Screening, Identification, and Optimization of Fermentation Conditions of an Antagonistic Endophyte to Wheat Head Blight

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Sample Collection and Endophyte Isolation
2.2. Dual Culture Assays for Antagonistic Tests
2.3. Classification of Endophyte Isolates
2.4. Optimization of Fermentation Conditions
2.5. In Vitro Antagonistic Test of Endophyte against F. graminearum
2.6. Statistical Analysis
3. Results
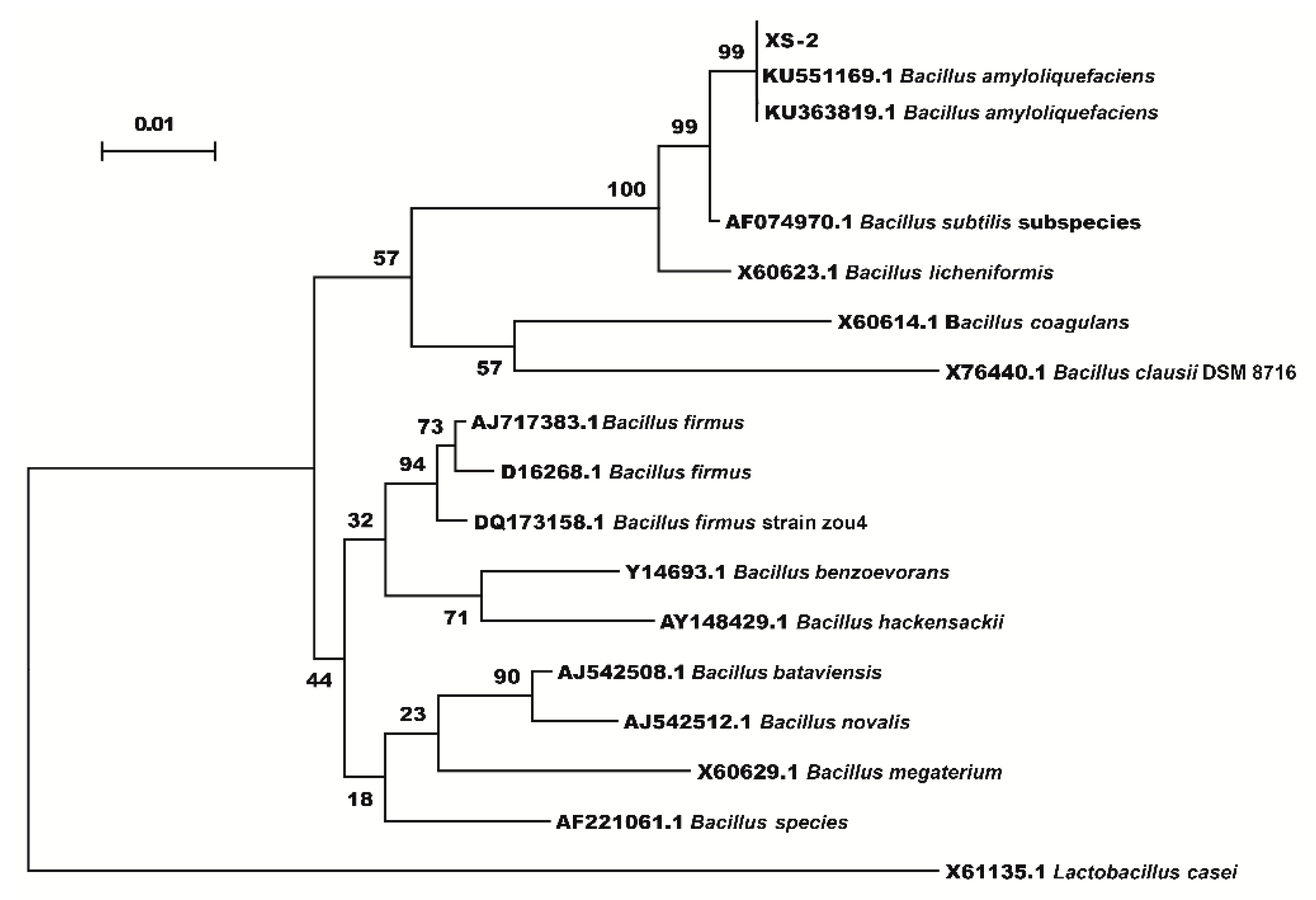
3.1. Identification of Endophytic Bacteria Strain XS-2
3.2. Determination of Antimicrobial Spectrum
3.3. Determination of the Optimal Fermentation Conditions
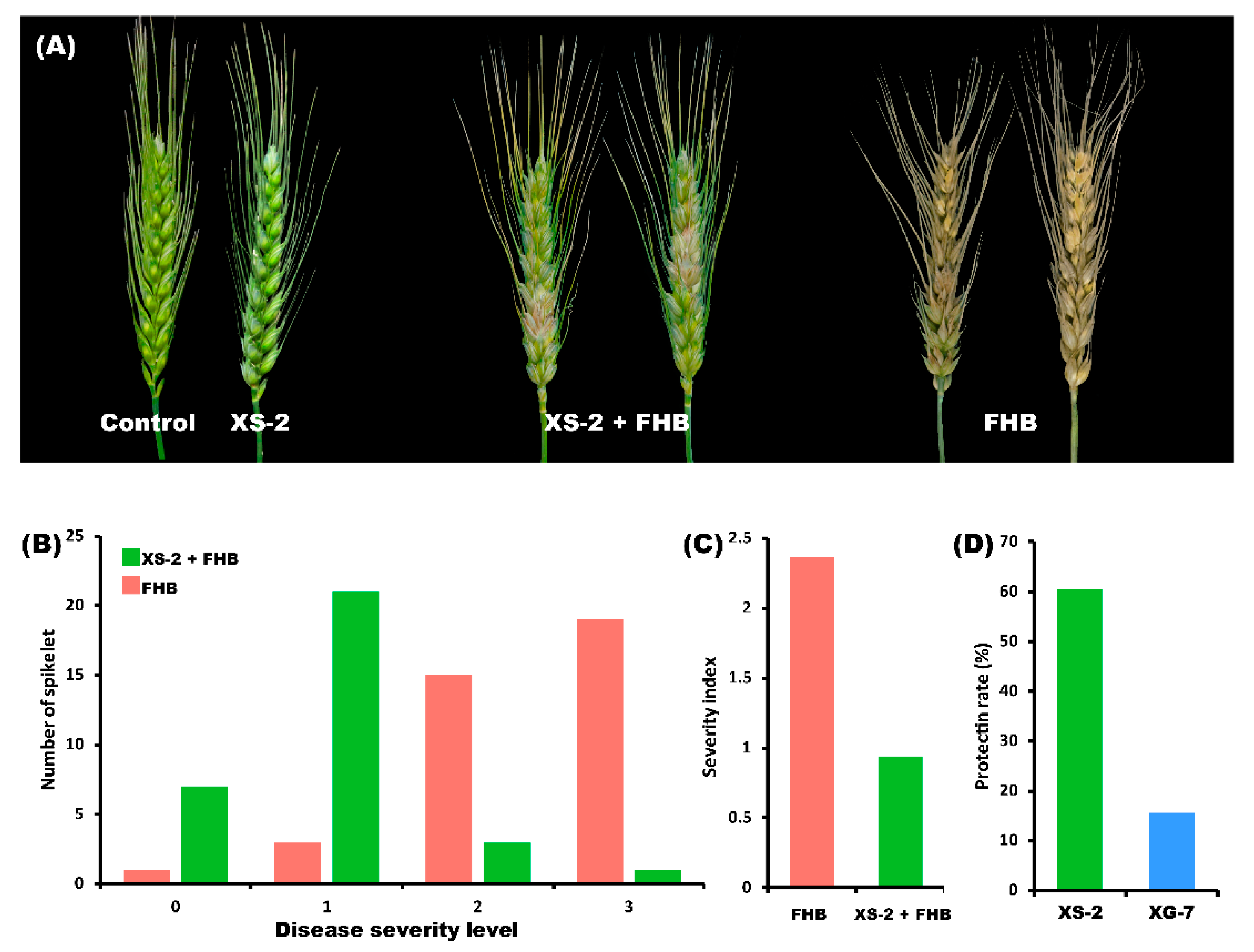
3.4. Antagonistic in Vitro Test of XS-2 on F. graminearum in Detached Spikelets
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jung, B.; Park, S.-Y.; Lee, Y.-W.; Lee, J. Biological Efficacy of Streptomyces sp. Strain BN1 against the Cereal Head Blight Pathogen Fusarium graminearum. Plant Pathol. J. 2013, 29, 52–58. [Google Scholar] [CrossRef]
- Crane, J.M.; Gibson, D.M.; Vaughan, R.H.; Bergstrom, G.C. Iturin Levels on Wheat Spikes Linked to Biological Control of Fusarium Head Blight by Bacillus amyloliquefaciens. Phytopathology 2013, 103, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Draeger, R.; Gosman, N.; Steed, A.; Chandler, E.; Thomsett, M.; Schondelmaier, J.; Buerstmayr, H.; Lemmens, M.; Schmolke, M.; Mesterhazy, A.; et al. Identification of QTLs for resistance to Fusarium head blight, DON accumulation and associated traits in the winter wheat variety Arina. Theor. Appl. Genet. 2007, 115, 617–625. [Google Scholar] [CrossRef]
- Miu, W.H. Characteristics and high yield cultivation of Sheng selected No. 6 wheat. Bull. Agric. Sci. Technol. 2010, 7, 139–140. [Google Scholar]
- Yin, J.L.; Zhang, X.; Ma, D.F.; Lu, C.; He, Y.Q.; Zhu, Y.X. A study on the sensitivity and synergistic effect of prochloraz and cypermethrin on Gibberella zeae in wheat. Acta Agric. Univ. Jiangxiensis 2018, 40, 920–924. [Google Scholar]
- Xia, Y.C.; Zhu, Y.X.; Ma, D.F.; Liu, L.C.; Yin, J.L. Isolation and identification of antagonistic strain of Fusarium graminearum. Acta Agric. Univ. Jiangxiensis 2018, 41, 33–42. [Google Scholar]
- Khan, N.I.; Schisler, D.A.; Boehm, M.J.; Slininger, P.J.; Bothast, R.J. Selection and Evaluation of Microorganisms for Biocontrol of Fusarium Head Blight of Wheat Incited by Gibberella zeae. Plant Dis. 2001, 85, 1253–1258. [Google Scholar] [CrossRef]
- Comby, M.; Gacoin, M.; Robineau, M.; Rabenoelina, F.; Ptas, S.; Dupont, J.; Profizi, C.; Baillieul, F. Screening of wheat endophytes as biological control agents against Fusarium head blight using two different in vitro tests. Microbiol. Res. 2017, 202, 11–20. [Google Scholar] [CrossRef]
- Shahzad, R.; Khan, A.L.; Bilal, S.; Waqas, M.; Kang, S.-M.; Lee, I.-J. Inoculation of abscisic acid-producing endophytic bacteria enhances salinity stress tolerance in Oryza sativa. Environ. Exp. Bot. 2017, 136, 68–77. [Google Scholar] [CrossRef]
- Mota, M.S.; Gomes, C.B.; Souza Júnior, I.T.; Moura, A.B. Bacterial selection for biological control of plant disease: Criterion determination and validation. Braz. J. Microbiol. 2017, 48, 62–70. [Google Scholar] [CrossRef]
- Bello, G.D.; Mónaco, C.; Simon, M.R. Biological control of seedling blight of wheat caused by Fusarium graminearum with beneficial rhizosphere microorganisms. World J. Microbiol. Biotechnol. 2002, 18, 627–636. [Google Scholar] [CrossRef]
- Alimi, M. Characterization and application of microbial antagonists for control of Fusarium head blight of wheat caused by Fusarium graminearum using single and mixture strain of antagonistic bacteria on resistance and susceptible cultivars. Afr. J. Microbiol. Res. 2012, 6, 326–334. [Google Scholar]
- Palazzini, J.M.; Alberione, E.; Torres, A.; Donat, C.; Köhl, J.; Chulze, S. Biological control of Fusarium graminearum sensu stricto, causal agent of Fusarium head blight of wheat, using formulated antagonists under field conditions in Argentina. Boil. Control 2016, 94, 56–61. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; McLean, J.E.; Britt, D.W.; Anderson, A.J. Antifungal activity of ZnO nanoparticles and their interactive effect with a biocontrol bacterium on growth antagonism of the plant pathogen Fusarium graminearum. BioMetals 2013, 26, 913–924. [Google Scholar] [CrossRef]
- Liu, H.F.; Zhou, J.; Liao, J.; Yi, J.P.; Ma, D.F.; Deng, J.X. Grafted twig rot on Citrus sinensis caused by a member of the Fusarium solani species complex. Can. J. Plant Pathol. 2019, 1–7. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Boil. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Malandrakis, A.A.; Markoglou, A.N.; Konstantinou, S.; Doukas, E.G.; Kalampokis, J.F.; Karaoglanidis, G.S. Molecular characterization, fitness and mycotoxin production of benzimidazole-resistant isolates of Penicillium expansum. Int. J. Food Microbiol. 2013, 162, 237–244. [Google Scholar] [CrossRef]
- Xu, J.H.; Wang, J.W.; Hu, X.D.; Ji, F.; Shi, J.R. Isolation, identification of antagonistic bacteria AF0907 against Fusarium graminearum and its characteristics. Jiangsu J. Agric. Sci. 2013, 29, 517–522. [Google Scholar]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef]
- Arrebola, E.; Sivakumar, D.; Bacigalupo, R.; Korsten, L. Combined application of antagonist Bacillus amyloliquefaciens and essential oils for the control of peach postharvest diseases. Crop. Prot. 2010, 29, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, S.; Hiradate, S.; Tsukamoto, T.; Hatakeda, K.; Shirata, A. Antimicrobial Activity of Culture Filtrate ofBacillus amyloliquefaciensRC-2 Isolated from Mulberry Leaves. Phytopathology 2001, 91, 181–187. [Google Scholar] [CrossRef]
- Fan, B.; Blom, J.; Klenk, H.-P.; Borriss, R. Bacillus amyloliquefaciens, Bacillus velezensis, and Bacillus siamensis Form an “Operational Group B. amyloliquefaciens” within the B. subtilis Species Complex. Front. Microbiol. 2017, 8, 1807. [Google Scholar] [CrossRef]
- Porter, J.R.; Gawith, M. Temperatures and the growth and development of wheat: A review. Eur. J. Agron. 1999, 10, 23–36. [Google Scholar] [CrossRef]
- Asseng, S.; Foster, I.; Turner, N.C. The impact of temperature variability on wheat yields. Glob. Chang. Boil. 2011, 17, 997–1012. [Google Scholar] [CrossRef]
- Collins, D.P.; Jacobsen, B.J. Optimizing a Bacillus subtilis isolate for biological control of sugar beet cercospora leaf spot. Boil. Control 2003, 26, 153–161. [Google Scholar] [CrossRef]
- Chiou, A.L.; Wu, W.S. Formulation of Bacillus amyloliquefaciens B190 for Control of Lily Grey Mould (Botrytis elliptica). J. Phytopathol. 2003, 151, 13–18. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen | Average Inhibition Zone Diameter (mm) |
|---|---|
| Colletotrichum gloeosporioides | 22.0 ± 6.8 d |
| Rhizoctonia solani | 25.4 ± 6.7 c |
| Sclerotium rolfsii | 29.6 ± 1.7 b |
| Alternaria alternata | 33.3 ± 0.2 a |
| Fusarium moniliforme | 22.3 ± 0.5 c,d |
| Colletotrichum truncatum | 23.4 ± 1.1 c,d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Zhu, Y.; Ma, D.; Xu, W.; Zhou, J.; Yan, H.; Yang, L.; Yin, J. Screening, Identification, and Optimization of Fermentation Conditions of an Antagonistic Endophyte to Wheat Head Blight. Agronomy 2019, 9, 476. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090476
Zhang P, Zhu Y, Ma D, Xu W, Zhou J, Yan H, Yang L, Yin J. Screening, Identification, and Optimization of Fermentation Conditions of an Antagonistic Endophyte to Wheat Head Blight. Agronomy. 2019; 9(9):476. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090476
Chicago/Turabian StyleZhang, Peigen, Yongxing Zhu, Dongfang Ma, Wenjie Xu, Jingjing Zhou, Hanwen Yan, Lei Yang, and Junliang Yin. 2019. "Screening, Identification, and Optimization of Fermentation Conditions of an Antagonistic Endophyte to Wheat Head Blight" Agronomy 9, no. 9: 476. https://0-doi-org.brum.beds.ac.uk/10.3390/agronomy9090476