive correlations with pod number (0.57 **), seed number (1.00 **), straw DW (0.61 **), N uptake (0.92 **) and P uptake (0.93 **), but there was a significant negative correlation with peN (−0.55 **) and little association with peP. In the second experiment, grain yield showed significant positive correlations with N uptake (0.80 **), peN (0.96 **), and peP (0.55), but weak positive relationship with P uptake.
4. Discussion
In general, plant growth is usually co-limited by both nitrogen (N) and phosphorus (P) supply [30,31]. In addition to biotic stress, production of pea can also be co-limited by abiotic stresses such as availability of these nutrients when growing in NP deficient soils. In the tripartite mycorrhiza–legume–Rhizobium symbiotic associations in particular, the acquisitions and allocations of N and P are indeed dependent on a complex set of exchanges between rhizobia, mycorrhizal fungi and their root association [32]. Several studies have demonstrated that their inoculation improves growth and productivity of legume plants under a variety of stress conditions [33,34].
In our studies, the overall results indicated that rhizobia inoculation alone consistently increased the productivity of pea in number of pod and seed as well as straw accumulation and seed yield as compared to the AM fungi inoculated plants under both health conditions. All that might have resulted from enhanced nitrogen fixation rate, NP uptake efficiency, and photosynthetic nitrogen use efficiency under such low content of plant available nitrogen (NO3-N + NH4-N) and phosphorous in the growing media [23]. The significant effects of rhizobia, pathogen and pea interactions on nodule, disease severity and shoot biomass or N acquisition have been reported [23,35]. Previously, we also reported that the symbiotic associations of these two microbes with pea after pathogen infection could influence early plant growth components (e.g., green areas and shoot biomass production), root nodulation and mycorrhizal colonization, disease severity as well as leaf metabolome and proteome of the plant differently [23,24].
The maximum nitrogen uptake and physiological N efficiency (peN) were shown in the two rhizobial-inoculated plants compared with solely M or SF treatments. As described above, that might have been also resulted from enhanced nitrogen fixation rate, NP uptake efficiency, and photosynthetic nitrogen use efficiency under such low content of plant available NP to crop in the growing media. The overall significantly increased phosphorous abundance in pathogen infected pea grains compared to healthy ones might be driven by greater demand for the shoots of infected plants that might have subsequently led to enhanced uptake of phosphorous ion. Previously, Baldy et al. [36] reported that an increase in P availability can also lead to an increase in P uptake.
Generally, studies on seed NP uptakes and efficiency in healthy and diseased crop plants are in agreement with some previous works done, for instance, the rates of phosphates uptake by the roots of mildewed plants increased in wheat [37] and barley [38]. Rust infection in leek also increased levels of both phosphorous and nitrogen [39]. Additionally, in a healthy plant, a supply of nutrients may boost growth and, therefore, lead to relatively lower bio-mass produced per milligram of elements absorbed (dilution effect) [36]. Another explanation might be leaf proteins and in particular photosynthetic proteins of plastids are extensively degraded during senescence after D. pinodes attack, providing an enormous source of nitrogen and phosphorous that plants can tap to supplement the nutrition of growing organs such as new leaves and seeds. In Arabidopsis thaliana (L.) Heynh and oilseed rape, it has been shown that nitrogen can be remobilized from senescing leaves to expanding leaves at the vegetative stage as well as from senescing leaves to seeds at the reproductive stage [40,41,42].
Likewise, grain N in wheat will come largely from remobilization from senescing canopy tissues [43] and P resorption was more efficient in infected Citrus limon plants than in healthy plants [44]. AMF root colonization can also promote plant growth due to improved uptake of nutrients [8]. Another explanation why NP concentrations, uptakes and efficiencies are higher in grains of susceptible cultivar or diseased plants can be due to the downward disease progress on the stem destroying the phloem which translocates sugars made by photosynthetic areas of plants to the roots. We indicated cultivar specific allocations of leaf metabolites in response to D. pinodes in a previous analysis [24].
Furthermore, the contribution of leaf N remobilization to rice, wheat or maize grain N content was previously shown to be cultivar dependent, varying from 50% to 90% [45]. Furthermore, we found evidence for the reallocation or accumulation of proteins and metabolites in seeds of the two pea cultivars during pathogen defense in our previous study [46].
The present study suggests that inoculation of symbionts, especially rhizobia, should be applied to enhance resistance or tolerance against disease infection and to improve seedling growth and final grain yield of field pea. Application of the two microbial symbionts are environmentally clean and safe [33,47], while the use of synthetic NP fertilizers has important environmental and sanitary shortcomings.
5. Conclusions
This study confirms differences among various NP regimes and their interactions in N and P availability together with their effects on plant productivity, NP uptakes and their physiological efficiency with and without pathogen infection conditions. Pathogen infection enhanced mycorrhizal P-supply in Protecta more than in Messire. Plants inoculated with only rhizobia showed much better overall performance. In both cultivars, with and without pathogen infection, however, a dual microbial (MR) inoculation failed to show a consistent synergetic effect on field pea grain yield or its yield components as well as NP uptakes and their physiological efficiency. The results obtained from this study could lead to reduced or replaced N and P mineral fertilizer requirements in field pea production. This can be particularly vital for sustainable or environmentally friendly farming practices as well as inoculant producers. We further suggest a study with an infection time series and field trials to better understand the effects of symbiotic soil microbes in interaction with D. pinodes on growth, development, nutrient acquisition, final yield and grain quality of pea genotypes.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2073-4395/9/2/52/s1, Figure S1: Biotic, Figure S2: Genotype × Symbiont. Pathogen influence on mycorrhizal root colonization and nodulation in a single (Figure S1) and two (Figure S2) pea genotypes.
Author Contributions
Conceptualization, D.G. and H.-P.K.; methodology and investigation, D.G., H.-P.K. and R.T.; data organization and formal analysis, D.G; writing—original draft preparation, D.G. and H.-P.K.; writing—review and editing, D.G., H.-P.K. and S.W.; and supervision, H.-P.K. and S.W. All authors read and approved the manuscript.
Funding
The authors gratefully acknowledge support of this work by the Austrian Science Fund (FWF) with grant number P24870-B22.
Acknowledgments
We fully appreciate the two partner universities (BOKU and UNIVIE) for their administrative, technical or material support and use of their facilities. Thanks to Craig Jackson for proofreading the article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Xue, A.G.; Warkentin, T.D.; Kenaschuk, E.O. Effects of timings of inoculation with Mycosphaerella pinodes on yield and seed infection of field pea. Can. J. Plant Sci. 1997, 77, 685–689. [Google Scholar] [CrossRef]
- Tivoli, B.; Banniza, S. Comparison of the epidemiology of ascochyta blights on grain legumes. Eur. J. Plant Pathol. 2007, 119, 59–76. [Google Scholar] [CrossRef]
- Crozat, Y.; Aveline, A.; Coste, F.; Gillet, J.P.; Domenach, A.M. Yield performance and seed production pattern of field-grown pea and soybean in relation to N nutrition. Eur. J. Agron. 1994, 3, 135–144. [Google Scholar] [CrossRef]
- Jensen, E.S. Seasonal patterns of growth and nitrogen fixation in field-grown pea. Plant Soil. 1987, 101, 29–37. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D. Growth and carbon economy of arbuscular mycorrhizal symbionts. In Mycorrhizal Symbiosis; Elsevier: Amsterdam, The Netherlands, 2008; pp. 117–144. [Google Scholar]
- Selosse, M.-A.; Richard, F.; He, X.; Simard, S.W. Mycorrhizal networks: Des liaisons dangereuses? Trends Ecol. Evol. 2006, 21, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Chandel, S. Arbuscular mycorrhizal networks: Process and functions. A review. Agron. Sustain. Dev. 2010, 30, 581–599. [Google Scholar] [CrossRef]
- Farzaneh, M.; Vierheilig, H.; Lössl, A.; Kaul, H.P. Arbuscular mycorrhiza enhances nutrient uptake in chickpea. Plant Soil Environ. 2011, 57, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Aryal, U.K.; Xu, H.L.; Fujita, M. Rhizobia and AM fungal inoculation improve growth and nutrient uptake of bean plants under organic fertilization. J. Sustain. Agric. 2003, 21, 29–41. [Google Scholar] [CrossRef]
- Harris, D.; Pacovsky, R.S.; Paul, E.A. Carbon economy of soybean-Rhizobium-Glomus associations. New Phytol. 1985, 101, 427–440. [Google Scholar] [CrossRef]
- Kawai, Y.; Yamamoto, Y. Increase in the formation and nitrogen fixation of soybean nodules by vesicular-arbuscular mycorrhiza. Plant Cell Physiol. 1986, 27, 399–405. [Google Scholar]
- Mortimer, P.E.; Perez-Fernandez, M.A.; Valentine, A.J. The role of arbuscular mycorrhizal colonization in the carbon and nutrient economy of the tripartite symbiosis with nodulated Phaseolus vulgaris. Soil Biol. Biochem. 2008, 40, 1019–1027. [Google Scholar] [CrossRef]
- Paul, E.A.; Kucey, R.M.N. Carbon flow in plant microbial associations. Science 1981, 213, 473–474. [Google Scholar] [CrossRef] [PubMed]
- Redecker, D.; von Berswordt-Wallrabe, P.; Beck, D.P.; Werner, D. Influence of inoculation with arbuscular mycorrhizal fungi on stable isotopes of nitrogen in Phaseolus vulgaris. Biol. Fertil. Soils 1997, 24, 344–346. [Google Scholar] [CrossRef]
- Antunes, P.M.; Deaville, D.; Goss, M.J. Effect of two AMF life strategies on the tripartite symbiosis with Bradyrhizobium japonicum and soybean. Mycorrhiza 2006, 16, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Ide Franzini, V.; Azcón, R.; Latanze Mendes, F.; Aroca, R. Interactions between Glomus species and Rhizobium strains affect the nutritional physiology of drought-stressed legume hosts. J. Plant Physiol. 2010, 167, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Scheublin, T.R.; van der Heijden, M.G.A. Arbuscular mycorrhizal fungi colonize nonfixing root nodules of several legume species. New Phytol. 2006, 172, 732–738. [Google Scholar] [CrossRef] [Green Version]
- Angelopoulou, D.J.; Naska, E.J.; Paplomatas, E.J.; Tjamos, S.E. Biological control agents (BCAs) of Verticillium wilt: Influence of application rates and delivery method on plant protection, triggering of host defence mechanisms and rhizosphere populations of BCAs. Plant Pathol. 2014, 63, 1062–1069. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Prakash, A.; Johri, B.N. Induced systemic resistance (ISR) in plants: Mechanism of action. Indian J. Microbiol. 2007, 47, 289–297. [Google Scholar] [CrossRef]
- Choudhary, K.K.; Agrawal, S.B. Ultraviolet-B induced changes in morphological, physiological and biochemical parameters of two cultivars of pea (Pisum sativum L.). Ecotoxicol. Environ. Saf. 2014, 100, 178–187. [Google Scholar] [CrossRef]
- Jacott, C.N.; Murray, J.D.; Ridout, C.J. Trade-offs in arbuscular mycorrhizal symbiosis: Disease resistance, growth responses and perspectives for crop breeding. Agronormy 2017, 7, 75. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Van Pelt, J.A.; Verhagen, B.W.M.; Ton, J.; Van Wees, S.C.M.; Leon-Kloosterziel, K.M.; Van Loon, L.C. Induced systemic resistance by plant growth-promoting rhizobacteria. Symbiosis 2003, 35, 39–54. [Google Scholar]
- Desalegn, G.; Turetschek, R.; Kaul, H.P.; Wienkoop, S. Microbial symbionts affect Pisum sativum proteome and metabolome under Didymella pinodes infection. J. Proteom. 2016, 143, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Turetschek, R.; Desalegn, G.; Epple, T.; Kaul, H.P.; Wienkoop, S. Key metabolic traits of Pisum sativum maintain cell vitality during Didymella pinodes infection: Cultivar resistance and the microsymbionts’ influence. J. Proteom. 2017, 169, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Stacey, G.; Zhang, X.C.; Freire, M.C.M.; Le, M.H.; Oliveira, L.O.D.; Pitkin, J.W.; Segers, G.; Concibido, V.C.; Baley, G.J.; Hartman, G.L.; Upchurch, G.; Pedley, K.F. Genetic diversity and origins of Phakopsora pachyrhizi isolates in the United States. Asian J. Plant Pathol. 2012, 6, 52–65. [Google Scholar] [CrossRef]
- Hoffmann, D.; Vierheilig, H.; Peneder, S.; Schausberger, P. Mycorrhiza modulates aboveground tri-trophic interactions to the fitness benefit of its host plant. Ecol. Entomol. 2011, 36, 574–581. [Google Scholar] [CrossRef]
- Broughton, W.J.; Dilworth, M.J. Handbook for rhizobia: Methods in legume-Rhizobium technology. In The Quarterly Review of Biology; Somasegaran, P., Hoben, H.J., Eds.; University of Chicago Press: Chicago, IL, USA, 1995; pp. 224–225. [Google Scholar]
- Davidson, J.A.; Krysinska-Kaczmarek, M.; Herdina; McKay, A.; Scott, E.S. Comparison of cultural growth and in planta quantification of Didymella pinodes, Phoma koolunga and Phoma medicaginis var. pinodella, causal agents of ascochyta blight on field pea (Pisum sativum). Mycologia 2012, 104, 93–101. [Google Scholar] [PubMed]
- Delogu, G.; Cattivelli, L.; Pecchioni, N.; De Falcis, D.; Maggiore, T.; Stanca, A.M. Uptake and agronomic efficiency of nitrogen in winter barley and winter wheat. Eur. J. Agron. 1998, 9, 11–20. [Google Scholar] [CrossRef]
- Jia, Y.; Gray, V.M. Interrelationships between nitrogen supply and photosynthetic parameters in Vicia faba L. Photosynthetica 2003, 41, 605–610. [Google Scholar] [CrossRef]
- Jia, Y.; Gray, V.M.; Straker, C.J. The influence of Rhizobium and arbuscular mycorrhizal fungi on nitrogen and phosphorus accumulation by Vicia faba. Ann. Bot. 2004, 94, 251–258. [Google Scholar] [CrossRef]
- Gray, V.M. The Role of the C:N:P Stoichiometry in the Carbon Balance Dynamics of the Legume–AMF–Rhizobium Tripartite Symbiotic Association. In Plant Growth and Health Promoting Bacteria; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Kumar, B.S.D.; Berggren, I.; Martensson, A.M. Potential for improving pea production by co-inoculation with fluorescent Pseudomonas and Rhizobium. Plant Soil 2001, 229, 25–34. [Google Scholar] [CrossRef]
- Larrainzar, E.; Wienkoop, S. A proteomic view on the role of legume symbiotic interactions. Front. Plant Sci. 2017, 8, 1267. [Google Scholar] [CrossRef] [PubMed]
- Bourion, V.; Laguerre, G.; Depret, G.; Voisin, A.S.; Salon, C.; Duc, G. Genetic variability in nodulation and root growth affects nitrogen fixation and accumulation in pea. Ann. Bot. 2007, 100, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Baldy, V.; Thiebaut, G.; Fernandez, C.; Sagova-Mareckova, M.; Korboulewsky, N.; Monnier, Y.; Perez, T.; Tremolieres, M. Experimental assessment of the water quality influence on the phosphorus uptake of an invasive aquatic plant: Biological responses throughout its phenological stage. PLoS ONE 2015, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.D. Effects of fungal pathogens on nitrogen phosphorus and sulphur relations of individual plants and populations. In Nitrogen, Phosphorus and Sulphur Utilisation by Fungi: Symposium of the British Mycological Society; Boddy, L., Marchant, R., Read, D.J., Eds.; Cambridge University Press: Cambridge, UK, 1989; pp. 155–179. [Google Scholar]
- Walters, D.R.; Ayres, P.G. Phosphate uptake and translocation by roots of powdery mildew infected barley. Physiol. Plant Pathol. 1981, 18, 195–205. [Google Scholar] [CrossRef]
- Roberts, A.M.; Walters, D.R. Shoot: Root interrelationships in leeks infected with the rust, Puccinia allii Rud.: Growth and nutrient relations. New Phytol. 1989, 111, 223–228. [Google Scholar] [CrossRef]
- Diaz, C.; Purdy, S.; Christ, A.; Morot-Gaudry, J.F.; Wingler, A.; Masclaux-Daubresse, C.L. Characterization of markers to determine the extent and variability of leaf senescence in Arabidopsis. A metabolic profiling approach. Plant Physiol. 2005, 138, 898–908. [Google Scholar] [CrossRef] [PubMed]
- Malagoli, P. Dynamics of nitrogen uptake and mobilization in field-grown winter oilseed rape (Brassica napus) from stem extension to harvest: I. Global N flows between vegetative and reproductive tissues in relation to leaf fall and their residual N. Ann. Bot. 2005, 95, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Musse, M.; De Franceschi, L.; Cambert, M.; Sorin, C.; Le Caherec, F.; Burel, A.; Bouchereau, A.; Mariette, F.; Leport, L. Structural changes in senescing oilseed rape leaves at tissue and subcellular levels monitored by nuclear magnetic resonance relaxometry through water status. Plant Physiol. 2013, 163, 392–406. [Google Scholar] [CrossRef] [PubMed]
- Barraclough, P.B.; Lopez-Bellido, R.; Hawkesford, M.J. Genotypic variation in the uptake, partitioning and remobilisation of nitrogen during grain-filling in wheat. Field Crop. Res. 2014, 156, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Cheng, C.; Yang, J.; Wang, Q. Pathogen infection drives patterns of nutrient resorption in citrus plants. Sci. Rep. 2015, 5, 14675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masclaux, C.; Quillere, I.; Gallais, A.; Hirel, B. The challenge of remobilisation in plant nitrogen economy. A survey of physio-agronomic and molecular approaches. Ann. Appl. Biol. 2001, 138, 69–81. [Google Scholar] [CrossRef]
- Sistani, N.R.; Kaul, H.P.; Desalegn, G.; Wienkoop, S. Rhizobium impacts on seed productivity, quality, and protection of Pisum sativum upon disease stress caused by didymella pinodes: Phenotypic, proteomic, and metabolomic traits. Front. Plant Sci. 2017, 8, 1961. [Google Scholar] [CrossRef] [PubMed]
- Habte, M.; Osorio, N.W. Arbuscular Mycorrhizas: Producing and Applying Arbuscular Mycorrhizal Inoculum. Honolulu (HI); University of Hawaii: Honolulu, HI, USA, 2001; p. 47. [Google Scholar]
Figure 1.
Pea productivity as affected by pathogen infection (D, diseased and H, healthy) and different NP regimes (M, R, MR and SF, synthetic NP fertilizers): pod number (i); seed number (ii); straw DW (iii); and grain DW (iv). Vertical bars represent SE (n = 4) of the means. Data with the same lower case letters are not significantly different (p ≤ 0.05) from each other.
Figure 1.
Pea productivity as affected by pathogen infection (D, diseased and H, healthy) and different NP regimes (M, R, MR and SF, synthetic NP fertilizers): pod number (i); seed number (ii); straw DW (iii); and grain DW (iv). Vertical bars represent SE (n = 4) of the means. Data with the same lower case letters are not significantly different (p ≤ 0.05) from each other.

Figure 2.
Crop productivity as affected by pathogen infection (D, diseased and H, healthy) pea cultivars (CV1, Messire and CV2, Protecta) and different NP sources (M, R, MR and SF): pod number (i,ii); seed number (iii,iv); straw DW (v,vi); and grain DW (vii,viii). Vertical bars represent SE (n = 4) of the means. Data with the same lower case letters are not significantly different (p ≤ 0.05) from each other.
Figure 2.
Crop productivity as affected by pathogen infection (D, diseased and H, healthy) pea cultivars (CV1, Messire and CV2, Protecta) and different NP sources (M, R, MR and SF): pod number (i,ii); seed number (iii,iv); straw DW (v,vi); and grain DW (vii,viii). Vertical bars represent SE (n = 4) of the means. Data with the same lower case letters are not significantly different (p ≤ 0.05) from each other.


Figure 3.
Nitrogen fixation (i); and P uptake (ii) as influenced by pathogen infection (D, diseased and H, healthy) and microbial symbionts (R, M and their combination MR). Vertical bars represent SE (n = 4) of the means. Different lower case letters indicate significant differences assessed by Tukey HSD test (p ≤ 0.05) after performing a one-way ANOVA.
Figure 3.
Nitrogen fixation (i); and P uptake (ii) as influenced by pathogen infection (D, diseased and H, healthy) and microbial symbionts (R, M and their combination MR). Vertical bars represent SE (n = 4) of the means. Different lower case letters indicate significant differences assessed by Tukey HSD test (p ≤ 0.05) after performing a one-way ANOVA.

Figure 4.
NP uptakes and physiological efficiencies (pe) as affected by pathogen infection (D, diseased and H, healthy), pea cultivars (CV1, Messire and CV2, Protecta) and different NP sources: N uptake (i,ii); P uptake (iii,iv); peN (v,vi); and peP (vii,viii). Vertical bars represent SE (n = 4) of the means. Data with the same lower case letters are not significantly different (p ≤ 0.05) from each other.
Figure 4.
NP uptakes and physiological efficiencies (pe) as affected by pathogen infection (D, diseased and H, healthy), pea cultivars (CV1, Messire and CV2, Protecta) and different NP sources: N uptake (i,ii); P uptake (iii,iv); peN (v,vi); and peP (vii,viii). Vertical bars represent SE (n = 4) of the means. Data with the same lower case letters are not significantly different (p ≤ 0.05) from each other.


Figure 5.
Nitrogen fixation (i); and P uptake (ii) as influenced by pathogen infection (D, diseased, and H, healthy), microbial symbionts (R, M and their combination MR) and pea cultivars (CV1, Messire and CV2, Protecta). Vertical bars represent SE (n = 4) of the means. Different lower case letters indicate significant differences assessed by Tukey HSD test (p ≤ 0.05) after performing a one-way ANOVA.
Figure 5.
Nitrogen fixation (i); and P uptake (ii) as influenced by pathogen infection (D, diseased, and H, healthy), microbial symbionts (R, M and their combination MR) and pea cultivars (CV1, Messire and CV2, Protecta). Vertical bars represent SE (n = 4) of the means. Different lower case letters indicate significant differences assessed by Tukey HSD test (p ≤ 0.05) after performing a one-way ANOVA.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Two-way ANOVA of pea productivity (F-values) in the first experiment with only pea cultivar Messire.
Table 1.
Two-way ANOVA of pea productivity (F-values) in the first experiment with only pea cultivar Messire.
| Crop Productivity (F-Values) | ||||
|---|---|---|---|---|
| Sources | Pod Number | Seed Number | Straw DW | Grain DW |
| Pathogen (PH) | 9.79 ** | 163.15 *** | 88.35 *** | 163.07 *** |
| NP sources (NPS) | 4.56 * | 43.91 *** | 25.26 *** | 43.90 *** |
| PH × NPS | 0.46 ns | 19.19 *** | 5.25 ** | 19.18 *** |
Determined effects include pathogen infection (PH); NP sources (NPS); DW, dry weight per gram (g). Probability levels are presented at F-test (* p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001; ns, not significant).
Table 2.
Three-way ANOVA of pea productivity (F-values) from second experiment using two pea cultivars (Messire and Protecta).
Table 2.
Three-way ANOVA of pea productivity (F-values) from second experiment using two pea cultivars (Messire and Protecta).
| Crop Productivity (F-Values) | ||||
|---|---|---|---|---|
| Sources | Pod Number | Seed Number | Straw DW | Grain DW |
| Cultivar (CV) | 208.59 *** | 194.58 *** | 204.11 *** | 198.03 *** |
| NP sources (NPS) | 35.04 *** | 36.62 *** | 35.59 *** | 36.28 *** |
| Pathogen (PH) | 34.69 *** | 258.08 *** | 85.55 *** | 185.37 *** |
| CV X NPS | 0.68 ns | 0.67 ns | 0.67 ns | 0.67 ns |
| CV X PH | 34.77 *** | 22.04 *** | 30.21 *** | 24.81 *** |
| NPS X PH | 5.32 ** | 7.13 *** | 5.86 ** | 6.67 *** |
| CV X PH X NPS | 7.08 *** | 7.01 *** | 7.09 *** | 7.01 *** |
Fixed effects include pathogen infection (PH); pea cultivar (CV); NP sources (M, R, MR and synthetic NP fertilizers); DW, dry weight per gram (g). Probability levels are presented at F-test (* p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001; ns, not significant).
Table 3.
Two-way ANOVA of pea NP uptakes and physiological efficiencies (F-values) in the first experiment with pea cultivar Messire.
Table 3.
Two-way ANOVA of pea NP uptakes and physiological efficiencies (F-values) in the first experiment with pea cultivar Messire.
| Sources | Seed N and P Uptake and Efficiency (F-Values) | |||
|---|---|---|---|---|
| N Uptake | peN | P Uptake | peP | |
| PH | 128.98 *** | 562.14 *** | 124.30 *** | 3.3 |
| NPS | 50.22 *** | 7.50 *** | 60.45 *** | 7.92 *** |
| PH X NPS | 7.95 *** | 4.52 ** | 12.11 *** | 8.61 *** |
Determined effects include pathogen infection (PH); NP regimes. peN, physiological efficiency of nitrogen in a grain; peP, physiological efficiency of phosphorous in a grain. Probability levels are presented at F-test (* p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001; ns, not significant).
Table 4.
NP uptakes and their physiological efficiencies as affected by pathogen infection and different NP regimes in pea cultivar Messire.
Table 4.
NP uptakes and their physiological efficiencies as affected by pathogen infection and different NP regimes in pea cultivar Messire.
| Pathogen | NP Sources | Seed NP Uptake and Efficiency | |||
|---|---|---|---|---|---|
| N Uptake (mg/pot) | peN (mg/pot) | P Uptake (mg/pot) | peP (mg/pot) | ||
| Infected | M | 342.66 a | 35.36 b | 38.06 a | 3.11 abc |
| R | 486.23 bc | 47.37 c | 67.99 bc | 3.33 c | |
| MR | 338.33 a | 33.46 b | 36.16 a | 2.48 a | |
| SF | 421.19 ab | 41.04 bc | 62.32 b | 3.25 bc | |
| Healthy | M | 393.79 a | 6.89 a | 42.08 a | 2.50 ab |
| R | 652.08 d | 9.06 a | 96.84 d | 3.54 c | |
| MR | 563.83 c | 8.36 a | 82.05 cd | 3.57 c | |
| SF | 659.01 d | 9.94 a | 97.10 d | 3.50 c | |
Fixed effects include pathogen infection and NP sources (M, R, MR and SF, synthetic mineral nitrogen and phosphorus fertilization). Means followed by the same lower case letters in a column are not significantly different (p ≤ 0.05) from each other.
Table 5.
Three-way ANOVA of NP uptakes and their physiological efficiencies (F-values) from second experiment using two pea cultivars (Messire and Protecta).
Table 5.
Three-way ANOVA of NP uptakes and their physiological efficiencies (F-values) from second experiment using two pea cultivars (Messire and Protecta).
| Sources | Seed N and P Uptake and Efficiency (F-Values) | |||
|---|---|---|---|---|
| N Uptake | peN | P Uptake | peP | |
| Cultivar (CV) | 201.62 *** | 201.29 *** | 220.18 *** | 231.98 *** |
| NP Sources (NPS) | 35.92 *** | 35.98 *** | 33.15 *** | 29.88 *** |
| Pathogen (PH) | 126.26 *** | 133.82 *** | 16.22 *** | 326.76 *** |
| CV X NPS | 0.66 ns | 0.66 ns | 0.76 ns | 1.00 ns |
| CV X PH | 27.75 *** | 27.36 *** | 49.27 *** | 72.49 *** |
| NPS X PH | 6.21 *** | 6.29 *** | 3.94 * | 2.70 ns |
| CV X PH X NPS | 7.06 *** | 7.06 *** | 7.05 *** | 6.85 *** |
Fixed effects include pathogen infection (PH); pea cultivar (CV); NP sources (M, R, MR and synthetic NP fertilizers). The abbreviations of peN and peP stand for physiological efficiencies (pe) of N and P, respectively. Probability levels are presented at F-test (* p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001; ns, not significant).
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Desalegn, G.; Turetschek, R.; Wienkoop, S.; Kaul, H.-P. Didymella pinodes Affects N and P Uptakes and Their Efficiencies in a Tripartite Mutualism of Pea. Agronomy 2019, 9, 52. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9020052
AMA Style
Desalegn G, Turetschek R, Wienkoop S, Kaul H-P. Didymella pinodes Affects N and P Uptakes and Their Efficiencies in a Tripartite Mutualism of Pea. Agronomy. 2019; 9(2):52. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9020052
Chicago/Turabian StyleDesalegn, Getinet, Reinhard Turetschek, Stefanie Wienkoop, and Hans-Peter Kaul. 2019. "Didymella pinodes Affects N and P Uptakes and Their Efficiencies in a Tripartite Mutualism of Pea" Agronomy 9, no. 2: 52. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9020052
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.