Brassinosteroids Regulate Antioxidant System and Protect Chloroplast Ultrastructure of Autotoxicity-Stressed Cucumber (Cucumis sativus L.) Seedlings


Abstract
:1. Introduction
2. Materials and Methods
2.1. Determination of Morphological Characteristics
2.2. Measurement of Biomass
2.3. Determination of Root Morphology and Root Activity
2.4. Determination of Photosynthetic Pigments and Observation of Stomatal Opening
2.5. Determination of Gas Exchange Parameters
2.6. Ultrastructure of Chloroplast and Thylakoid
2.7. ROS and Lipid Peroxidation Detection and Quantification
2.8. Activity of Antioxidant Enzymes and Contents of Proline, Soluble Sugar, and Protein
2.9. Statistical Analysis
3. Results
3.1. Morphological Changes
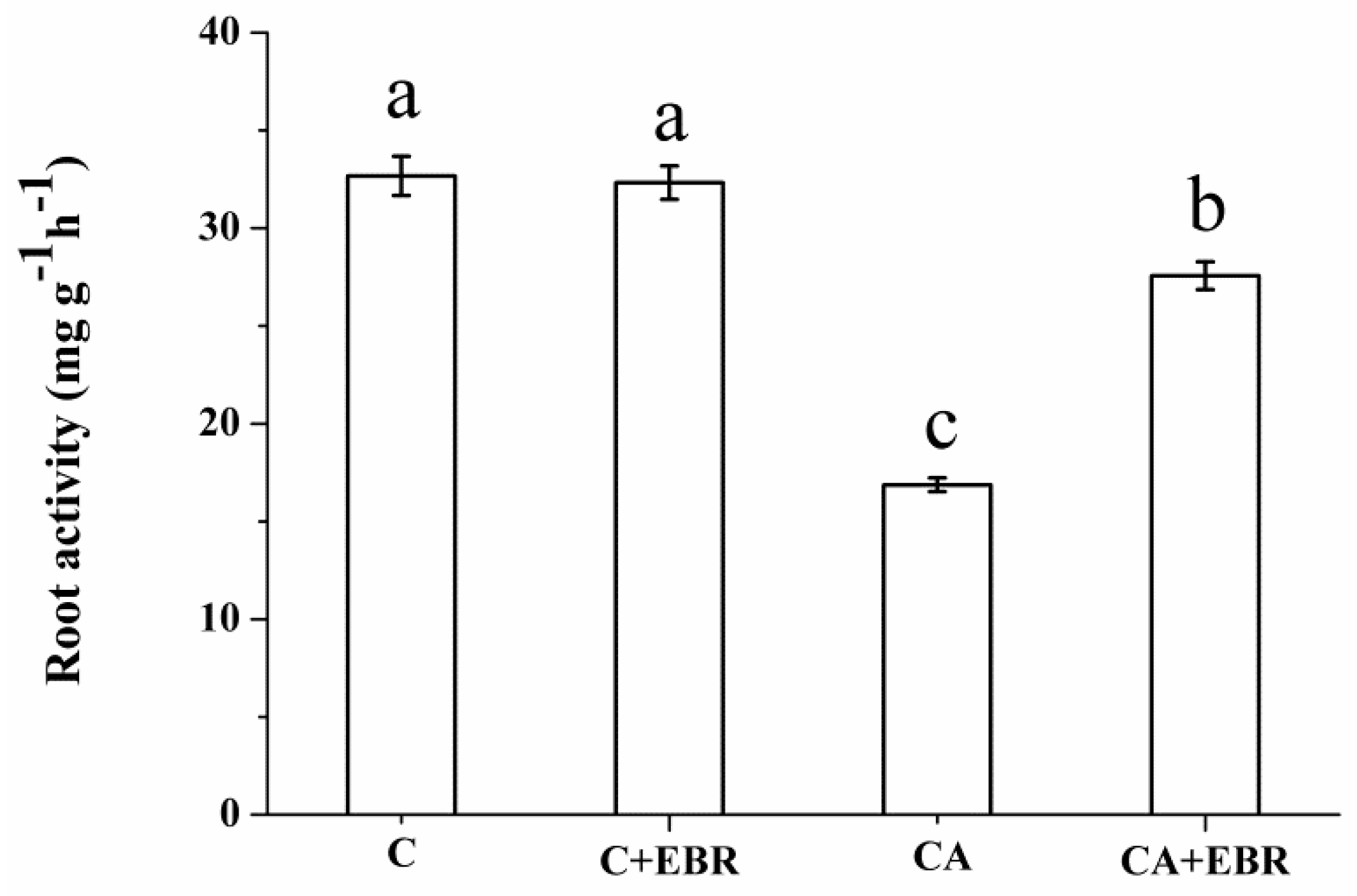
3.2. Root Morphology and Root Activity
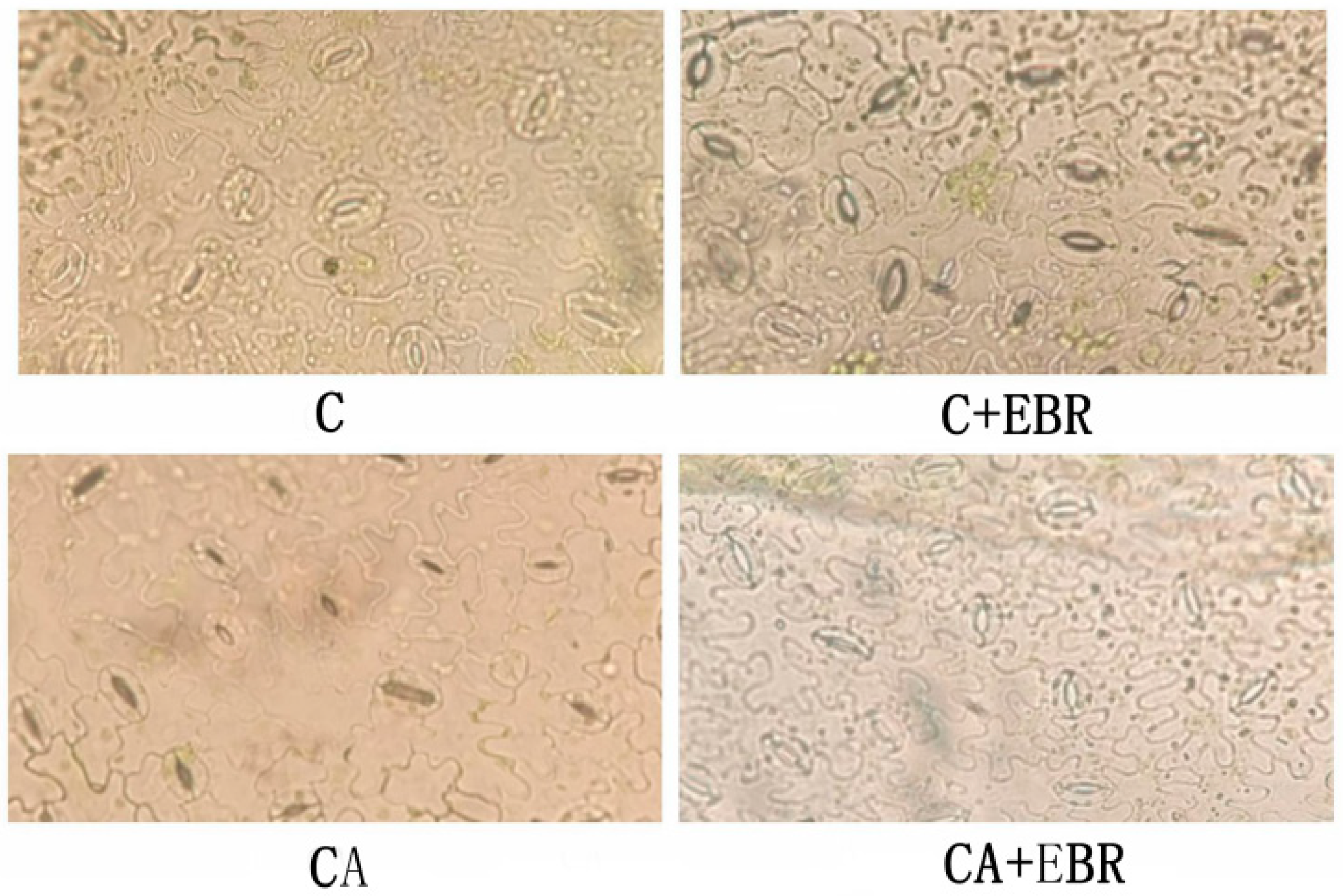
3.3. Photosynthetic Pigments and Stomatal Opening
3.4. Gas Exchange
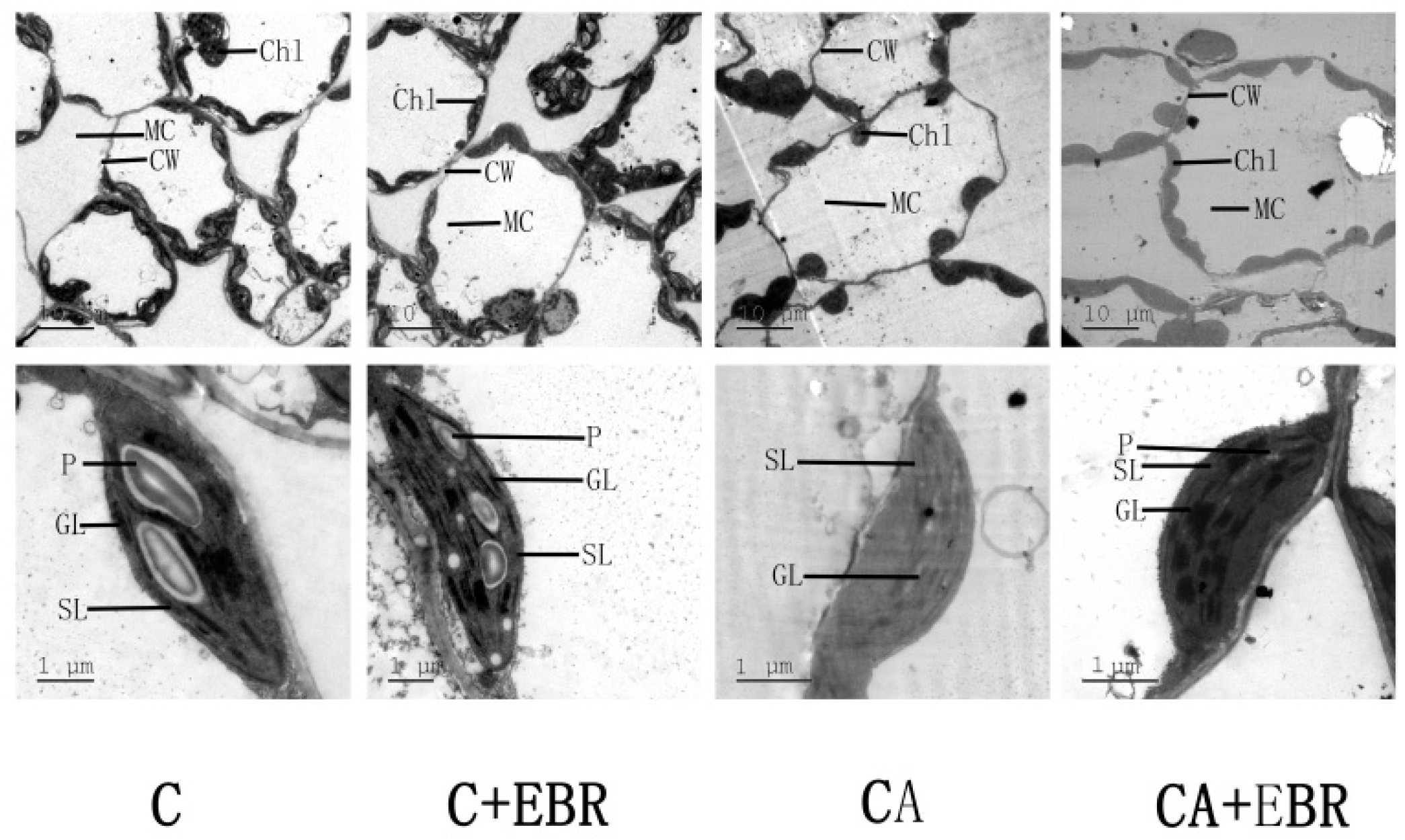
3.5. Ultrastructure of Chloroplast and Thylakoid
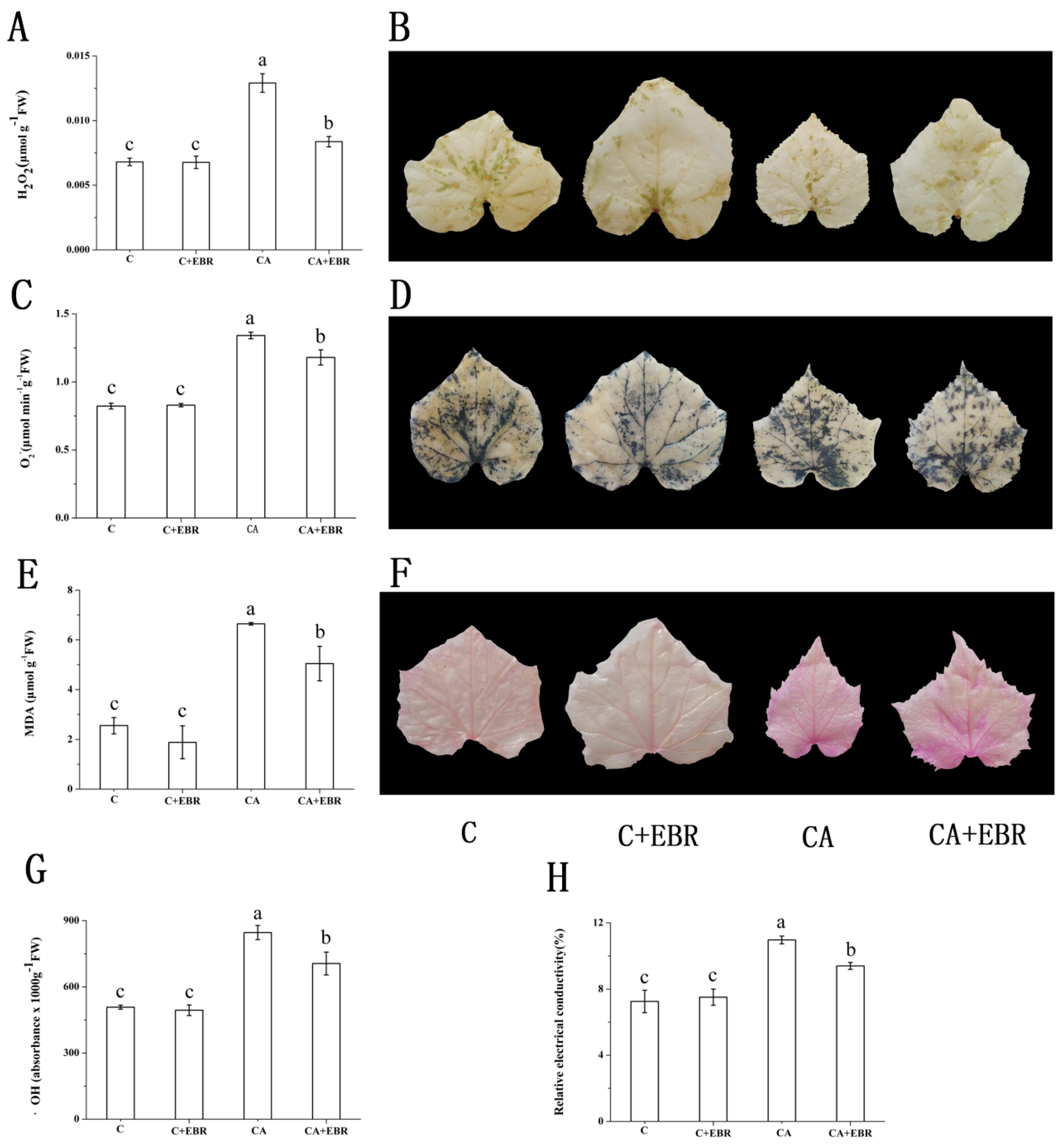
3.6. ROS, MDA and Relative Membrane Permeability
3.7. Antioxidant System
3.8. Proline, Soluble Sugar and Protein Contents
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Gu, M.; Xia, X.; Shi, K.; Zhou, Y.; Yu, J. Alleviation of autotoxin-induced growth in hibition and respiration by sucrose in Cucumis sativus (L.). Allelopathy J. 2010, 25, 147–154. [Google Scholar]
- Paudel, M.N. Multiple Cropping for Raising Productivity and Farm Income of Small Farmers. J. Nepal Agric. Res. Counc. 2016, 2, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Chou, C. Roles of allelopathy in plant biodiversity and sustainable agriculture. Crit. Rev. Plant Sci. 1999, 18, 609–636. [Google Scholar] [CrossRef]
- Bie, Z.; Nawaz, M.A.; Huang, Y.; Lee, J.M.; Colla, G. Introduction of vegetable grafting. In Vegetable Grafting, Principles and Practices; Colla, G., Alfocea, F.P., Schwarz, D., Eds.; CABI Publishing: Wallingford, UK, 2017; pp. 1–21. [Google Scholar]
- Callaway, R.; Vivanco, J. Can plant biochemistry contribute to understanding of invasion ecology? Trends Plant Sci. 2006, 11, 574–580. [Google Scholar] [CrossRef]
- Qiao, Y. Study on Different Responses to Autotoxin Toxicity in Cucumber and Figleaf Gourd Seedlings. Ph.D. Thesis, China Agricultural University, Beijing, China, 2013. [Google Scholar]
- Li, J.; Li, Y.; Tian, Y.; Qu, M.; Zhang, W.; Gao, L. Melatonin has the potential to alleviate cinnamic acid stress in cucumber seedlings. Front. Plant Sci. 2017, 8, 1193. [Google Scholar] [CrossRef]
- Seal, A.; Haig, T.; Pratley, J. Evaluation of putative allelochemicals in rice root exudates for their role in the suppression of arrowhead root growth. J. Chem. Ecol. 2004, 30, 1663–1678. [Google Scholar] [CrossRef]
- Belz, R. Allelopathy in crop/weed interactions-an update. Pest Manag. Sci. 2007, 63, 308–326. [Google Scholar] [CrossRef]
- FAO. FAOSTAT Domains/Production/Crops. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 19 December 2016).
- Xie, J.; Yu, J.; Chen, B.; Feng, Z.; Li, J.; Zhao, C.; Lyu, J.; Hu, L.; Gan, Y.; Kadambot, H. Facility cultivation systems “设施农业”: A Chinese model for the planet. Adv. Agron. 2017, 145, 1–42. [Google Scholar] [CrossRef]
- Bennett, A.; Bending, G.; Chandler, D.; Hilton, S.; Mills, P. Meeting the demand for crop production: The challenge of yield decline in crops grown in short rotations. Biol. Rev. 2012, 87, 52–71. [Google Scholar] [CrossRef] [PubMed]
- Al-Harbi, A.; Al-Omran, A.; Alharbi, K. Grafting improves cucumber water stress tolerance in Saudi Arabia. Saudi J. Biol. Sci. 2018, 25, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.A.; Shireen, F.; Huang, Y.; Bie, Z.; Waqar, A.; Basharat, A. Perspectives of vegetable grafting in Pakistan, current status, challenges and opportunities. Int. Agric. Biol. 2017, 19, 1165–1174. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Muhamamd, I.; Kong, Q.; Cheng, F.; Waqar, A.; Huang, Y.; Bie, Z. Grafting: A technique to modify ion accumulation in horticultural crops. Front. Plant Sci. 2016, 7, 1457. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Jiao, Y.; Nawaz, M.A.; Chen, C.; Liu, L.; Lu, Z.; Kong, Q.; Cheng, F.; Bie, Z. Improving magnesium uptake, photosynthesis and antioxidant enzyme activities of watermelon by grafting onto pumpkin rootstock under low magnesium. Plant Soil 2016, 409, 229–246. [Google Scholar] [CrossRef]
- Alfocea, P.; Schwarz, D. (Eds.) Vegetable Grafting. Principles and Practices; CABI Publishing: Wallingford, UK, 2017; pp. 1–21. [Google Scholar]
- Velkov, N.; Pevicharova, G. Effects of cucumber grafting on yield and fruit sensory characteristics. Zemdirbyste 2016, 103, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Vriet, C.; Russinova, E.; Reuzeau, C. Boosting crop yields with plant steroids. Plant Cell 2012, 24, 842–857. [Google Scholar] [CrossRef]
- Krishna, P. Brassinosteroid-mediated stress responses. J. Plant Growth Regul. 2003, 22, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Bajguz, A.; Hayat, S. Effects of brassinosteroids on the plant responses to environmental stresses. Plant Physiol. Biochem. 2009, 47, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Zhou, Y.; Ding, J.; Xia, X.; Shi, K.; Chen, S.; Asami, T.; Chen, Z.; Yu, J. Role of nitric oxide in hydrogenperoxide-dependent induction of abiotic stress tolerance by brassinosteroids in cucumber. Plant Cell Environ. 2011, 34, 347–358. [Google Scholar] [CrossRef]
- Xia, X.; Fang, P.; Guo, X.; Qian, X.; Zhou, J.; Shi, K.; Zhou, Y.; Yu, J. Brassinosteroid-mediated apoplastic H2O2-glutaredoxin 12/14 cascade regulates antioxidant capacity in response to chilling in tomato. Plant Cell Environ. 2018, 41, 1052–1064. [Google Scholar] [CrossRef]
- Hu, W.; Wu, Y.; Zeng, J.; He, L.; Zeng, Q. Chill-induced inhibition of photosynthesis was alleviated by 24-epibrassinolide pretreatment in cucumber during chilling and subsequent recovery. Photosynthetica 2010, 48, 537–544. [Google Scholar] [CrossRef]
- Eremina, M.; Unterholzner, S.; Rathnayake, A.; Castellanos, M.; Khan, M.; Kugler, K.; Poppenberger, B. Brassinosteroids participate in the control of basal and acquired freezing tolerance of plants. Proc. Natl. Acad. Sci. USA 2016, 113, E5982–E5991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Huang, L.; Cheng, F.; Zhou, Y.; Xia, X.; Mao, W.; Shi, K.; Yu, J. Brassinosteroids accelerate recovery of photosynthetic apparatus from cold stress by balancing the electron partitioning, carboxylation and redox homeostasis in cucumber. Physiol. Plant. 2013, 148, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Todorova, D.; Talaat, N.B.; Katerova, Z.; Alexieva, V.; Shawky, B. Polyamines and brassinosteroids in drought stress responses and tolerance in plants. In Water Stress and Crop Plants, 1st ed.; Ahmad, P., Ed.; John Wiley & Sons, Ltd.: Hoboken, NY, USA, 2016; pp. 608–627. [Google Scholar]
- Liu, Z.; Li, L.; Luo, Z.; Zeng, F.; Jiang, L.; Tang, K. Effect of brassinolide on energy status and proline metabolism in postharvest bamboo shoot during chilling stress. Postharvest Biol. Technol. 2016, 111, 240–246. [Google Scholar] [CrossRef]
- Kang, Y.; Guo, S.; Li, J.; Duan, J. Effect of root applied 24-epibrassinolide on carbohydrate status and fermentative enzyme activities in cucumber (Cucumis sativus L.) seedlings under hypoxia. Plant Growth Regul. 2008, 57, 259–269. [Google Scholar] [CrossRef]
- Bu, R.; Wang, R.; Wei, Q.; Hu, H.; Sun, H.; Song, P.; Yu, Y.; Liu, Q.; Zheng, Z.; Li, T.; et al. Silencing of glycerol-3-phosphate acyltransferase 6 (GPAT6) gene using a newly established virus induced gene silencing (VIGS) system in cucumber alleviates autotoxicity mimicked by cinnamic acid (CA). Plant Soil. 2019, 438, 329–346. [Google Scholar] [CrossRef]
- Li, J.; Yang, P.; Xie, J.; Yu, J. Effects of 24-epibrassinolide on growth and antioxidant enzymes system in pepper roots under chilling stress. J. Nucl. Agric. Sci. 2015, 29, 1001–1008. [Google Scholar] [CrossRef]
- Arnon, D. Copper enzymes in isolated chloroplasts polyphenoloxidase in beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Zhang, Y.; Xiang, J.; Wu, H.; Chen, H.; Zhang, Y.; Zhu, D. Effects of chilling tolerance induced by spermidine pretreatment on antioxidative activity, endogenous hormones and ultrastructure of indica-japonica hybrid rice seedlings. J. Integr. Agric. 2016, 15, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Ma, N.; Zuo, Q.; Liang, X.; Yin, B.; Wang, Q.; Meng, D. The multiple stress-responsive transcription factor SlNAC1 improves the chilling tolerance of tomato. Physiol. Plant. 2013, 149, 474–486. [Google Scholar] [CrossRef]
- Li, J.; Yang, P.; Gan, Y.; Yu, J.; Xie, J. Brassinosteroid alleviates chilling-induced oxidative stress in pepper by enhancing antioxidation systems and maintenance of photosystem II. Acta Physiol. Plant. 2015, 37, 222. [Google Scholar] [CrossRef]
- Stevens, R.; Page, D.; Gouble, B.; Garchery, C.; Zamir, D.; Causse, M. Tomato fruit ascorbic acid content is linked with monodehydroascorbate reductase activity and tolerance to chilling stress. Plant Cell Environ. 2008, 31, 1086–1096. [Google Scholar] [CrossRef]
- Griffith, O.W. Determination of glutathione and glutathione disulphide using glutathione reductase and 2-vinylpyridine. Anal. Biochem. 1980, 106, 207–212. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Buysse, J.; Merckx, R. An improved colorimetric method to quantify sugar content of plant tissue. J. Exp. Bot. 1993, 44, 1627–1629. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sadura, I.; Janeczko, A. Physiological and molecular mechanisms of brassinosteroid-induced tolerance to high and low temperature in plants. Biol. Plant. 2018, 62, 601–616. [Google Scholar] [CrossRef] [Green Version]
- Shahbaz, M.; Ashraf, M.; Athar, H. Does exogenous application of 24-epibrassinolide ameliorate salt induced growth inhibition in wheat (Triticum aestivum L.)? Plant Growth Regul. 2008, 55, 51–64. [Google Scholar] [CrossRef]
- Sharma, I.; Pati, P.; Bhardwaj, R. Effect of 24-epibrassinolide on oxidative stress markers induced by nickel-ion in Raphanus sativus L. Acta Physiol. Plant. 2011, 33, 1723–1735. [Google Scholar] [CrossRef]
- Cheng, J.; Dai, T.; Jing, Q.; Jiang, D.; Pan, X.; Cao, W. Root morphological and physiological characteristics in relation to nitrogen absorption efficiency in different rice genotypes. Acta Pedol. Sin. 2007, 44, 266–272. [Google Scholar] [CrossRef]
- Pierret, A.; Doussan, C.; Capowiez, Y.; Bastardie, F.; Pagès, L. Root functional architecture: A framework for modeling the interplay between roots and soil. Vadose Zone J. 2007, 6, 269–281. [Google Scholar] [CrossRef]
- Müssig, C.; Shin, G.; Altmann, T. Brassinosteroids promote root growth in Arabidopsis. Plant Physiol. 2003, 133, 1261–1271. [Google Scholar] [CrossRef]
- Li, L.; Xu, J.; Xu, Z.; Xue, H. Brassinoeroids stimulate plant tropisms through modulation of polar auxin transport in Brassica and Arabidopsis. Plant Cell 2005, 17, 2738–2753. [Google Scholar] [CrossRef] [PubMed]
- Ahammed, G.; Yuan, L.; Ogweno, J.; Zhou, Y.; Xia, X.; Mao, W.; Shi, K.; Yu, J. Brassinosteroid alleviates phenanthrene and pyrene phytotoxicity by increasing detoxification activity and photosynthesis in tomato. Chemosphere 2012, 86, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Wen, L. 2-Photonic sensitive switchable materials A2-Meglinski, Igor. In Biophotonics for Medical Applications; Woodhead Publishing: Cambridge, UK, 2015; pp. 25–51. [Google Scholar]
- Xia, X.; Wang, Y.; Zhou, Y.; Tao, Y.; Mao, W.; Shi, K.; Asami, T.; Chen, Z.; Yu, J. Reactive oxygen species are involved in brassinosteroid-induced stress tolerance in cucumber. Plant Physiol. 2009, 150, 801–814. [Google Scholar] [CrossRef]
- Xia, X.; Huang, L.; Zhou, Y.; Mao, W.; Shi, K.; Wu, J.; Asami, T.; Chen, Z.; Yu, J. Brassinosteroids promote photosynthesis and growth by enhancing activation of Rubisco and expression of photosynthetic genes in Cucumis sativus. Planta 2009, 230, 1185–1196. [Google Scholar] [CrossRef]
- Cao, S.; Xu, Q.; Cao, Y.; Qian, K.; An, K.; Zhu, Y.; Hu, B.; Zhao, H.; Kuai, B. Loss of function mutations in DET2 gene lead to an enhanced resistance to oxidative stress in Arabidopsis. Physiol. Plant. 2005, 123, 57–66. [Google Scholar] [CrossRef]
- Muhammad, A.; Chen, C.; Fareeha, S.; Zheng, Z.; Jiao, Y.; Hamza, S.; Muhammad, A.; Muhammad, I.; Muhammad, A.; Huang, Y.; et al. Improving vanadium stress tolerance of watermelon by grafting onto bottle gourd and pumpkin rootstock. Plant Growth Regul. 2018, 85, 41–56. [Google Scholar] [CrossRef]
- Wang, P.; Sun, X.; Li, C.; Wei, Z.; Liang, D.; Ma, F. Long-term exogenous application of melatonin delays drought-induced leaf senescence in apple. J. Pineal Res. 2013, 54, 292–302. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Plant Height (cm) | Stem Diameter (cm) | Leaf Area (mm2) |
|---|---|---|---|
| C | 16.07 ± 0.53 a | 0.65 ± 0.14 a | 2434.31 ± 25.94 a |
| C + EBR | 16.97 ± 0.23 a | 0.59 ± 0.01 a | 2545.29 ± 67.98 a |
| CA | 9.65 ± 0.61 c | 0.40 ± 0.02 b | 879.72 ± 35.41 c |
| CA + EBR | 14.13 ± 0.94 b | 0.49 ± 0.03 ab | 1583.58 ± 29.81 b |
| Treatment | Fresh Weight (g) | Dry Weight (g) | ||
|---|---|---|---|---|
| Above-Ground | Underground | Above-Ground | Underground | |
| C | 6.67 ± 0.69 a | 0.74 ± 0.12 a | 0.49 ± 0.03 a | 0.054 ± 0.008 a |
| EBR | 7.14 ± 0.70 a | 0.83 ± 0.31 a | 0.48 ± 0.06 a | 0.065 ± 0.012 ab |
| CA | 3.74 ± 0.24 c | 0.63 ± 0.04 a | 0.28 ± 0.03 c | 0.041 ± 0.007 ab |
| CA+EBR | 5.57 ± 0.37 b | 0.73 ± 0.06 a | 0.40 ± 0.01 b | 0.052 ± 0.003 b |
| Treatment | Total Root Length (mm) | Root Surface (cm2) | Root Volume (cm3) | Root Tip Number |
|---|---|---|---|---|
| C | 1642.6 ± 162.0 a | 354.9 ± 27.3 a | 1.42 ± 0.14 a | 161 ± 12 a |
| C + EBR | 1770.3 ± 198.8 a | 331.1 ± 27.3 a | 1.25 ± 0.33 a | 150 ± 22 a |
| CA | 588.2 ± 41.1 c | 169.0 ± 29.2 c | 0.69 ± 0.07 b | 100 ± 7 b |
| CA + EBR | 1005.0 ± 8.5 b | 257.0 ± 40.5 b | 0.32 ± 0.08 c | 69 ± 8 c |
| Treatment | Chl a Content | Chl b Content | Total Chl Content | Carotenoid Content |
|---|---|---|---|---|
| C | 27.19 ± 0.07 a | 31.70 ± 1.03 a | 58.89 ± 1.07 a | 9.93 ± 1.03 a |
| C + EBR | 27.30 ± 0.58 a | 31.80 ± 2.18 a | 59.10 ± 1.83 a | 10.41 ± 1.45 a |
| CA | 21.14 ± 0.05 c | 22.83 ± 1.74 c | 43.97 ± 1.69 c | 6.64 ± 0.47 b |
| CA + EBR | 25.11 ± 1.77 b | 28.09 ± 0.61 b | 53.19 ± 1.54 b | 8.64 ± 0.32 ab |
| Treatment | Pn (μmol (CO2) m−2 s−1) | Ci (μmol mol−1) | Gs (mmol (H2O) m2 s−) | Tr (mmol m−2 s−1) |
|---|---|---|---|---|
| C | 12.7 ± 1.0 a | 341.3 ± 4.7 a | 32.2 ± 3.9 a | 443.3 ± 31.0 a |
| C + EBR | 11.3 ± 1.1 a | 306.7 ± 43.4 a | 23.9 ± 16.3 a | 319.3 ± 146.0 a |
| CA | 4.2 ± 0.3 c | 202.3 ± 51.3 b | 3.5 ± 0.7 b | 71.0 ± 4.2 b |
| CA + EBR | 6.6 ± 1.4 b | 167.0 ± 36.3 b | 4.5 ± 1.4 b | 115.3 ± 20.3 b |
| Treatment | Soluble Protein (mg g−1 Fresh weight) | Soluble Sugar (mg g−1 Fresh weight) | Proline (μg g−1 Fresh weight) |
|---|---|---|---|
| C | 15.90 ± 0.48 c | 0.44 ± 0.02 c | 19.66 ± 3.16 c |
| C + EBR | 17.76 ± 0.74 c | 0.46 ± 0.02 c | 16.37 ± 1.80 c |
| CA | 23.67 ± 0.25 b | 0.67 ± 0.08 b | 68.83 ± 3.16 b |
| CA + EBR | 25.38 ± 1.07 a | 1.07 ± 0.10 a | 94.01 ± 2.89 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, P.; Azher Nawaz, M.; Li, F.; Bai, L.; Li, J. Brassinosteroids Regulate Antioxidant System and Protect Chloroplast Ultrastructure of Autotoxicity-Stressed Cucumber (Cucumis sativus L.) Seedlings. Agronomy 2019, 9, 265. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9050265
Yang P, Azher Nawaz M, Li F, Bai L, Li J. Brassinosteroids Regulate Antioxidant System and Protect Chloroplast Ultrastructure of Autotoxicity-Stressed Cucumber (Cucumis sativus L.) Seedlings. Agronomy. 2019; 9(5):265. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9050265
Chicago/Turabian StyleYang, Ping, Muhammad Azher Nawaz, Fuxin Li, Lisha Bai, and Jie Li. 2019. "Brassinosteroids Regulate Antioxidant System and Protect Chloroplast Ultrastructure of Autotoxicity-Stressed Cucumber (Cucumis sativus L.) Seedlings" Agronomy 9, no. 5: 265. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9050265