Grassland Management Influences the Response of Soil Respiration to Drought
by
 , ,
, ,
Gabriel Y. K. Moinet
1,* ,
,
Andrew J. Midwood
2,
John E. Hunt
1,
Cornelia Rumpel
3,4,
Peter Millard
1 and
Abad Chabbi
3,4,5 1
Manaaki Whenua–Landcare Research, PO Box 69040, Lincoln 7640, New Zealand
2
Department of Biology, University of British Columbia–Okanagan, Kelowna, BC V1V 1V7, Canada
3
The French National Center for Scientific Research (CNRS), Institut d’Ecologie et des Sciences de l’Environnement Paris (IEES), UMR 7618, Batiment EGER, Aile B, F-78850 Thiverval Grignon, France
4
AgroParisTech, French Natl Inst Agr Res INRA, UMR ECOSYS, F-78850 Thiverval Grignon, France
5
National Institute of Agricultural Research (INRA), Centre de recherché, Nouvelle-Aquitaine-Poitiers, URP3F, 86600 Lusignan, France
*
Author to whom correspondence should be addressed.
Agronomy 2019, 9(3), 124; https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030124
Submission received: 24 February 2019
/
Accepted: 3 March 2019
/
Published: 7 March 2019
(This article belongs to the Special Issue Grassland Management for Sustainable Agroecosystems)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Increasing soil carbon stocks in agricultural grasslands has a strong potential to mitigate climate change. However, large uncertainties around the drivers of soil respiration hinder our ability to identify management practices that enhance soil carbon sequestration. In a context where more intense and prolonged droughts are predicted in many regions, it is critical to understand how different management practices will temper drought-induced carbon losses through soil respiration. In this study, we compared the impact of changing soil volumetric water content during a drought on soil respiration in permanent grasslands managed either as grazed by dairy cows or as a mowing regime. Across treatments, root biomass explained 43% of the variability in soil respiration (p < 0.0001). Moreover, analysis of the isotopic composition of CO2 emitted from the soil, roots, and root-free soil suggested that the autotrophic component largely dominated soil respiration. Soil respiration was positively correlated with soil water content (p = 0.03) only for the grazed treatment. Our results suggest that the effect of soil water content on soil respiration was attributable mainly to an effect on root and rhizosphere activity in the grazed treatment. We conclude that farm management practices can alter the relationship between soil respiration and soil water content.
1. Introduction
Grasslands cover 26% of Earth’s ice-free land surface area, representing 70% of the world’s agricultural area [1] and containing 20% of global soil carbon stocks [2]. There is now a wide interest in improving carbon storage on land as a negative emission technology to stay below the 1.5 °C global warming limit [3]. Increasing soil carbon stocks in grasslands has a strong potential to contribute to this effort [4,5]. However, the impacts of different grassland management practices are not well understood [6], and recent reviews highlight a lack of data to clarify the mechanisms by which various management practices affect soil organic carbon (SOC) stocks [7,8]. Moreover, there are large uncertainties concerning the drivers of soil respiration (Rs), which is the second largest terrestrial carbon flux globally [9].
Global climate change is predicted to lead to an increase in the intensity and frequency of extreme events in large areas of the world. Droughts may cause intense water stress for plants and soil organisms [10]. In other areas, increasing rainfall together with evapotranspiration and changes in precipitation patterns could potentially lead to constrained water availability during the growing season [11]. In a context where the global water cycle is predicted to change, understanding the effect of soil water content on soil respiration is critical.
Soil respiration is the result of two processes: root and rhizosphere respiration (autotrophic component), and microbial decomposition of soil organic matter (heterotrophic component). Both plants [12] and micro-organisms [13] are sensitive to changes in soil water availability, resulting in soil respiration being sensitive to both soil water content [14,15] and the frequency and intensity of precipitation [16]. However, our understanding of the effect of drought on soil respiration remains relatively limited, notably due to differential responses of the autotrophic and heterotrophic components and the difficulties associated with quantifying them [17,18].
Usually, studies addressing the effects of soil water content on heterotrophic soil respiration are carried out in the laboratory in the absence of plants. Partitioning soil respiration into its autotrophic and heterotrophic components can be achieved with minimal soil disturbance in the field using the stable isotopes of carbon [17,18,19,20,21], although this can prove challenging in dry conditions where low CO2 efflux may be below the detection limits of instrumentation [17]. Nonetheless, even when the rate of autotrophic and heterotrophic respiration cannot be determined, the 13C isotopic signature (δ13C) of CO2 emitted from the soil (δ13CRs) can provide qualitative information on the dynamic soil respiration components. This is because the δ13C of emitted CO2 becomes more enriched as the contribution of microbially derived Soil Organic Matter (SOM) becomes an increasing part of total soil respiration [22,23].
Understanding the effect of management practices on the drought response of soil respiration and the plant and microbial processes driving it is critical to develop agricultural systems that are resistant to extreme climatic events. Here we compared the impact of changing soil water content during a severe drought on soil respiration and the δ13C of CO2 emitted from soils (δ13CRs), roots, and rhizosphere (δ13CRa), and root-free soils (δ13CRh) in a permanent pasture grazed by dairy cows and a permanent pasture under a mowing regime with no grazing mammals. These two management systems are among the main types of grassland management in Europe [24] and have a strong potential to store carbon below ground and partly offset greenhouse gases emissions [25]. The main objective was to determine whether these two different grassland management practices influence the response of soil respiration and its components to drought.
2. Materials and Methods
2.1. Site Description
The site is located at the national long-term experimental observatory Système d’Observation et d’Expérimentation pour la Recherche en Environnement-Agroecosystems, Biogeochemical Cycles and Biodiversity (SOERE-ACBB) near Lusignan, western France (46°25′12.91″ N; 0°07′29.35″ E). The soil is classified as a Dystric Cambisol [26]. It developed from loamy parent material of unknown origin over red clay [27]. The soil profile can be divided into two main domains: Upper soil horizons are characterized by a loamy texture, classified as Cambisol, whereas lower soil horizons are clayey rubefied horizons, rich in kaolinite and iron oxides, classified as a Paleo-Ferralsol [28].
The two experimental paddocks of about 3 ha in surface area were converted to permanent grasslands in 2005. Since then, one paddock was grazed by a herd of dairy cows, and the other paddock was managed by periodically mowing and harvesting the biomass. The timing for harvest and mowing are made so as to maximize above-ground production and rarely happen at the same time. Following this guideline, harvest happened 2 days before our experiment started, while grazing had happened 2 weeks before the start of the experiment.
The plant community in the mown grassland was a mixture including Dactylis glomerata L. (cocksfoot) cultivar Ludac, Festuca arundinacea Schreb (tall fescue) cultivar Soni, and Lolium perenne L. (rye-grass) cultivar Milca. In the grazed grassland, Trifolium repens L. (white clover) cultivar Menna was added to the multispecies mixture. The mown grassland was cut four times a year with biomass exported, and nitrogen (N) fertilizer was applied at rates comprised between 120 and 310 kg N ha−1 year−1. Fertilizer application rates were adjusted to maintain the nitrogen nutrition index between 0.9 and 1.0, that is, close to non-limiting nitrogen nutrition to near maximum plant production [28]. Grazing in the grazed paddock took place from March to December with 50 days per year using 15 to 20 livestock unit per hectare. Grazed grasslands did not receive nitrogen fertilization but nitrogen losses were returned by dung and urine and through the presence of leguminous species.
In 2011, six years after management conversion to permanent grasslands, bulk density was identical in the mown and grazed treatments, averaging 1.4 g cm−3 in the top 300 mm. Carbon stocks in 2014 ranged between 55 (mown grassland) and 64 (grazed grassland) t ha−1 in the first 300 mm. Nitrogen stocks were also similar in both grasslands, with 1.7 and 1.9 t ha−1 for the mown and grazed grasslands, respectively. No significant changes had occurred during the first 9 years (2014) after the conversion to permanent grasslands (A. Crème, personal communication).
2.2. Experimental Design
The experiment took place in early June 2017, at the beginning of a heat wave that affected large parts of Europe, including western France. At the beginning of a period of 10 days with no precipitation, we created a large range of soil water contents by applying a large artificial rainfall event. The responses of soil respiration (Rs) of the 13C isotopic signature of CO2 respired from the whole soil (δ13CRs), and its autotrophic (live roots and rhizosphere, δ13CRa) and heterotrophic (root and rhizosphere-free soil, δ13CRh) components to changes in soil water content were compared for the grazing and mown systems.
One month before the start of the experiment, 10 rectangular plots of 1 × 2 m were positioned at random locations in each paddock, and four PVC collars (100 mm diameter, 30 mm depth) were fully inserted at random locations within each plot. On each day of measurement, a plot was randomly selected in each paddock (mown and grazed) and a set of measurements of Rs, δ13CRs, δ13CRa, δ13CRh, soil volumetric water content (θs), and soil temperature (Ts) were taken from each collar.
A full set of measurements were taken on the four collars in one plot of each paddock before water was applied (day 0). Two hundred liters of water per plot was progressively applied over the course of 12 h to prevent run-off and pooling to the nine remaining plots in each paddock, equivalent to 100 mm of rainfall. Measurements were then made from the four collars in one randomly selected plot of each paddock every morning for the next 8 days, starting about 24 h after the end of watering to avoid the potential short-term burst of CO2 emissions (the so-called “Birch effect”) observed when dry soils are rewetted.
One plot in the grazed paddock was selected to characterize short-term changes in soil respiration due to the application of water. An additional set of four PVC collars were inserted in a 1 × 2 m area adjacent to the selected plot but received no water addition, constituting a control treatment. Measurements of Rs, θs, and Ts were taken 2 h and 1 h before water application on each of the eight collars. Water was then applied on one of the plots over the course of 1 h by spreading 100 mm evenly on the surface area using a hand watering can. Measurements of Rs, θs, and Ts were then carried out on the four watered and four dry collars every hour for 12 h.
2.3. Measurements of Soil Respiration and δ13C of Respired CO2
Except for the short-term watering experiment, where Rs was measured using a closed dynamic chamber system (EGM-4, PP systems, Amesbury, MA, USA), measurements of Rs were taken using two custom-built open chamber systems with four chambers each, adapted from Midwood et al. [29] and Midwood and Millard [30]. The chambers were placed on the collars set in the soil, and CO2-free air was supplied to the chambers using mass flow controllers (model FMA5510, Omega Engineering Ltd., Stamford, CT, USA). The air was pumped out of the chambers using diaphragm pumps (TD−3, Brailsford and Co. Inc., Antrim, NH, USA), and the flow rate was controlled by mass flow controllers and adjusted to15 mL min−1 lower than the inflow of CO2-free air to avoid any air ingress from the atmosphere. The CO2 concentration of the air leaving the chamber was measured using an infrared gas analyzer (IRGA) (Li840, LiCor Biosciences, Cambridge, UK), and the entire system was controlled by a datalogger (CR1000 and SDM-CV04, Campbell Scientific Ltd., Logan, UT, USA), allowing adjustment of the inflow of CO2-free air to the measured CO2 concentration to obtain a constant target concentration value in the chamber. After an equilibration period of 90 min and when the chamber’s CO2 concentrations were constant at 440 ppm, approximately 500 mL of respired air was collected in pre-evacuated air-tight bags (Tedlar® Keika Ventures, Chapel Hill, NC, USA) and the gas samples were analyzed for δ13C values. All gas samples were analyzed for δ13C values using a tuneable diode laser (TDL, TGA100A; Campbell Scientific Inc., Logan, UT, USA). The rate of Rs was calculated from the measured CO2 concentration in the chamber and the flow rate of the CO2-free air delivered to the chamber.
Measurements of the isotopic signatures of the CO2 respired by the heterotrophic (root- and rhizosphere-free soil, δ13CRh) and autotrophic (roots and rhizosphere, δ13CRa) components of the soil were made by adapting the technique described by Snell et al. [23]. After RS had been measured and the soil surface efflux sampled, roots and soils were collected. The collars were removed, and a soil core was extracted using a 100 mm diameter steel tube hammered into the soil to a depth of 250 mm. The soil from the core was broken up loosely and the roots removed by hand.
Shifts in δ13CRh have been shown to change exponentially with time after a soil core is extracted and broken up [19,23]. Thus, to obtain values of δ13CRh that are representative of the isotopic signature of the carbon pool used as a substrate before disturbance (time zero), one needs to work as rapidly as possible. To be as consistent as possible between samples, each operation was kept to a constant time. The process of breaking up the soil core in a tray and removing the roots by hand was kept close to 90 s. Subsequently, a subsample of root-free soil was placed in an air-tight bag, flushed quickly three times with nitrogen gas to purge atmospheric air from the soil, then filled with approximately 500 mL of CO2-free air and allowed to incubate at the ambient temperature until the time from sampling the soil core reached 3 min. The air in the bag was then sampled and analyzed for δ13CO2.
The roots were then cleaned of most of the remaining soil attached to them and placed in a separate air-tight bag. The bags were evacuated and then filled with approximately 500 mL of CO2-free air and allowed to incubate at ambient temperature for between 20 and 60 min, after which the air in the bags was sampled for measurements of δ13CO2. The roots were then dried at 105 °C for 24 h and weighted to give an estimate of the root biomass in each soil core.
2.4. Statistical Analyses
To test for the effect of soil volumetric water content (θs) on Rs, δ13CRs, δ13CRa, and δ13CRh for the paddocks managed as grazed and mown systems, a backwards stepwise regression approach was used separately on each explained variable [31]. Each measurement of Rs, δ13CRs, δ13CRa, and δ13CRh was treated as a sample. The full linear models included the three explanatory variables θs, root biomass, and paddock management (as a factor), as well as every two-way interaction between them. Model selection was based on a comparison of Akaike’s Information Criterion (AIC), the model with the lowest AIC value being the most strongly supported. As a rule of thumb, when two models presented a ΔAIC < 2, the simpler model was selected [32]. All statistical analyses were conducted using R version 3.4.2 [33] (R Development Core Team, 2017).
3. Results
3.1. Hourly Measurements: Short-Term Response of Soil Respiration to Water Addition
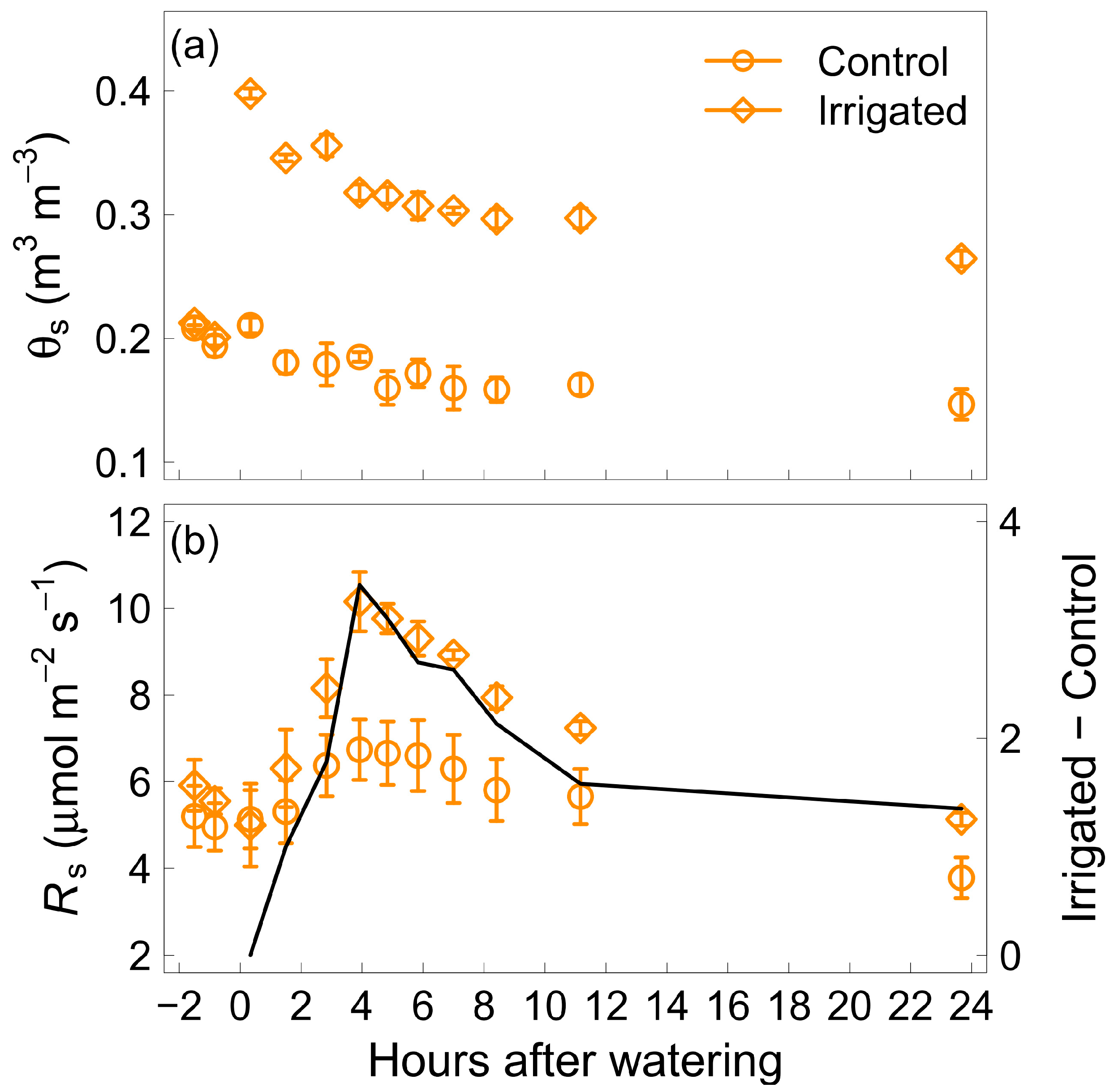
In the grazed paddock, soil volumetric water content (θs) greatly increased immediately after 100 mm of water was added (irrigated plot), approximately doubling from 0.2 to 0.4 m3 m−3 and becoming significantly higher than in the control plot without water addition. θs then slowly decreased over the next 24 h in both plots but remained significantly higher in the irrigated plot (Figure 1a), with a difference between the irrigated and the control plot of over 0.11 m3 m−3 throughout the 24 h.
Soil respiration (Rs) started to increase 2 h after water was added to the irrigated plot, becoming significantly higher than in the adjacent control plot and remaining higher during the 24 h measuring period (Figure 1b). Rs also varied in the control plot due to diel variation. Rs peaked for both plots 4 h after water was added. The difference between mean Rs in the irrigated and the control plot was null before and just after water was added. This difference started to increase 1 h after water addition, was highest 4 h after water addition with a value of 3.4 µmolCO2 m−2 s−1, and then decreased and levelled off, being similar 11 h and 24 h after water addition (Figure 1b).
3.2. Daily Changes after Water Addition
To assess the effect of water addition on Rs, δ13CRs, δ13CRa, δ13CRh, only the measurements taken from 24 h onward after water addition were included to exclude the pulse of respiration measured shortly after water addition.
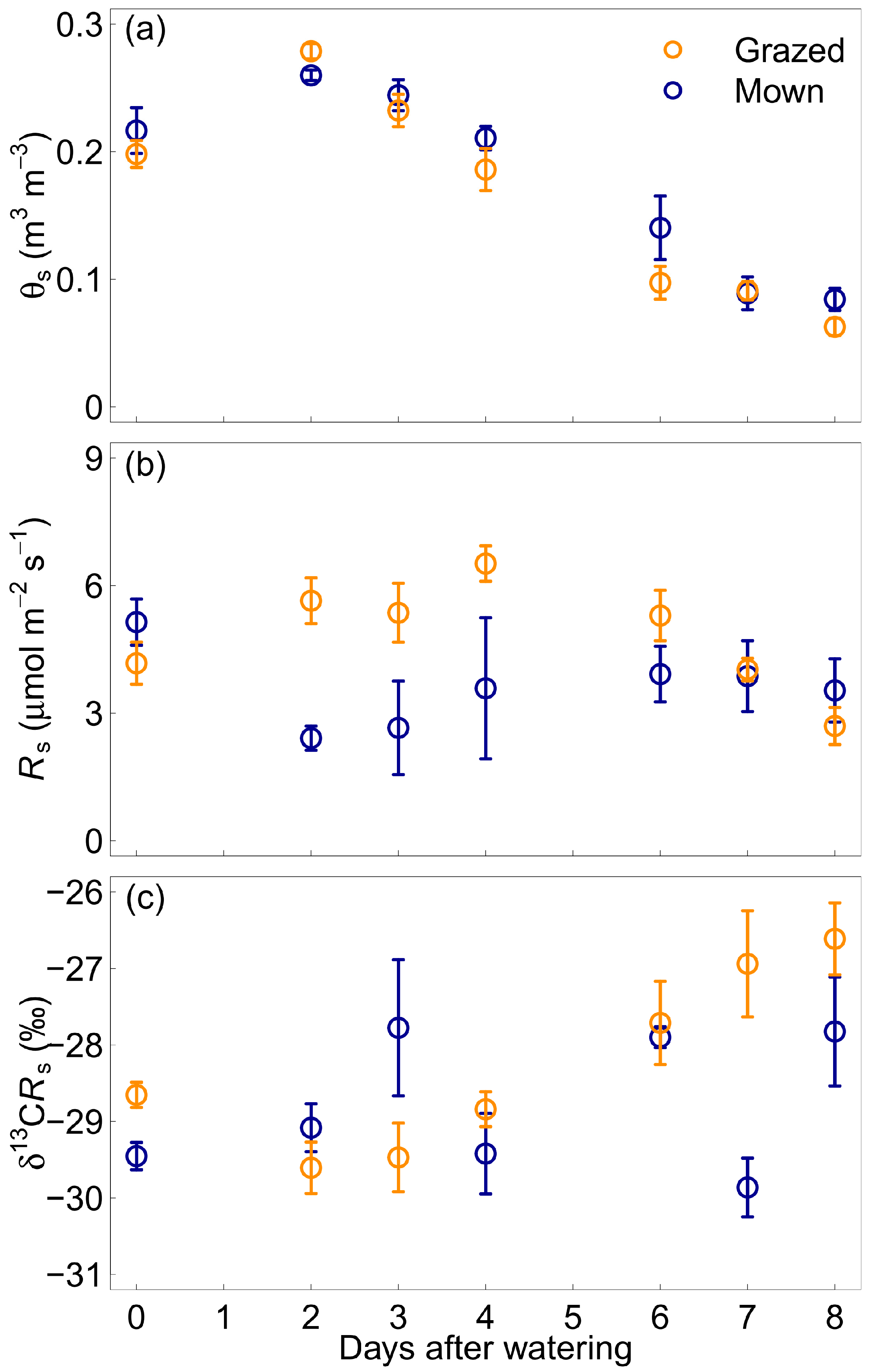
Soil volumetric water content (θS) increased after the addition of 100 mm of water on the plots and remained higher than before watering for 3 days. After the initial increase, θS decreased steadily for the whole experiment period of 8 days (Figure 2a). Individual values of θS overall ranged from 0.05 to 0.31 m3 m−3.
Although the trend for θS was identical in both paddocks, Rs showed different patterns in the grazed and mown paddocks over the days after water was added (Figure 2b). In the grazed paddock, Rs roughly followed a similar pattern to θS, showing higher values after water addition for 3 days and decreasing steadily after that. Rs did not show any obvious pattern in the mown paddock and was more variable than in the grazed paddock. Similarly, the 13C isotopic signature of soil-respired CO2 (δ13CRs) presented a different pattern for the two paddocks (Figure 2c). Changes in δ13CRs were tightly coupled with changes in θS in the grazed paddock, with more depleted values 2 days after water addition and a steady change to more enriched values over the measurement period. As for Rs, δ13CRs did not show any obvious pattern over the days after water addition in the mown paddock.
The 13C isotopic signature of CO2 respired from the roots and from the root- and rhizosphere-free soil did not show clear patterns after water addition for either paddock (Figure S1).
3.3. Effect of Soil Volumetric Water Content
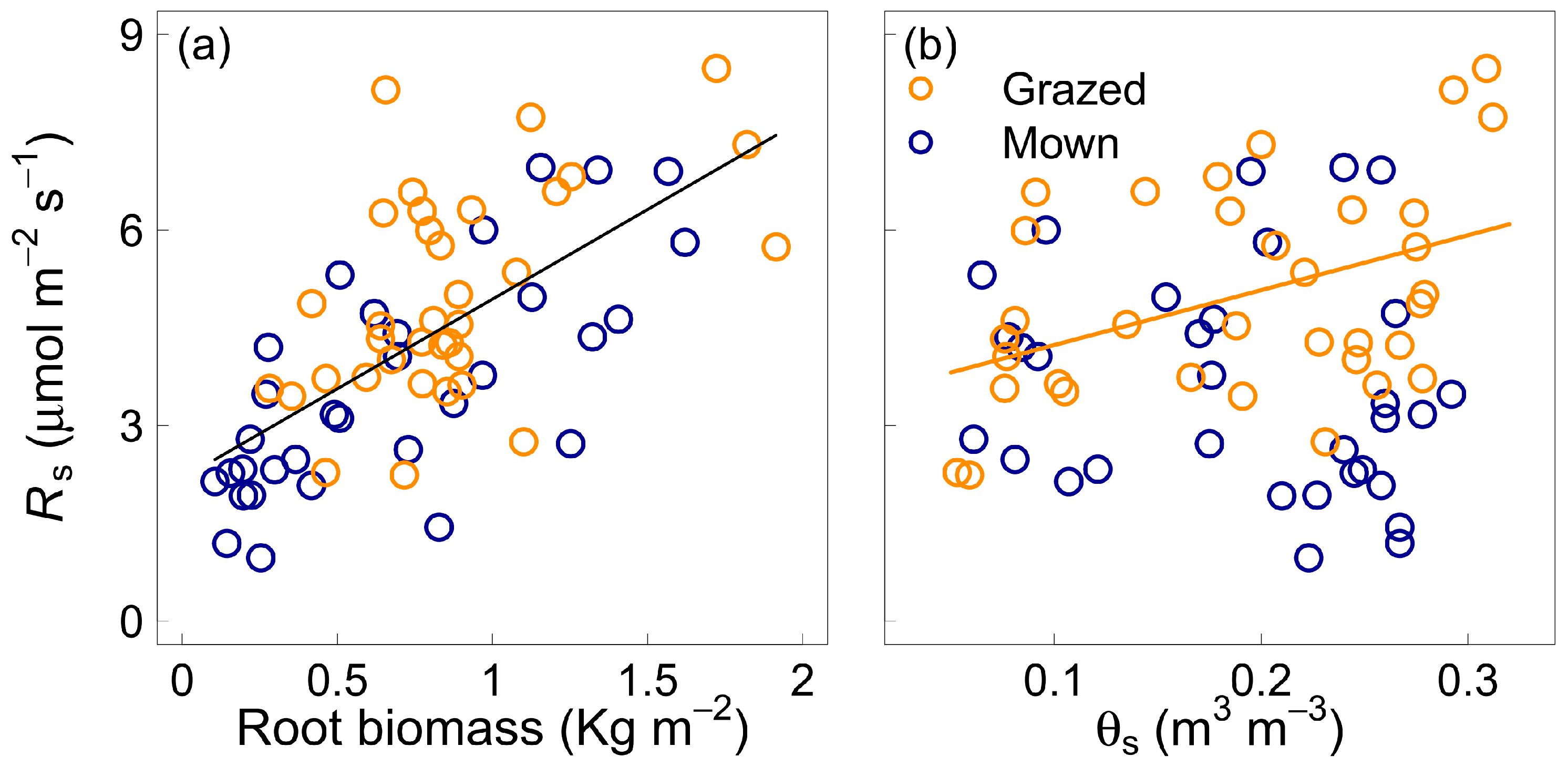
The model for Rs with the best fit selected from the backwards stepwise regression, explained 54% of the variability in Rs and incorporated the effect of root biomass (F = 55.74, p < 0.0001) and the interactive effect of soil water content and paddock management (F = 4.99, p = 0.03). The single-variate model including root biomass explained 43% of the variability in Rs (Figure 3a). Soil respiration was positively correlated with θs in the grazed paddock (p = 0.03) but was not significantly affected by θS in the mown paddock (p = 0.2, Figure 3b).
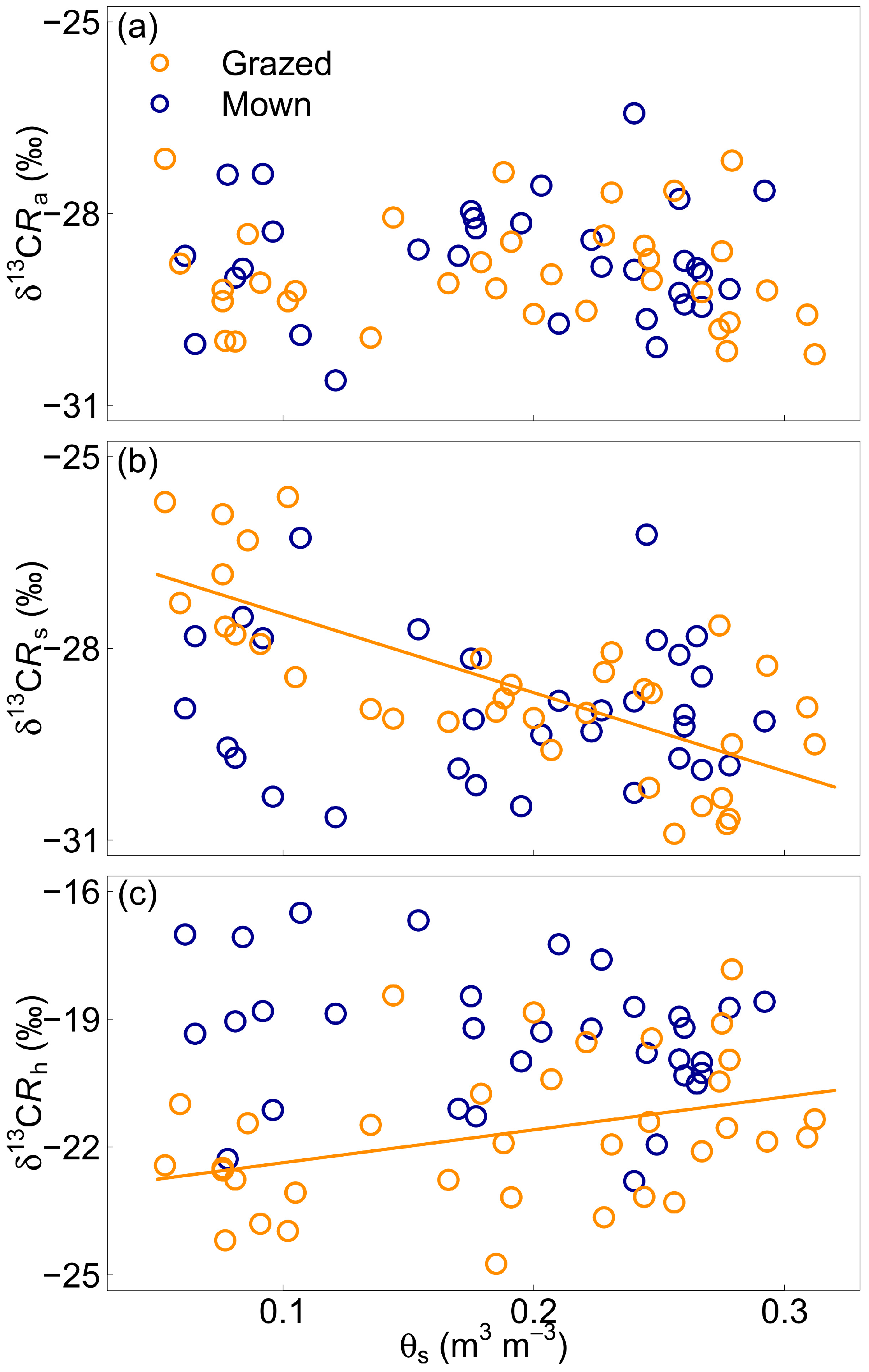
As for Rs, the model for δ13CRs with the best fit incorporated the effect of root biomass (F = 4.41, p = 0.04) and the interactive effect of soil water content and paddock management (F = 9.65, p < 0.01). This model explained 39% of the variability in δ13CRs. Soil volumetric water content had different effects for the two paddocks (Figure 4a), with a significant effect of θS on δ13CRs for the grazed paddock (p < 0.01), but no significant effect for the mown paddock (p = 0.6).
The model for δ13CRh with the best fit incorporated the interactive effect of root biomass and paddock management (F = 7.63, p < 0.01) and the interactive effect of soil water content and paddock management (F = 6.44, p = 0.01), and explained 39% of the variability in δ13CRh. The effect of root biomass and of θs on δ13CRh was significant only for the grazed paddock (p < 0.001 for root biomass and p = 0.04 for θs). The model for δ13CRa with the best fit was a single-variate model incorporating only root biomass (F = 7.23, p < 0.01) and explained 10% of the variability in δ13CRa. There was no significant effect of θs on δ13CRa or δ13CRh (Figure 4a,c).
It is noteworthy that soil temperature (Ts) ranged from 17.6 to 29.6 °C and was negatively correlated with θS (F = 33.84, p < 0.0001). Because of this correlation, the effect of Ts and θS was analyzed separately. In the model with the best fit for each backwards stepwise regression analysis, θS was replaced with Ts, and the effect was assessed by comparing the coefficient of determination (R2) for the models including θs and Ts. Replacing θS by Ts in the best models resulted in a lower coefficient of determination for all the variables.
4. Discussion
4.1. Birch Effect
It is well documented that rewetting dry soils is usually followed by a burst of respiration, which can be very short-lived [34] or last for several days [35], after which respiration rates stabilize to that of a continuously wet soil. In our study, with a flush of CO2 starting 4 h after watering and the difference between dry and wet soils coming to equilibrium over the few hours after that, the Birch effect was observed to last less than a day. Therefore, we considered measurements taken 24 h after watering for analyzing Rs and its components to exclude the influence of the Birch effect.
4.2. Soil Water Content and Isotopic Discrimination
Changes in δ13CRa and δ13CRh with soil water content were largely insignificant, in line with previous results where irrigated and non-irrigated plots were not found to differ significantly in their values of δ13CRa and to marginally affect δ13CRh [17]. This may appear surprising, particularly for the autotrophic component. Under conditions of limited water availability, plants close their stomata, improving their water use efficiency and usually leading to lower 13C discrimination [36,37]. Moreover, substrates respired in the roots are supplied mainly by recent photo-assimilates [38,39]. We would, therefore, have expected more enriched values for δ13CRa with higher water stress and lower photosynthetic 13C discrimination. Our result suggests that the substrates for respiration of the roots were not tightly coupled with photo-assimilates. Water stress may reduce the phloem diffusion rate, therefore, increasing the time lag between assimilation and utilization of compounds in the roots and rhizosphere and resulting in photosynthesis to decrease faster than respiration rates under water stress [38,39]. We conclude that carbohydrate reserves provided substrates for the autotrophic component of soil respiration and were not exhausted during the 8 days following watering.
4.3. Soil Water Content and Components of Soil Respiration
The 13C signatures of CO2 respired from the roots and from the whole soil were similar in value for high water content, ranging between −31 and −27‰. Probably due to the relatively low carbon content at this site, the contribution of SOM decomposition to total RS was small and δ13CRs was largely dominated by the signature from the roots, also in line with the observation that roots can be a dominant contributor to total Rs in many ecosystems [40,41,42]. Not surprisingly, soil respiration was strongly positively correlated with root biomass for both treatments.
Soil water content is a major driver of root respiration and can be the limiting factor for plant activity in dry conditions [43,44,45]. When the soil was wet just after water addition, the contribution of roots and rhizosphere respiration to total soil respiration was probably at its maximum and was close to 100%, leading to values of δ13CRs and δ13CRa being indistinguishable. With such small changes in the components of soil respiration (δ13CRa and δ13CRh), changes in δ13CRs must have been due to changes in the relative contribution of the autotrophic and heterotrophic components.
Because the autotrophic component dominated soil respiration, the enrichment of δ13CRs with decreasing water content in the grazed paddock was likely to be due to the decreasing contribution of the root and rhizosphere component rather than an increasing contribution of the heterotrophic component. This hypothesis is in line with the results from a laboratory study showing that microbial activity in the rhizosphere of grassland species was less affected than root activity [46]. This enrichment, therefore, suggests that in the grazed treatment, soil respiration decreased with water content due to a decreasing autotrophic component.
No changes were observed in the mown treatment. Carbohydrate reserves may vary with grass species and development stage [47]. It is possible that the plant community in the mown treatment presented a rooting system with more carbon reserves and increased resilience to drought. Another explanation could be found in the fact that grazing and mowing happened at different times. Due to a farm management decision, mowing had happened shortly before the beginning of our experiment. Grasses have been observed to mobilize their root reserves and allocate them above-ground for leaf regrowth after cutting or grazing [48,49]. It is, therefore, possible that root exudation and respiration were decreased due to mowing, thereby, cancelling the positive effect of adding water on root activity observed in the grazed treatment.
4.4. Consequences for Soil Carbon Sequestration
Agricultural management practices are known to influence the fate of soil carbon stocks, in interaction with climate and soil properties [5]. Although permanent grasslands are known to have a strong potential to store carbon below ground [24], soil carbon sequestration may be reversible under the effect of climate change and changes in management practices, particularly those involving soil disturbance [25]. Our results showed that different management practices and decisions are strong contributors to determine the response of soil carbon dynamics to soil water content. Although we were not able to conclude directly about which of mowing or grazing management will enhance drought resilience, we showed that the adaptation and resilience of agricultural grasslands to an intensified global water cycle will likely strongly depend on management practices and decisions. These decisions will, therefore, determine the magnitude of climate-induced changes in grasslands soil carbon stocks and should be the subject of careful evaluation.
5. Conclusions
Soil respiration in grazed and mown grasslands responded differently to changes in soil water content. It is possible that the unsynchronized cutting and grazing regimes partly explained this difference. Our results suggest that farm management practices and the timing of management decisions potentially alter the relationship between soil carbon dynamics and soil water content. Therefore, management choices and decisions will likely significantly contribute to driving changes in soil carbon stocks under the influence of climate change as the global water cycle changes and the intensity and frequency of droughts increase. More studies are needed to fully identify which management practices can lead to increased resilience and adaptation of grassland ecosystems to drought.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2073-4395/9/3/124/s1, Figure S1: 13C isotopic signature of CO2 respired from the roots (δ13CRa, panel a) and of CO2 respired from root and rhizosphere free soil (δ13CRh, panel b) over the days after adding 100 mm of water in the grazed (orange circles) and mown (blue circles) grasslands. Vertical bars represent one standard error of the mean (n = 4).
Author Contributions
Conceptualization, P.M., A.C. and C.R.; Methodology, P.M., A.J.M. and J.E.H.; Formal Analysis, G.Y.K.M.; Investigation, G.Y.K.M., A.J.M., J.E.H. and P.M.; Resources, A.C. and P.M.; Writing—Original Draft Preparation, G.Y.K.M.; Writing—Review and Editing, C.R., A.C.; A.J.M. and P.M.; Project Administration, G.Y.K.M., P.M. and A.C.; Funding Acquisition, P.M & A.C.
Funding
This research was funded by ENVRIPlus transnational access grant—Grant Agreement number 654182- delivered to GYKM, and by the New Zealand Government to support the objectives of the Livestock Research Group of the Global Research Alliance on Agricultural Greenhouse Gases - Grants SOW12-GPLER-LCR-PM (Proposal ID 16949-15 LCR).
Acknowledgments
We are grateful to Xavier Charrier, François Gastal, Jerome Chargelegue, Jean-François Bouhiron, Christophe de Berranger, Christophe Huguet, Camille Bartolini, and Aicha Chabbi for help with site access, organization, and field measurements.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Rs | Soil respiration |
| δ13CRs | 13C isotopic signature of CO2 respired from the whole soil |
| δ13CRa | 13C isotopic signature of CO2 respired from roots |
| δ13CRh | 13C isotopic signature of CO2 respired from root and rhizosphere free soil |
| θs | Soil volumetric soil water content |
| Ts | Soil temperature at 100 mm depth |
References
- Steinfeld, H.; Gerber, P.; Wassenaar, T.; Castel, V.; Rosales, M.; Haan, C. Livestock’s Long Shadow: Environmental Issues and Options; Food & Agriculture Organization: Rome, Italy, 2006. [Google Scholar]
- Stockmann, U.; Adams, M.A.; Crawford, J.W.; Field, D.J.; Henakaarchchi, N.; Jenkins, M.; Minasny, B.; McBratney, A.B.; de Courcelles, V.D.R.; Singh, K.; et al. The knowns, known unknowns and unknowns of sequestration of soil organic carbon. Agric. Ecosyst. Environ. 2013, 164, 80–99. [Google Scholar] [CrossRef]
- De Coninck, H.; Revi, A.; Babiker, M.; Bertoldi, P.; Buckeridge, M.; Cartwright, A.; Dong, W.; Ford, J.; Fuss, S.; Hourcade, J.C.; et al. Strengthening and implementing the global response. In Global Warming of 1.5 °C, An IPCC Special Report; Masson-Delmotte, V., Zhai, P., Pörtner, H.O., Roberts, D., Skea, J., Shukla, P.R., Pirani, A., Eds.; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2018. [Google Scholar]
- Paustian, K.; Lehmann, J.; Ogle, S.; Reay, D.; Robertson, G.P.; Smith, P. Climate-smart soils. Nature 2016, 532, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Chabbi, A.; Lehmann, L.; Ciais, P.; Loescher, H.L.; Cotrufo, M.F.; Don, A.; San-Clements, M.; Schipper, L.; Six, J.; Smith, P.; et al. Aligning agriculture and climate policy. Nat. Clim. Chang. 2017, 7, 307–309. [Google Scholar] [CrossRef]
- Rumpel, C.; Crème, A.; Ngo, P.T.; Velásquez, G.; Mora, M.L.; Chabbi, A. The impact of grassland management on biogeochemical cycles involving carbon, nitrogen and phosphorus. J. Soil Sci. Plant Nutr. 2015, 15, 353–371. [Google Scholar] [CrossRef]
- Dignac, M.-F.; Derrien, D.; Barré, P.; Barot, S.; Cécillon, L.; Chenu, C.; Chevallier, T.; Freschet, G.T.; Garnier, P.; Guenet, B.; et al. Increasing soil carbon storage: Mechanisms, effects of agricultural practices and proxies. A review. Agron. Sustain. Dev. 2017, 37, 14. [Google Scholar] [CrossRef]
- Whitehead, D.; Schipper, L.A.; Pronger, J.; Moinet, G.Y.K.; Mudge, P.L.; Calvelo Pereira, R.; Kirschbaum, M.U.F.; McNally, S.R.; Beare, M.H.; Camps-Arbestain, M. Management practices to reduce losses or increase soil carbon stocks in temperate grazed grasslands: New Zealand as a case study. Agric. Ecosyst. Environ. 2018, 265, 432–443. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Andrews, J.A. Soil respiration and the global carbon cycle. Biogeochemistry 2000, 48, 7–20. [Google Scholar] [CrossRef]
- Field, C.B.; Intergovernmental Panel on Climate Change; Working Group II. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2014. [Google Scholar]
- Borken, W.; Matzner, E. Reappraisal of drying and wetting effects on C and N mineralization and fluxes in soils. Glob. Chang. Biol. 2009, 15, 808–824. [Google Scholar] [CrossRef]
- Tezara, W.; Mitchell, V.J.; Driscoll, S.D.; Lawlor, D.W. Water stress inhibits plant photosynthesis by decreasing coupling factor and ATP. Nature 1999, 401, 914–917. [Google Scholar] [CrossRef]
- Moyano, F.E.; Manzoni, S.; Chenu, C. Responses of soil heterotrophic respiration to moisture availability: An exploration of processes and models. Soil Biol. Biochem. 2013, 59, 72–85. [Google Scholar] [CrossRef]
- Davidson, E.A.; Verchot, L.V.; Cattânio, J.H.; Ackerman, I.L.; Carvalho, J.E.M. Effects of soil water content on soil respiration. Biogeochemistry 2000, 48, 53–69. [Google Scholar] [CrossRef]
- Moinet, G.Y.K.; Cieraad, E.; Turnbull, M.H.; Whitehead, D. Effects of irrigation and addition of nitrogen fertiliser on net ecosystem carbon balance for a grassland. Sci. Total Environ. 2017, 579, 1715–1725. [Google Scholar] [CrossRef] [PubMed]
- Canarini, A.; Dijkstra, F.A. Dry-rewetting cycles regulate wheat carbon rhizodeposition, stabilization and nitrogen cycling. Soil Biol. Biochem. 2015, 81, 195–203. [Google Scholar] [CrossRef]
- Moinet, G.Y.K.; Cieraad, E.; Hunt, J.E.; Fraser, A.; Turnbull, M.H.; Whitehead, D. Soil heterotrophic respiration is insensitive to changes in soil water content but related to microbial access to organic matter. Geoderma 2016, 274, 68–78. [Google Scholar] [CrossRef]
- Huang, S.; Ye, G.; Lin, J.; Chen, K.; Xu, X.; Ruan, H.; Tan, F.; Chen, H.Y.H. Autotrophic and heterotrophic soil respiration responds asymmetrically to drought in a subtropical forest in the Southeast China. Soil Biol. Biochem. 2018, 123, 242–249. [Google Scholar] [CrossRef]
- Millard, P.; Midwood, A.J.; Hunt, J.E.; Barbour, M.M.; Whitehead, D. Quantifying the contribution of soil organic matter turnover to forest soil respiration, using natural abundance δ13C. Soil Biol. Biochem. 2010, 42, 935–943. [Google Scholar] [CrossRef]
- Moinet, G.Y.K.; Hunt, J.E.; Kirschbaum, M.U.F.; Morcom, C.P.; Midwood, A.J.; Millard, P. The temperature sensitivity of soil organic matter decomposition is constrained by microbial access to substrates. Soil Biol. Biochem. 2018, 116, 333–339. [Google Scholar] [CrossRef]
- Moinet, G.Y.K.; Midwood, A.J.; Hunt, J.E.; Whitehead, D.; Hannam, K.D.; Jenkins, M.; Brewer, M.J.; Adams, M.A.; Millard, P. Estimates of rhizosphere priming effects are affected by soil disturbance. Geoderma 2018, 313, 1–6. [Google Scholar] [CrossRef]
- Boström, B.; Comstedt, D.; Ekblad, A. Isotope fractionation and 13C enrichment in soil profiles during the decomposition of soil organic matter. Oecologia 2007, 153, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Snell, H.S.K.; Robinson, D.; Midwood, A.J. Minimising methodological biases to improve the accuracy of partitioning soil respiration using natural abundance 13C. Rapid Commun. Mass Spectrom. 2014, 28, 2341–2351. [Google Scholar] [CrossRef] [PubMed]
- Soussana, J.F.; Allard, V.; Pilegaard, K.; Ambus, P.; Amman, C.; Campbell, C.; Ceschia, E.; Clifton-Brown, J.; Czobel, S.; Domingues, R.; et al. Full accounting of the greenhouse gas (CO2, N2O, CH4) budget of nine European grassland sites. Agric. Ecosyst. Environ. 2007, 121, 121–134. [Google Scholar] [CrossRef]
- Soussana, J.F.; Tallec, T.; Blanfort, V. Mitigating the greenhouse gas balance of ruminant production systems through carbon sequestration in grasslands. Animal 2010, 4, 334–350. [Google Scholar] [CrossRef] [PubMed]
- FAO-ISRIC. Guidelines for Soil Description, revised 3rd ed.; Food and Agricultural Organisation: Rome, Italy, 1990. [Google Scholar]
- Chabbi, A.; Kögel-Knabner, I.; Rumpel, C. Stabilised carbon in subsoil horizons is located in spatially distinct parts of the soil profile. Soil Biol. Biochem 2009, 41, 256–261. [Google Scholar] [CrossRef]
- Senapati, N.; Chabbi, A.; Giostri, A.F.; Yeluripati, J.B.; Smith, P. Modelling nitrous oxide emissions from mown-grass and grain-cropping systems: Testing and sensitivity analysis of DailyDayCent using high frequency measurements. Sci. Total Environ. 2016, 572, 955–977. [Google Scholar] [CrossRef] [PubMed]
- Midwood, A.J.; Thornton, B.; Millard, P. Measuring the 13C content of soil-respired CO2 using a novel open chamber system. Rapid Commun. Mass Spectrom. 2008, 22, 2073–2081. [Google Scholar] [CrossRef] [PubMed]
- Midwood, A.J.; Millard, P. Challenges in measuring the δ13C of the soil surface CO2 efflux. Rapid Commun. Mass Spectrom. 2011, 25, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Gail, M., Krickeberg, K., Samet, J.M., Tsiatis, A., Wong, W., Eds.; Spring Science and Business Media: New York, NY, USA, 2009. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2003. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Powers, H.H.; Hunt, J.E.; Hanson, D.T.; McDowell, N.G. A dynamic soil chamber system coupled with a tunable diode laser for online measurements of δ13C, δ18O, and efflux rate of soil-respired CO2: Measurements of δ13C, δ18O, and efflux rate of soil-respired CO2. Rapid Commun. Mass Spectrom. 2010, 24, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Schimel, J.P. Effects of drying–rewetting frequency on soil carbon and nitrogen transformations. Soil Biol. Biochem. 2002, 34, 777–787. [Google Scholar] [CrossRef]
- Read, J.; Farquhar, G. Comparative studies in Nothofagus (Fagaceae). I. Leaf carbon isotope discrimination. Funct. Ecol. 1991, 5, 684–695. [Google Scholar] [CrossRef]
- Schulze, E.-D.; Williams, R.J.; Farquhar, G.D.; Schulze, W.; Langridge, J.; Miller, J.M.; Walker, B.H. Carbon and nitrogen isotope discrimination and nitrogen nutrition of trees along a rainfall gradient in northern Australia. Funct. Plant Biol. 1998, 25, 413–425. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Gavrichkova, O. Review: Time lag between photosynthesis and carbon dioxide efflux from soil: A review of mechanisms and controls. Glob. Chang. Biol. 2010, 16, 3386–3406. [Google Scholar] [CrossRef]
- Barthel, M.; Cieraad, E.; Zakharova, A.; Hunt, J.E. Sudden cold temperature delays plant carbon transport and shifts allocation from growth to respiratory demand. Biogeosciences 2014, 11, 1425–1433. [Google Scholar] [CrossRef]
- Raich, J.W.; Schlesinger, W.H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B 1992, 44, 81–99. [Google Scholar] [CrossRef]
- Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Subke, J.-A.; Inglima, I.; Francesca Cotrufo, M. Trends and methodological impacts in soil CO2 efflux partitioning: A metaanalytical review. Glob. Chang. Biol. 2006, 12, 921–943. [Google Scholar] [CrossRef]
- Flanagan, L.B.; Wever, L.A.; Carlson, P.J. Seasonal and interannual variation in carbon dioxide exchange and carbon balance in a northern temperate grassland. Glob. Chang. Biol. 2002, 8, 599–615. [Google Scholar] [CrossRef]
- Hunt, J.E.; Kelliher, F.M.; McSeveny, T.M.; Ross, D.J.; Whitehead, D. Long-term carbon exchange in a sparse, seasonally dry tussock grassland. Glob. Chang. Biol. 2004, 10, 1785–1800. [Google Scholar] [CrossRef]
- Zhang, L.; Guo, H.; Jia, G.; Wylie, B.; Gilmanov, T.; Howard, D.; Ji, L.; Xiao, J.; Li, J.; Yuan, W.; et al. Net ecosystem productivity of temperate grasslands in northern China: An upscaling study. Agric. For. Meteorol. 2014, 184, 71–81. [Google Scholar] [CrossRef]
- Sanaullah, M.; Chabbi, A.; Rumpel, C.; Kuzyakov, Y. Carbon allocation in grassland communities under drought stress followed by 14C pulse labeling. Soil Biol. Biochem. 2012, 55, 132–139. [Google Scholar] [CrossRef]
- White, L.M. Carbohydrate reserves of grasses: A review. J. Range Manag. 1973, 26, 13–18. [Google Scholar] [CrossRef]
- Steen, E.; Larsson, K. Carbohydrates in roots and rhizomes of perennial grasses. New Phytol. 1986, 104, 339–346. [Google Scholar] [CrossRef]
- Donaghy, D.J.; Fulkerson, W.J. The importance of water-soluble carbohydrate reserves on regrowth and root growth of Lolium perenne (L.). Grass Forage Sci. 1997, 52, 401–407. [Google Scholar] [CrossRef]
Figure 1.
Soil volumetric water content (θs, panel a) and soil respiration (RS, panel b) before and over 24 h after the addition of 100 mm of water in the grazed treatment for watered plots (diamonds) and control non-watered plots (circles). The line in panel b is the difference between mean Rs in the watered plots and mean Rs in the control plot for each hour after watering. Vertical bars represent one standard error of the mean (n = 4).
Figure 1.
Soil volumetric water content (θs, panel a) and soil respiration (RS, panel b) before and over 24 h after the addition of 100 mm of water in the grazed treatment for watered plots (diamonds) and control non-watered plots (circles). The line in panel b is the difference between mean Rs in the watered plots and mean Rs in the control plot for each hour after watering. Vertical bars represent one standard error of the mean (n = 4).

Figure 2.
Soil volumetric water content (θs, panel a), soil respiration (Rs, panel b) and 13C isotopic signature of soil respired CO2 (δ13CRs, panel c) over the days after adding 100 mm of water in the grazed (orange circles) and mown (blue circles) grasslands. Vertical bars represent one standard error of the mean (n = 4).
Figure 2.
Soil volumetric water content (θs, panel a), soil respiration (Rs, panel b) and 13C isotopic signature of soil respired CO2 (δ13CRs, panel c) over the days after adding 100 mm of water in the grazed (orange circles) and mown (blue circles) grasslands. Vertical bars represent one standard error of the mean (n = 4).

Figure 3.
Soil respiration (RS) as a function of root biomass (panel a) and soil volumetric water content (θs, panel b) for the grazed (orange circles) and mown (blue circles) grasslands. The lines represent the fit of linear regressions. The black line in panel a indicates identical fit for the two treatments. Only significant correlations appear.
Figure 3.
Soil respiration (RS) as a function of root biomass (panel a) and soil volumetric water content (θs, panel b) for the grazed (orange circles) and mown (blue circles) grasslands. The lines represent the fit of linear regressions. The black line in panel a indicates identical fit for the two treatments. Only significant correlations appear.

Figure 4.
13C isotopic signature of CO2 respired from the roots and rhizosphere (δ13CRa, panel a), the whole soil (δ13CRs, panel b) and the root- and rhizosphere-free soil (δ13CRh, panel c) as functions of soil volumetric water content (θs) for the grazed (orange circles) and mown (blue circles) grasslands. The lines represent the fit of linear regressions. Only significant correlations appear.
Figure 4.
13C isotopic signature of CO2 respired from the roots and rhizosphere (δ13CRa, panel a), the whole soil (δ13CRs, panel b) and the root- and rhizosphere-free soil (δ13CRh, panel c) as functions of soil volumetric water content (θs) for the grazed (orange circles) and mown (blue circles) grasslands. The lines represent the fit of linear regressions. Only significant correlations appear.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Moinet, G.Y.K.; Midwood, A.J.; Hunt, J.E.; Rumpel, C.; Millard, P.; Chabbi, A. Grassland Management Influences the Response of Soil Respiration to Drought. Agronomy 2019, 9, 124. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030124
AMA Style
Moinet GYK, Midwood AJ, Hunt JE, Rumpel C, Millard P, Chabbi A. Grassland Management Influences the Response of Soil Respiration to Drought. Agronomy. 2019; 9(3):124. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030124
Chicago/Turabian StyleMoinet, Gabriel Y. K., Andrew J. Midwood, John E. Hunt, Cornelia Rumpel, Peter Millard, and Abad Chabbi. 2019. "Grassland Management Influences the Response of Soil Respiration to Drought" Agronomy 9, no. 3: 124. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9030124
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.