
Neutrophil Extracellular Traps Contribute to COVID-19 Hyperinflammation and Humoral Autoimmunity

 , , , ,
, , , ,  , and
, and
Abstract
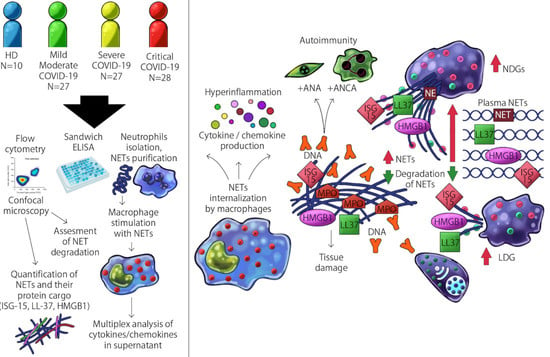
:
1. Introduction
2. Materials and Methods
- Mild/moderate disease: Fever, upper respiratory infection symptoms, with or without pneumonia.
- Severe: Any of the following: respiratory failure, respiratory rate ≥ 30 breaths per minute, oxygen saturation at rest ≤ 93%, PaO2/FiO2 ≤ 300 mmHg.
- Critical: Any of the following: need for invasive mechanical ventilation (IMV), shock, multiple organ failure.
2.1. Assessment of the Proportion of LDG by Multiparametric Flow-Cytometry
2.2. Assessment of Plasmatic Circulating NETs and Their Protein Cargo
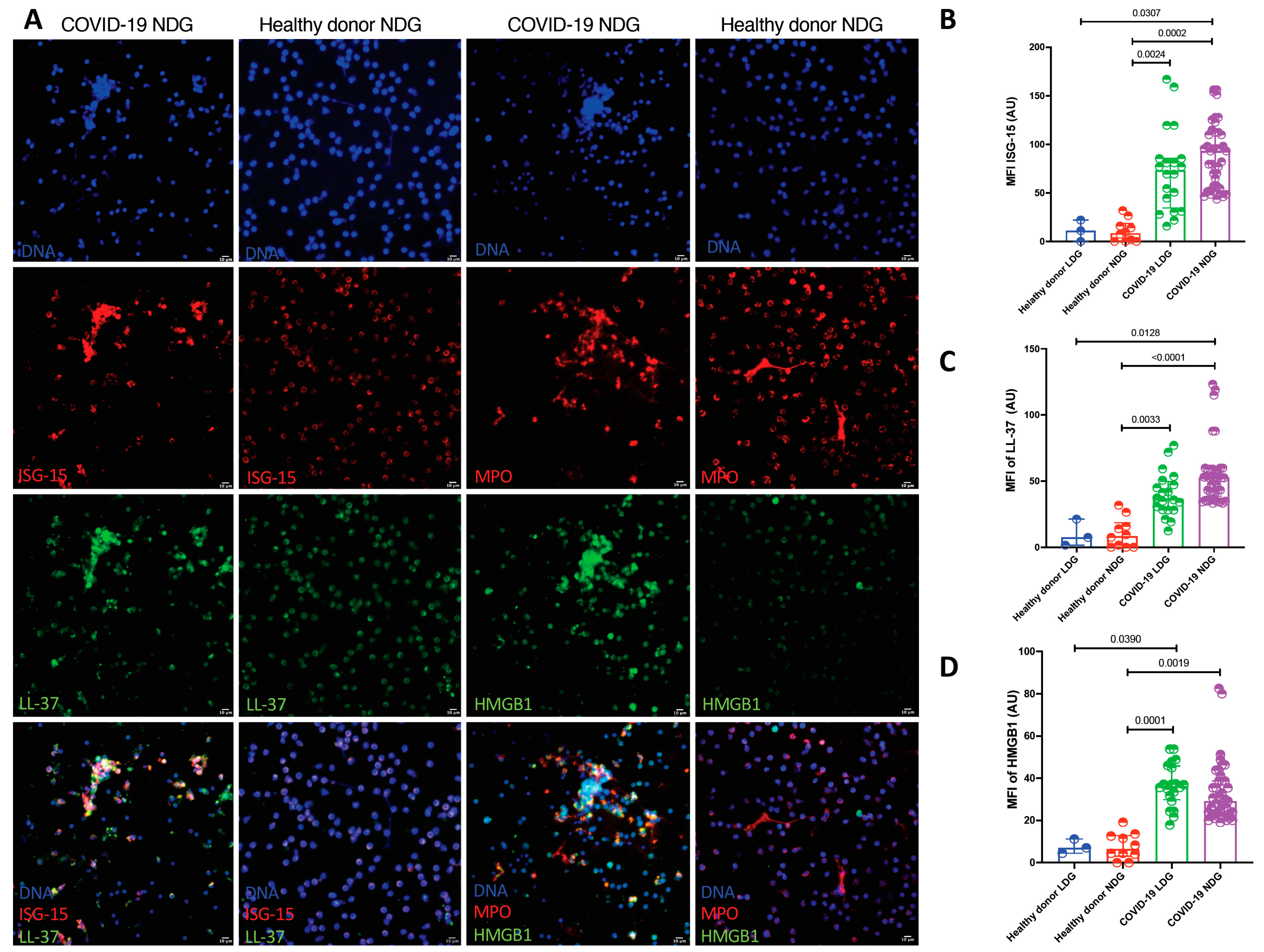
2.3. Quantification of the Amount of LDG and NDG Derived NETs and Their Protein Cargo by Confocal Microscopy
2.4. Obtention of NETs from LDG and NDG
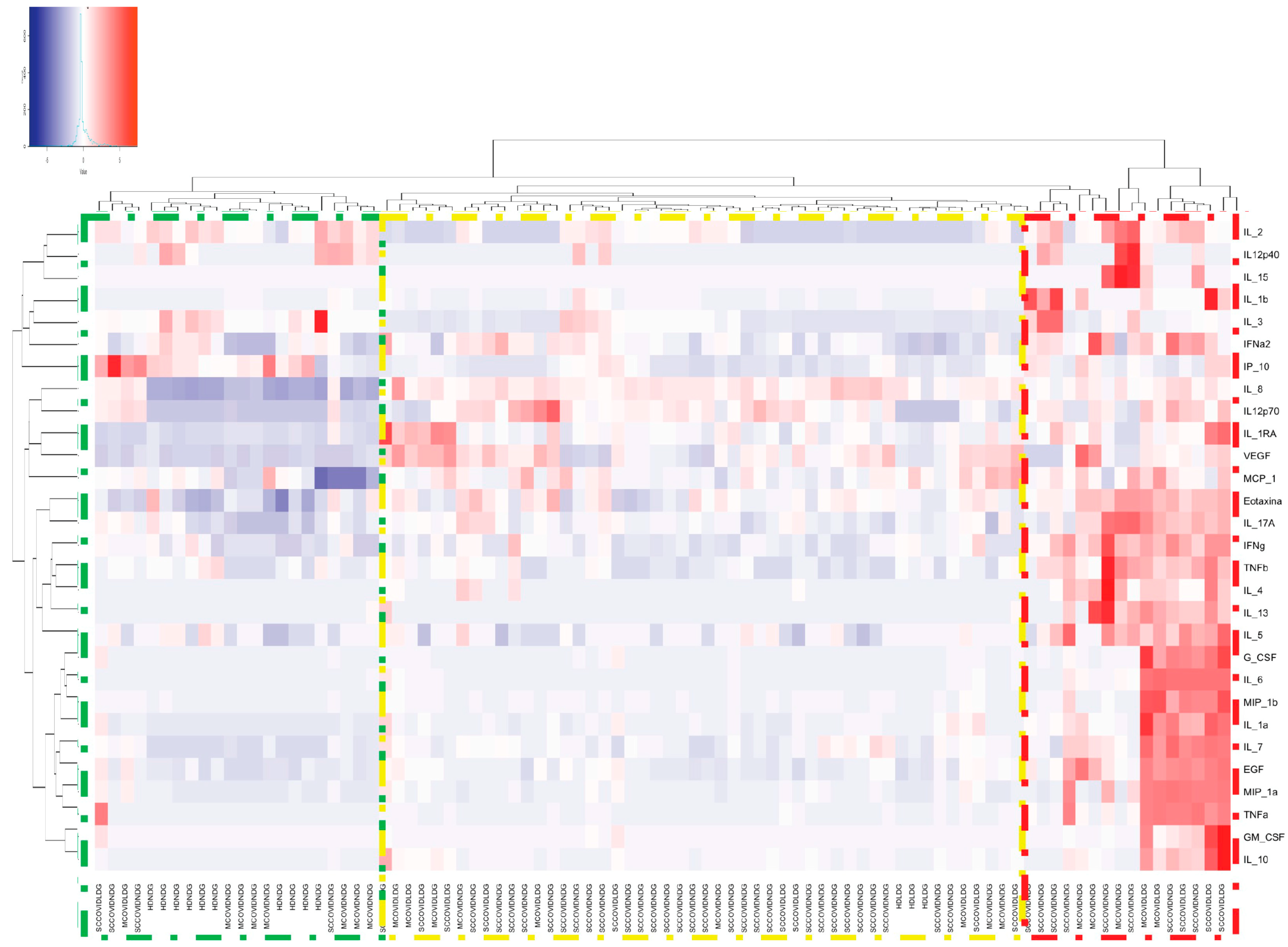
2.5. Stimulation of Monocyte-Derived Macrophages with Neutrophil Extracellular Traps and Assessment of Cytokine/Chemokine Production
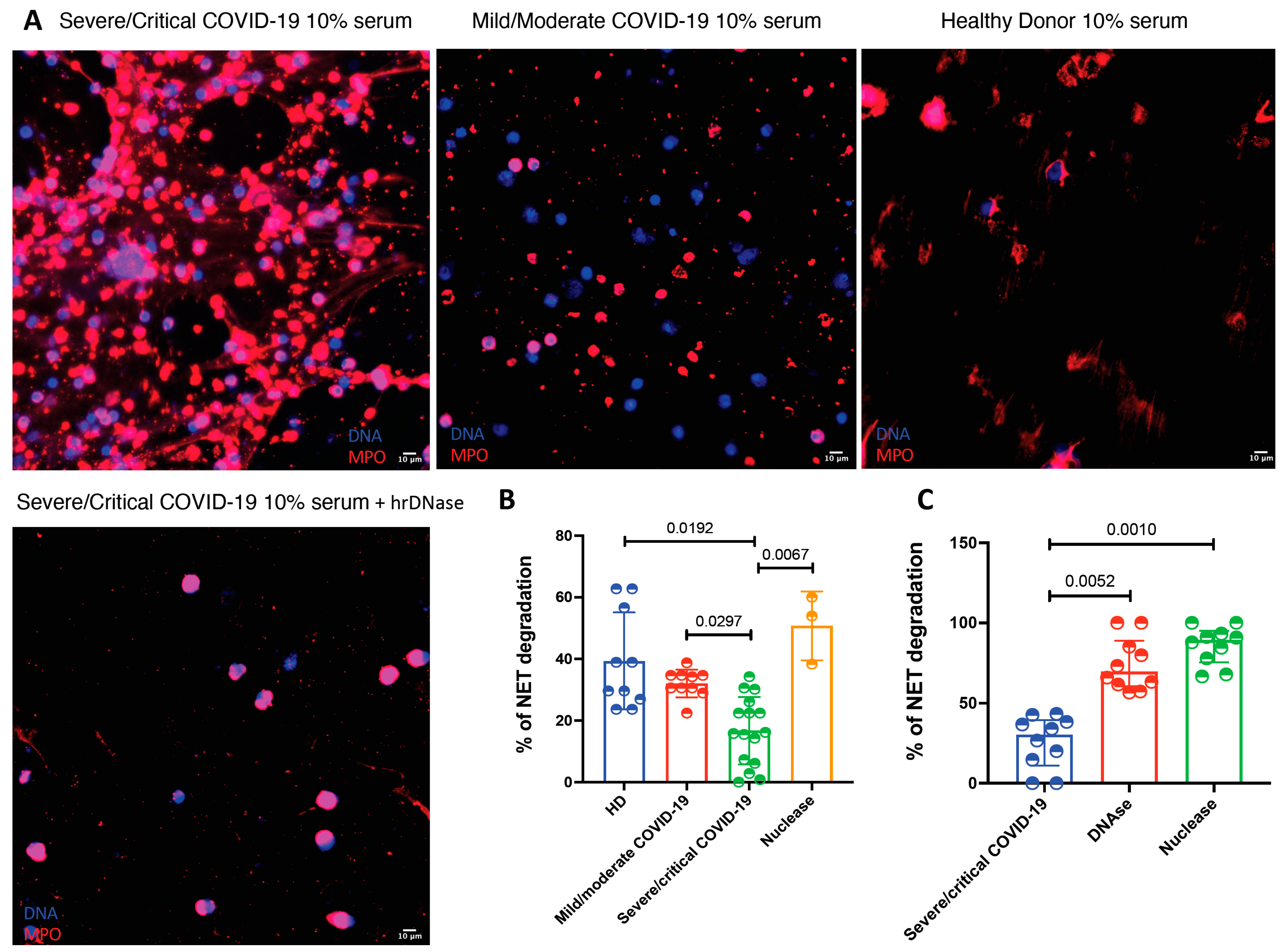
2.6. Assessment of the Serum Degradation of NETs
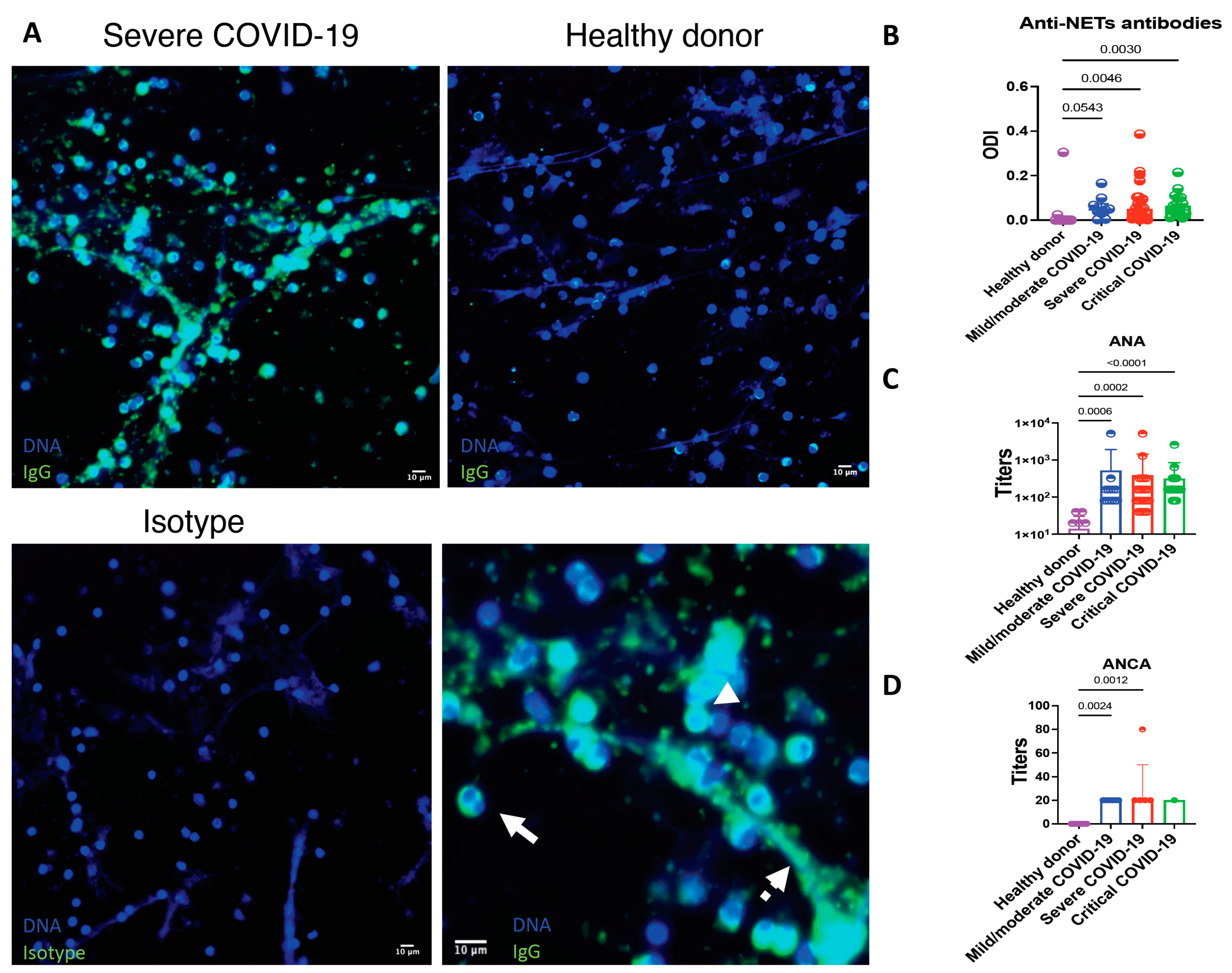
2.7. Appraisal of Anti-NET Antibodies, Antinuclear Antibodies (ANA) and Anti-Neutrophil Cytoplasmic Antibodies (ANCA)
3. Statistical Analysis
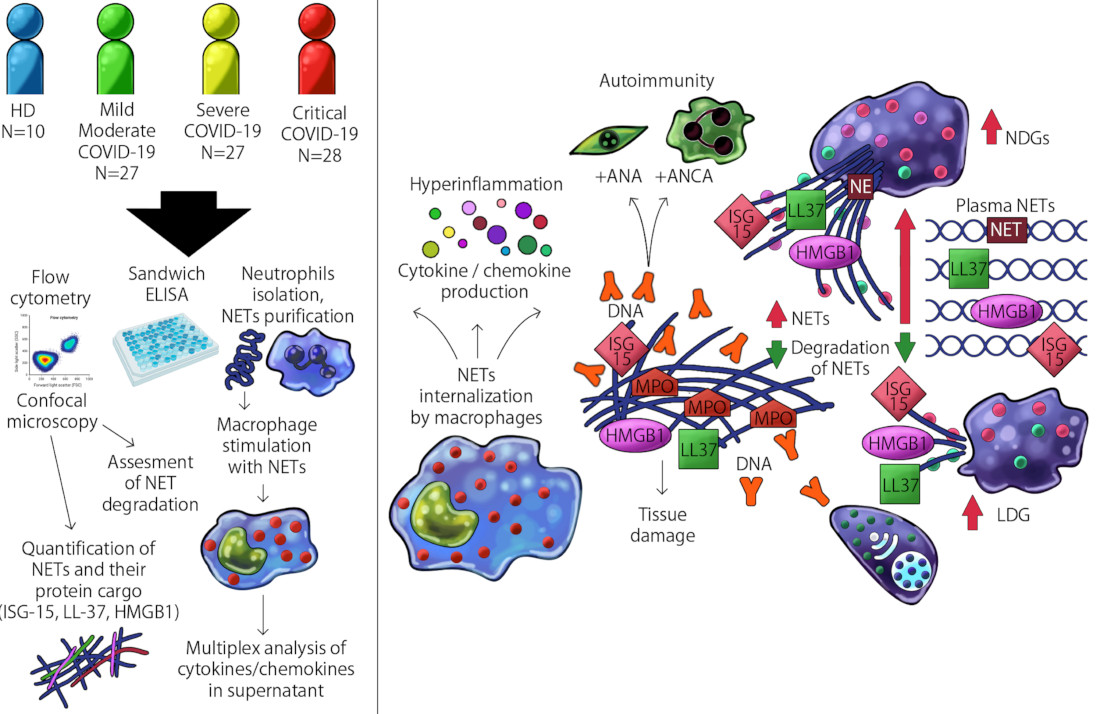
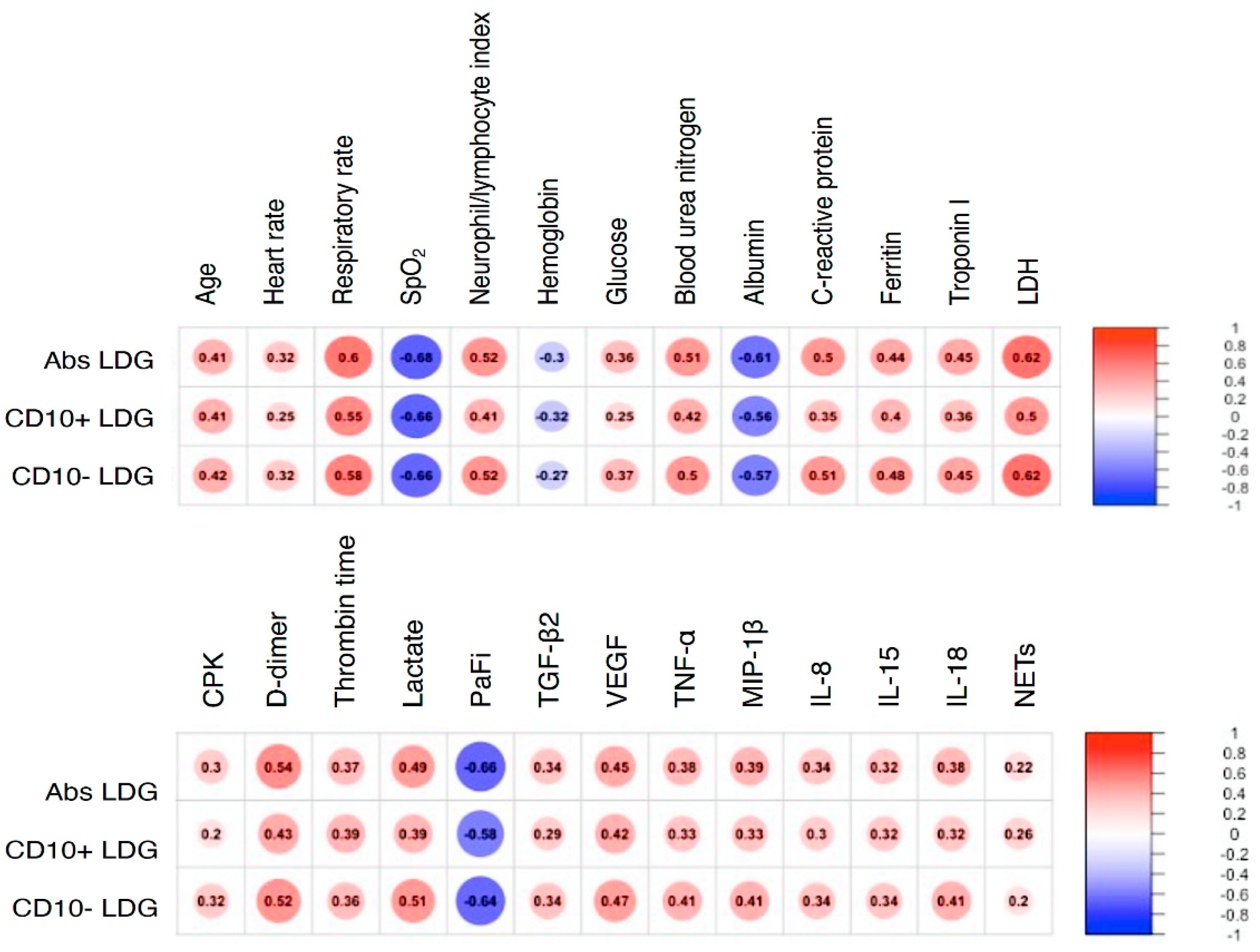
4. Results
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saad, M.A.; Alfishawy, M.; Nassar, M.; Mohamed, M.; Esene, I.N.; Elbendary, A. COVID-19 and Autoimmune Diseases: A Systematic Review of Reported Cases. Curr. Rheumatol. Rev. 2021, 17, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, Y.; Novelli, L.; Rojas, M.; De Santis, M.; Acosta-Ampudia, Y.; Monsalve, D.M.; Ramirez-Santana, C.; Costanzo, A.; Ridgway, W.M.; Ansari, A.A.; et al. Autoinflammatory and autoimmune conditions at the crossroad of COVID-19. J. Autoimmun. 2020, 114, 102506. [Google Scholar] [CrossRef]
- Mantovani, A.; Netea, M.G. Trained Innate Immunity, Epigenetics, and Covid-19. N. Engl. J. Med. 2020, 383, 1078–1080. [Google Scholar] [CrossRef] [PubMed]
- Shaath, H.; Vishnubalaji, R.; Elkord, E.; Alajez, N.M. Single-Cell Transcriptome Analysis Highlights a Role for Neutrophils and Inflammatory Macrophages in the Pathogenesis of Severe COVID-19. Cells 2020, 9, 2374. [Google Scholar] [CrossRef] [PubMed]
- Garcia, L.F. Immune Response, Inflammation, and the Clinical Spectrum of COVID-19. Front. Immunol. 2020, 11, 1441. [Google Scholar] [CrossRef] [PubMed]
- Gardinassi, L.G.; Souza, C.O.S.; Sales-Campos, H.; Fonseca, S.G. Immune and Metabolic Signatures of COVID-19 Revealed by Transcriptomics Data Reuse. Front. Immunol. 2020, 11, 1636. [Google Scholar] [CrossRef]
- Banchereau, R.; Hong, S.; Cantarel, B.; Baldwin, N.; Baisch, J.; Edens, M.; Cepika, A.M.; Acs, P.; Turner, J.; Anguiano, E.; et al. Personalized Immunomonitoring Uncovers Molecular Networks that Stratify Lupus Patients. Cell 2016, 165, 1548–1550. [Google Scholar] [CrossRef] [PubMed]
- Onodi, F.; Bonnet-Madin, L.; Meertens, L.; Karpf, L.; Poirot, J.; Zhang, S.Y.; Picard, C.; Puel, A.; Jouanguy, E.; Zhang, Q.; et al. SARS-CoV-2 induces human plasmacytoid predendritic cell diversification via UNC93B and IRAK4. J. Exp. Med. 2021, 218. [Google Scholar] [CrossRef]
- McGonagle, D.; Sharif, K.; O’Regan, A.; Bridgewood, C. The Role of Cytokines including Interleukin-6 in COVID-19 induced Pneumonia and Macrophage Activation Syndrome-Like Disease. Autoimmun. Rev. 2020, 19, 102537. [Google Scholar] [CrossRef]
- Kaplan, M.J.; Radic, M. Neutrophil extracellular traps: Double-edged swords of innate immunity. J. Immunol. 2012, 189, 2689–2695. [Google Scholar] [CrossRef] [Green Version]
- Denny, M.F.; Yalavarthi, S.; Zhao, W.; Thacker, S.G.; Anderson, M.; Sandy, A.R.; McCune, W.J.; Kaplan, M.J. A distinct subset of proinflammatory neutrophils isolated from patients with systemic lupus erythematosus induces vascular damage and synthesizes type I IFNs. J. Immunol. 2010, 184, 3284–3297. [Google Scholar] [CrossRef] [Green Version]
- Hakkim, A.; Furnrohr, B.G.; Amann, K.; Laube, B.; Abed, U.A.; Brinkmann, V.; Herrmann, M.; Voll, R.E.; Zychlinsky, A. Impairment of neutrophil extracellular trap degradation is associated with lupus nephritis. Proc. Natl. Acad. Sci. USA 2010, 107, 9813–9818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahajan, A.; Herrmann, M.; Munoz, L.E. Clearance Deficiency and Cell Death Pathways: A Model for the Pathogenesis of SLE. Front. Immunol. 2016, 7, 35. [Google Scholar] [CrossRef] [Green Version]
- Daniel, C.; Leppkes, M.; Munoz, L.E.; Schley, G.; Schett, G.; Herrmann, M. Extracellular DNA traps in inflammation, injury and healing. Nat. Rev. Nephrol. 2019, 15, 559–575. [Google Scholar] [CrossRef]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhou, X.; Tan, H.; Hu, Y.; Zhang, L.; Liu, S.; Dai, M.; Li, Y.; Li, Q.; Mao, Z.; et al. Neutrophil extracellular traps contribute to the pathogenesis of acid-aspiration-induced ALI/ARDS. Oncotarget 2018, 9, 1772–1784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, Y.; Yalavarthi, S.; Shi, H.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.; Weber, A.; Barnes, B.J.; Egeblad, M.; et al. Neutrophil extracellular traps in COVID-19. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [Green Version]
- Middleton, E.A.; He, X.Y.; Denorme, F.; Campbell, R.A.; Ng, D.; Salvatore, S.P.; Mostyka, M.; Baxter-Stoltzfus, A.; Borczuk, A.C.; Loda, M.; et al. Neutrophil extracellular traps contribute to immunothrombosis in COVID-19 acute respiratory distress syndrome. Blood 2020, 136, 1169–1179. [Google Scholar] [CrossRef]
- Andersson, U.; Ottestad, W.; Tracey, K.J. Extracellular HMGB1: A therapeutic target in severe pulmonary inflammation including COVID-19? Mol. Med. 2020, 26, 42. [Google Scholar] [CrossRef]
- Thierry, A.R.; Roch, B. SARS-CoV2 may evade innate immune response, causing uncontrolled neutrophil extracellular traps formation and multi-organ failure. Clin. Sci. (Lond.) 2020, 134, 1295–1300. [Google Scholar] [CrossRef]
- Woodruff, M.C.; Ramonell, R.P.; Nguyen, D.C.; Cashman, K.S.; Saini, A.S.; Haddad, N.S.; Ley, A.M.; Kyu, S.; Howell, J.C.; Ozturk, T.; et al. Extrafollicular B cell responses correlate with neutralizing antibodies and morbidity in COVID-19. Nat. Immunol. 2020, 21, 1506–1516. [Google Scholar] [CrossRef]
- Mathew, D.; Giles, J.R.; Baxter, A.E.; Oldridge, D.A.; Greenplate, A.R.; Wu, J.E.; Alanio, C.; Kuri-Cervantes, L.; Pampena, M.B.; D’Andrea, K.; et al. Deep immune profiling of COVID-19 patients reveals distinct immunotypes with therapeutic implications. Science 2020, 369, eabc8511. [Google Scholar] [CrossRef] [PubMed]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Beziat, V.; et al. Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science 2020, 370. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, M.C.; Ramonell, R.P.; Lee, F.E.; Sanz, I. Broadly-targeted autoreactivity is common in severe SARS-CoV-2 Infection. medRxiv 2020. [Google Scholar] [CrossRef]
- Liu, W.; Tao, Z.W.; Wang, L.; Yuan, M.L.; Liu, K.; Zhou, L.; Wei, S.; Deng, Y.; Liu, J.; Liu, H.G.; et al. Analysis of factors associated with disease outcomes in hospitalized patients with 2019 novel coronavirus disease. Chin. Med. J. (Engl.) 2020, 133, 1032–1038. [Google Scholar] [CrossRef] [PubMed]
- Lood, C.; Blanco, L.P.; Purmalek, M.M.; Carmona-Rivera, C.; De Ravin, S.S.; Smith, C.K.; Malech, H.L.; Ledbetter, J.A.; Elkon, K.B.; Kaplan, M.J. Neutrophil extracellular traps enriched in oxidized mitochondrial DNA are interferogenic and contribute to lupus-like disease. Nat. Med. 2016, 22, 146–153. [Google Scholar] [CrossRef] [Green Version]
- O’Neil, L.J.; Barrera-Vargas, A.; Sandoval-Heglund, D.; Merayo-Chalico, J.; Aguirre-Aguilar, E.; Aponte, A.M.; Ruiz-Perdomo, Y.; Gucek, M.; El-Gabalawy, H.; Fox, D.A.; et al. Neutrophil-mediated carbamylation promotes articular damage in rheumatoid arthritis. Sci. Adv. 2020, 6, eabd2688. [Google Scholar] [CrossRef]
- Carmona-Rivera, C.; Kaplan, M.J. Induction and Quantification of NETosis. Curr. Protoc. Immunol. 2016, 115, 11–14. [Google Scholar] [CrossRef]
- Veras, F.P.; Pontelli, M.C.; Silva, C.M.; Toller-Kawahisa, J.E.; de Lima, M.; Nascimento, D.C.; Schneider, A.H.; Caetite, D.; Tavares, L.A.; Paiva, I.M.; et al. SARS-CoV-2-triggered neutrophil extracellular traps mediate COVID-19 pathology. J. Exp. Med. 2020, 217, e20201129. [Google Scholar] [CrossRef]
- Rocha, B.C.; Marques, P.E.; Leoratti, F.M.S.; Junqueira, C.; Pereira, D.B.; Antonelli, L.; Menezes, G.B.; Golenbock, D.T.; Gazzinelli, R.T. Type I Interferon Transcriptional Signature in Neutrophils and Low-Density Granulocytes Are Associated with Tissue Damage in Malaria. Cell Rep. 2015, 13, 2829–2841. [Google Scholar] [CrossRef] [Green Version]
- Izquierdo, E.; Cuevas, V.D.; Fernandez-Arroyo, S.; Riera-Borrull, M.; Orta-Zavalza, E.; Joven, J.; Rial, E.; Corbi, A.L.; Escribese, M.M. Reshaping of Human Macrophage Polarization through Modulation of Glucose Catabolic Pathways. J. Immunol. 2015, 195, 2442–2451. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Yalavarthi, S.; Gockman, K.; Madison, J.A.; Gudjonsson, J.E.; Kahlenberg, J.M.; Joseph McCune, W.; Bockenstedt, P.L.; Karp, D.R.; Knight, J.S. Anti-Neutrophil Extracellular Trap Antibodies and Impaired Neutrophil Extracellular Trap Degradation in Antiphospholipid Syndrome. Arthritis Rheumatol. 2020, 72, 2130–2135. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Rivera, C.; Kaplan, M.J. Detection of SLE antigens in neutrophil extracellular traps (NETs). Methods Mol. Biol. 2014, 1134, 151–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agmon-Levin, N.; Damoiseaux, J.; Kallenberg, C.; Sack, U.; Witte, T.; Herold, M.; Bossuyt, X.; Musset, L.; Cervera, R.; Plaza-Lopez, A.; et al. International recommendations for the assessment of autoantibodies to cellular antigens referred to as anti-nuclear antibodies. Ann. Rheum. Dis. 2014, 73, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Leppkes, M.; Knopf, J.; Naschberger, E.; Lindemann, A.; Singh, J.; Herrmann, I.; Sturzl, M.; Staats, L.; Mahajan, A.; Schauer, C.; et al. Vascular occlusion by neutrophil extracellular traps in COVID-19. EBioMedicine 2020, 58, 102925. [Google Scholar] [CrossRef]
- Vitte, J.; Diallo, A.B.; Boumaza, A.; Lopez, A.; Michel, M.; Allardet-Servent, J.; Mezouar, S.; Sereme, Y.; Busnel, J.M.; Miloud, T.; et al. A Granulocytic Signature Identifies COVID-19 and Its Severity. J. Infect. Dis. 2020, 222, 1985–1996. [Google Scholar] [CrossRef] [PubMed]
- Mann, E.R.; Menon, M.; Knight, S.B.; Konkel, J.E.; Jagger, C.; Shaw, T.N.; Krishnan, S.; Rattray, M.; Ustianowski, A.; Bakerly, N.D.; et al. Longitudinal immune profiling reveals key myeloid signatures associated with COVID-19. Sci. Immunol. 2020, 5, eabd6197. [Google Scholar] [CrossRef] [PubMed]
- Parackova, Z.; Zentsova, I.; Bloomfield, M.; Vrabcova, P.; Smetanova, J.; Klocperk, A.; Meseznikov, G.; Casas Mendez, L.F.; Vymazal, T.; Sediva, A. Disharmonic Inflammatory Signatures in COVID-19: Augmented Neutrophils’ but Impaired Monocytes’ and Dendritic Cells’ Responsiveness. Cells 2020, 9, 2206. [Google Scholar] [CrossRef]
- Mistry, P.; Nakabo, S.; O’Neil, L.; Goel, R.R.; Jiang, K.; Carmona-Rivera, C.; Gupta, S.; Chan, D.W.; Carlucci, P.M.; Wang, X.; et al. Transcriptomic, epigenetic, and functional analyses implicate neutrophil diversity in the pathogenesis of systemic lupus erythematosus. Proc. Natl. Acad. Sci. USA 2019, 116, 25222–25228. [Google Scholar] [CrossRef]
- Korakas, E.; Ikonomidis, I.; Kousathana, F.; Balampanis, K.; Kountouri, A.; Raptis, A.; Palaiodimou, L.; Kokkinos, A.; Lambadiari, V. Obesity and COVID-19: Immune and metabolic derangement as a possible link to adverse clinical outcomes. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E105–E109. [Google Scholar] [CrossRef]
- Guo, C.; Li, B.; Ma, H.; Wang, X.; Cai, P.; Yu, Q.; Zhu, L.; Jin, L.; Jiang, C.; Fang, J.; et al. Single-cell analysis of two severe COVID-19 patients reveals a monocyte-associated and tocilizumab-responding cytokine storm. Nat. Commun. 2020, 11, 3924. [Google Scholar] [CrossRef]
- Arunachalam, P.S.; Wimmers, F.; Mok, C.K.P.; Perera, R.; Scott, M.; Hagan, T.; Sigal, N.; Feng, Y.; Bristow, L.; Tak-Yin Tsang, O.; et al. Systems biological assessment of immunity to mild versus severe COVID-19 infection in humans. Science 2020, 369, 1210–1220. [Google Scholar] [CrossRef]
- Chakraborty, S.; Gonzalez, J.; Edwards, K.; Mallajosyula, V.; Buzzanco, A.S.; Sherwood, R.; Buffone, C.; Kathale, N.; Providenza, S.; Xie, M.M.; et al. Proinflammatory IgG Fc structures in patients with severe COVID-19. Nat. Immunol. 2021, 22, 67–73. [Google Scholar] [CrossRef]
- Radermecker, C.; Detrembleur, N.; Guiot, J.; Cavalier, E.; Henket, M.; d’Emal, C.; Vanwinge, C.; Cataldo, D.; Oury, C.; Delvenne, P.; et al. Neutrophil extracellular traps infiltrate the lung airway, interstitial, and vascular compartments in severe COVID-19. J. Exp. Med. 2020, 217, e20201012. [Google Scholar] [CrossRef] [PubMed]
- Cortjens, B.; de Boer, O.J.; de Jong, R.; Antonis, A.F.; Sabogal Pineros, Y.S.; Lutter, R.; van Woensel, J.B.; Bem, R.A. Neutrophil extracellular traps cause airway obstruction during respiratory syncytial virus disease. J. Pathol. 2016, 238, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Magro, C.; Mulvey, J.J.; Berlin, D.; Nuovo, G.; Salvatore, S.; Harp, J.; Baxter-Stoltzfus, A.; Laurence, J. Complement associated microvascular injury and thrombosis in the pathogenesis of severe COVID-19 infection: A report of five cases. Transl. Res. 2020, 220, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Carvelli, J.; Demaria, O.; Vely, F.; Batista, L.; Chouaki Benmansour, N.; Fares, J.; Carpentier, S.; Thibult, M.L.; Morel, A.; Remark, R.; et al. Association of COVID-19 inflammation with activation of the C5a-C5aR1 axis. Nature 2020, 588, 146–150. [Google Scholar] [CrossRef]
- Skendros, P.; Mitsios, A.; Chrysanthopoulou, A.; Mastellos, D.C.; Metallidis, S.; Rafailidis, P.; Ntinopoulou, M.; Sertaridou, E.; Tsironidou, V.; Tsigalou, C.; et al. Complement and tissue factor-enriched neutrophil extracellular traps are key drivers in COVID-19 immunothrombosis. J. Clin. Investig. 2020, 130, 6151–6157. [Google Scholar] [CrossRef]
- Xu, G.; Qi, F.; Li, H.; Yang, Q.; Wang, H.; Wang, X.; Liu, X.; Zhao, J.; Liao, X.; Liu, Y.; et al. The differential immune responses to COVID-19 in peripheral and lung revealed by single-cell RNA sequencing. Cell Discov. 2020, 6, 73. [Google Scholar] [CrossRef]
- Perng, Y.C.; Lenschow, D.J. ISG15 in antiviral immunity and beyond. Nat. Rev. Microbiol. 2018, 16, 423–439. [Google Scholar] [CrossRef]
- Hadjadj, J.; Yatim, N.; Barnabei, L.; Corneau, A.; Boussier, J.; Smith, N.; Pere, H.; Charbit, B.; Bondet, V.; Chenevier-Gobeaux, C.; et al. Impaired type I interferon activity and inflammatory responses in severe COVID-19 patients. Science 2020, 369, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Barlow, P.G.; Svoboda, P.; Mackellar, A.; Nash, A.A.; York, I.A.; Pohl, J.; Davidson, D.J.; Donis, R.O. Antiviral activity and increased host defense against influenza infection elicited by the human cathelicidin LL-37. PLoS ONE 2011, 6, e25333. [Google Scholar] [CrossRef] [PubMed]
- Currie, S.M.; Findlay, E.G.; McHugh, B.J.; Mackellar, A.; Man, T.; Macmillan, D.; Wang, H.; Fitch, P.M.; Schwarze, J.; Davidson, D.J. The human cathelicidin LL-37 has antiviral activity against respiratory syncytial virus. PLoS ONE 2013, 8, e73659. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Adhikarakunnathu, S.; Bhardwaj, K.; Ranjith-Kumar, C.T.; Wen, Y.; Jordan, J.L.; Wu, L.H.; Dragnea, B.; San Mateo, L.; Kao, C.C. LL37 and cationic peptides enhance TLR3 signaling by viral double-stranded RNAs. PLoS ONE 2011, 6, e26632. [Google Scholar] [CrossRef] [Green Version]
- Cicco, S.; Cicco, G.; Racanelli, V.; Vacca, A. Neutrophil Extracellular Traps (NETs) and Damage-Associated Molecular Patterns (DAMPs): Two Potential Targets for COVID-19 Treatment. Mediators Inflamm. 2020, 2020, 7527953. [Google Scholar] [CrossRef]
- Wei, J.; Alfajaro, M.M.; DeWeirdt, P.C.; Hanna, R.E.; Lu-Culligan, W.J.; Cai, W.L.; Strine, M.S.; Zhang, S.M.; Graziano, V.R.; Schmitz, C.O.; et al. Genome-wide CRISPR Screens Reveal Host Factors Critical for SARS-CoV-2 Infection. Cell 2021, 184, 76–91.e13. [Google Scholar] [CrossRef]
- Nass, K.; Frenkel, G.D. Adenovirus-induced inhibition of cellular DNase. J. Virol. 1978, 26, 540–543. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.Y.; Park, H.H.; Park, W.; Kim, H.; Jang, J.G.; Hong, K.S.; Lee, J.Y.; Seo, H.S.; Na, D.H.; Kim, T.H.; et al. Long-acting nanoparticulate DNase-1 for effective suppression of SARS-CoV-2-mediated neutrophil activities and cytokine storm. Biomaterials 2021, 267, 120389. [Google Scholar] [CrossRef]
- Okur, H.K.; Yalcin, K.; Tastan, C.; Demir, S.; Yurtsever, B.; Karakus, G.S.; Kancagi, D.D.; Abanuz, S.; Seyis, U.; Zengin, R.; et al. Preliminary report of in vitro and in vivo effectiveness of dornase alfa on SARS-CoV-2 infection. New Microbes New Infect 2020, 37, 100756. [Google Scholar] [CrossRef]
- Narasaraju, T.; Yang, E.; Samy, R.P.; Ng, H.H.; Poh, W.P.; Liew, A.A.; Phoon, M.C.; van Rooijen, N.; Chow, V.T. Excessive neutrophils and neutrophil extracellular traps contribute to acute lung injury of influenza pneumonitis. Am. J. Pathol. 2011, 179, 199–210. [Google Scholar] [CrossRef]
- Liao, M.; Liu, Y.; Yuan, J.; Wen, Y.; Xu, G.; Zhao, J.; Cheng, L.; Li, J.; Wang, X.; Wang, F.; et al. Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nat. Med. 2020, 26, 842–844. [Google Scholar] [CrossRef]
- Vojdani, A.; Kharrazian, D. Potential antigenic cross-reactivity between SARS-CoV-2 and human tissue with a possible link to an increase in autoimmune diseases. Clin. Immunol. 2020, 217, 108480. [Google Scholar] [CrossRef] [PubMed]
- DiNardo, A.R.; Netea, M.G.; Musher, D.M. Postinfectious Epigenetic Immune Modifications—A Double-Edged Sword. N. Engl. J. Med. 2021, 384, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Pascolini, S.; Vannini, A.; Deleonardi, G.; Ciordinik, M.; Sensoli, A.; Carletti, I.; Veronesi, L.; Ricci, C.; Pronesti, A.; Mazzanti, L.; et al. COVID-19 and Immunological Dysregulation: Can Autoantibodies be Useful? Clin. Transl. Sci. 2020, 14, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.Y.; Mao, T.; Klein, J.; Dai, Y.; Huck, J.D.; Liu, F.; Zheng, N.S.; Zhou, T.; Israelow, B.; Wong, P.; et al. Diverse Functional Autoantibodies in Patients with COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Zhou, Y.; Han, T.; Chen, J.; Hou, C.; Hua, L.; He, S.; Guo, Y.; Zhang, S.; Wang, Y.; Yuan, J.; et al. Clinical and Autoimmune Characteristics of Severe and Critical Cases of COVID-19. Clin. Transl. Sci. 2020, 13, 1077–1086. [Google Scholar] [CrossRef]
- Vlachoyiannopoulos, P.G.; Magira, E.; Alexopoulos, H.; Jahaj, E.; Theophilopoulou, K.; Kotanidou, A.; Tzioufas, A.G. Autoantibodies related to systemic autoimmune rheumatic diseases in severely ill patients with COVID-19. Ann. Rheum. Dis. 2020, 79, 1661–1663. [Google Scholar] [CrossRef] [PubMed]
- Beyer, C.; Stearns, N.A.; Giessl, A.; Distler, J.H.; Schett, G.; Pisetsky, D.S. The extracellular release of DNA and HMGB1 from Jurkat T cells during in vitro necrotic cell death. Innate Immun. 2012, 18, 727–737. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Healthy Donors Median (IQR) n = 10 | Mild/Moderate Median (IQR) n = 27 | Severe Median (IQR) n = 27 | Critical Median (IQR) n = 28 | p-Value |
|---|---|---|---|---|---|
| Low Density Granulocytes (LDG) | |||||
| Total LDG (%) | 0.46 (0.31–0.68) | 0.59 (0.34–1.23) | 2.35 (0.97–4.43) | 12.75 (3.19–18.43) | <0.001 |
| Total LDG (cells/mm3) | 23 (15.75–34.13) | 42.33 (19.1–99.79) | 181.00 (55.29–307.20) | 1109 (332.10–4080.00) | <0.001 |
| CD10 + LDG (%) | 0.13 (0.05–0.22) | 0.05 (0.01–0.10) | 0.20 (0.07–0.21) | 0.156 (0.30–12.63) | <0.001 |
| CD10 + LDG (cells/mm3) | 0.02 (0.01–0.06) | 0.02 (0.00–0.03) | 0.39 (0.05–5.06) | 18.98 (1.18–323.50) | <0.001 |
| CD10 − LDG (%) | 0.31 (0.17–0.46) | 0.52 (0.31–1.00) | 1.65 (0.61–3.32) | 5.98 (2.22–11.53) | <0.001 |
| CD10 − LDG (cells/mm3) | 0.07 (0.02–0.15) | 0.22 (0.05–0.84) | 2.54 (0.45–6.64) | 106.5 (7.2–225.7) | <0.001 |
| Circulating Plasmatic NETs | |||||
| HMGB1-DNA complexes (ODI) | 0.5 (0.35–0.55) | 0.45 (0.32–0.62) | 0.59 (0.44–0.87) | 1.57 (0.98–2.49) | <0.001 |
| ISG-15-DNA complexes (ODI) | 0.50 (0.40–0.53) | 0.69 (0.58–1.07) | 0.61 (0.47–0.76) | 0.81 (0.68–0.97) | <0.001 |
| LL-37-DNA complexes (ODI) | 0.33 (0.30–0.85) | 1.03 (0.52–1.30) | 1.12 (0.75–2.49) | 0.93 (0.69–1.05) | 0.05 |
| NE-DNA (NETs) complexes (ODI) | 0.58 (0.38–1.72) | 0.83 (0.49–1.23) | 1.39 (0.86–1.73) | 1.11 (0.85–1.68) | 0.001 |
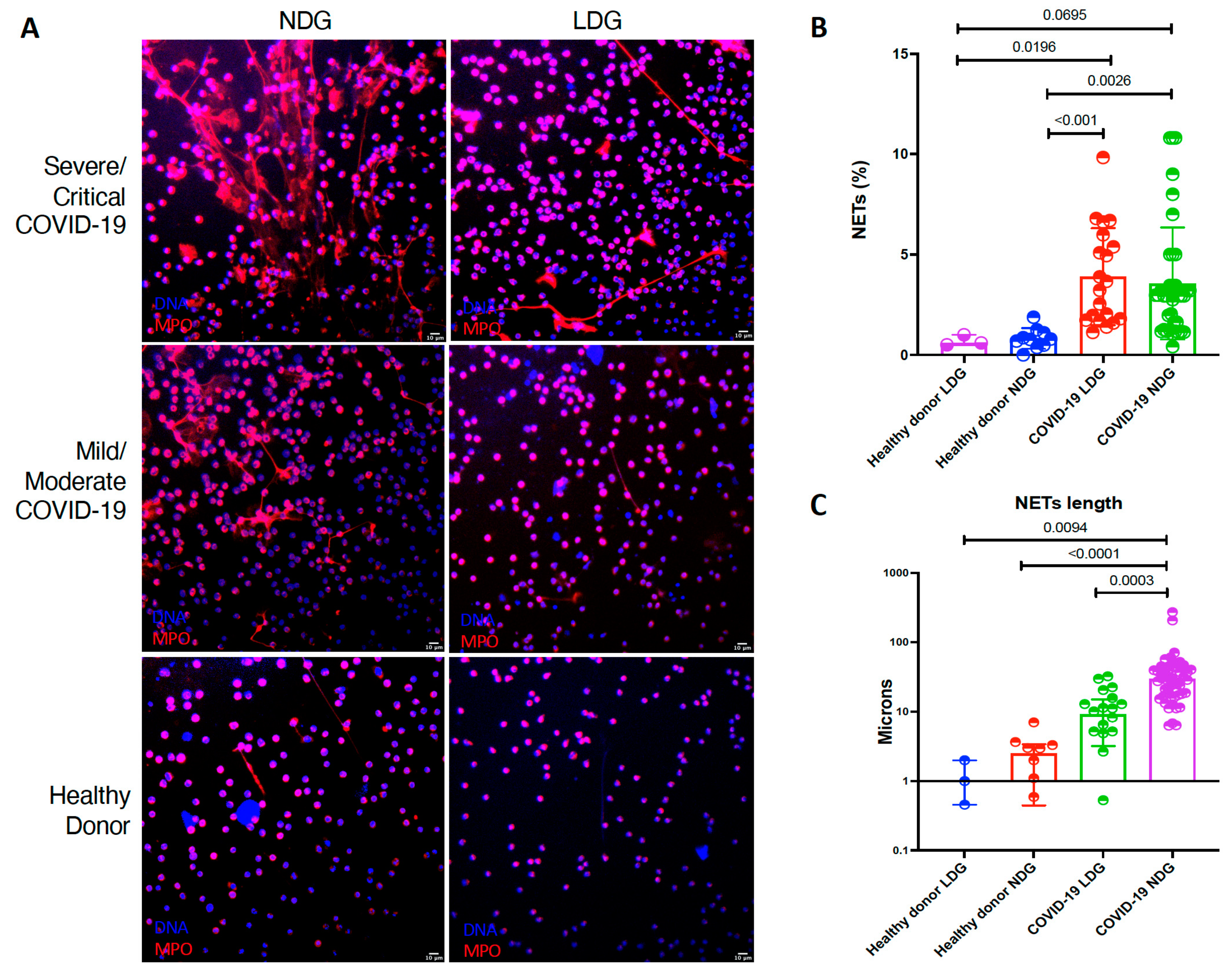
| Variable | Healthy Donor LDG (n = 3) | Healthy Donor NDG (n = 10) | COVID-19 LDG (n = 20) | COVID 19 NDG (n = 41) | p |
|---|---|---|---|---|---|
| NETs (%) | 0.60 (0.50- 1.00) | 0.78 (0.46–1.13) | 3.44 (1.75–5.83) | 2.04 (1.22–3.20) | <0.001 |
| NETs length (µm) | 1.00 (0.45–2.00) | 2.48 (0.44–3.38) | 8.22 (2.65–15.78) | 27.15 (14.80–41.80) | <0.001 |
| MFI HMGB1 | 7.4 (0.49–11.2) | 6.39 (3.00–12.85) | 35.21 (27.94–39.95) | 30.58 (24.75–43.16) | <0.001 |
| MFI LL-37 | 7.46 (1.60–21.30) | 8.48 (0.00–18.51) | 36.58 (28.16–46.80) | 52.78 (37.05–58.74) | <0.001 |
| MF1 ISG-15 | 11.00 (0.00–22.00) | 8.48 (0.00–18.51) | 54.51 (30.23–85.34) | 85.02 (50.59 113.10) | <0.001 |
| Variable | HD NDGs n = 10 | HD LDGs n = 3 | Mild/Moderate COVID-19 NGDs n = 14 | Mild/Moderate COVID-19 LDGs n = 8 | Severe/Critical COVID-19 NDGs n = 41 | Severe/Critical COVID-19 LDGs n = 12 | p |
|---|---|---|---|---|---|---|---|
| EGF (pg/mL) | 6.41 (2.85–8.23) | 3.50 (1.46–6.98) | 6.12 (1.46–19.94) | 18.11 (14.05–53.53) | 10.27 (6.7–16.48) | 12.73 (7.98–60.14) | 0.003 |
| Eotaxin (pg/mL) | 11.70 (9.18–13.94) | 16.26 (15.00–16.26) | 16.90 (14.19–20.80) | 18.43 (15.94–22.29) | 17.13 (14.91–19.31) | 17.17 (14.23–20.06) | 0.01 |
| G-CSF (pg/mL) | 2.25 (2.25–2.97) | 1 (0.68–2.25) | 24.27 (2.25–69.95) | 89.39 (24.95–3923.00) | 25.43 (8.97–52.13) | 79.21 (25.99–6976.00) | <0.001 |
| IFN-α (pg/mL) | 15.1 (11.27–17.27) | 5.00 (4.28–6.00) | 10.88 (2.19–15.65) | 10.95 (8.96–13.97) | 15.10 (8.27–17.28) | 12.49 (7.35–19.36) | 0.076 |
| IFN-γ (pg/mL) | 1.56 (0.66–2.49) | 2.67 (2.00–3.01) | 2.28 (1.33–3.98) | 2.66 (2.13–5.21) | 2.32 (1.56–4.38) | 2.28 (1.56–5.80) | 0.35 |
| IL-10 (pg/mL) | 1.95 (0.55–3.93) | 9.00 (8.32–10.46) | 13.98 (3.97–36.09) | 92.85 (21.69–180.5) | 23.83 (7.39–41.98) | 54.21 (22.01–284.50) | <0.001 |
| IL12 p40 (pg/mL) | 0.40 (0.40–9.35) | 0.40 (0.40–0.40) | 0.40 (0.40–5.05) | 0.40 (0.40–0.40) | 0.40 (0.40–0.40) | 0.40 (0.40–0.40) | 0.19 |
| IL-12 p70 (pg/mL) | 1.02 (1.02–1.02) | 1.02 (1.02–1.02) | 3.16 (1.02–4.56) | 5.65 (2.72–10.93) | 6.76 (4.06–9.71) | 7.78 (4.28–8.95) | <0.001 |
| IL-13 (pg/mL) | 0.32 (0.32–0.32) | 0.32 (0.32–0.32) | 0.32 (0.32–0.49) | 0.32 (0.32–2.04) | 0.32 (0.32–0.32) | 0.32 (0.32–1.85) | 0.37 |
| IL-15 (pg/mL) | 0.25 (0.25–0-25) | 0.25 (0.25–0-25) | 0.25 (0.25–0-25) | 0.25 (0.25–0-25) | 0.25 (0.25–0-25) | 0.25 (0.25–0-25) | 0.79 |
| IL-17A (pg/mL) | 0.76 (0.36–1.37) | 1.04 (1.00–1.31) | 1.57 (0.09–2.38) | 1.45 (1.31–3.63) | 1.31 (0.76–1.95) | 2.44 (1.36–2.90) | 0.05 |
| IL-1RA (pg/mL) | 30.99 (22.17–44.24) | 76.97 (50.00–100.80 | 96.80 (30.41–220.60) | 187.80 (86.50–393.30) | 142.00 (96.65–208.20) | 184.00 (104.50–591.10) | <0.001 |
| IL-1A (pg/mL) | 0.60 (0.60–1.95) | 8.00 (7.52–10-62) | 10.36 (1.70–20.89) | 24.13 (16.04–185.10) | 17.93 (11.69–35.22) | 29.52 (13.16–206-90) | <0.001 |
| IL-1b (pg/mL) | 6.42 (4.80–9.67) | 17.00 (16.65–18.90) | 19.23 (17.57–54.05) | 35.83 (17.33–84.03) | 25.16 (17.90–81.10) | 20.43 (13.76–175.20) | <0.001 |
| 1L-2 (pg/mL) | 2.75 (2.50–0.91) | 0.06 (0.06–0.06) | 0.55 (0.47–0.83) | 0.12 (0.06–0.53) | 0.48 (0.06–0.72) | 0.17 (0.06–0.50) | 0.006 |
| IL-3 (pg/mL) | 0.95 (0.64–1.40) | 0.24 (0.24–0.26) | 0.45 (0.28–0.49) | 0.26 (0.21–0.37) | 0.48 (0.20–0.61) | 0.23 (0.18–0.52) | 0.001 |
| IL-4 (pg/mL) | 1.12 (1.12–1.12) | 1.12 (1.12–1.12) | 1.12 (1.12–8.94) | 1.12 (1.12–7.65) | 1.12 (1.12–1.12) | 1.12 (1.12–3.78) | 0.304 |
| IL-5 (pg/mL) | 0.59 (0.51–0.68) | 0.59 (0.59–0.59) | 0.59 (0.59–0.59) | 0.63 (0.53–0.74) | 0.59 (0.51–0.68) | 0.59 (0.53–0.85) | 0.973 |
| IL-6 (pg/mL) | 12.66 (6.05–20.03) | 9.33 (9.00–10.30) | 23.20 (14.20–104.60) | 86.09 (29.33–1675.00) | 27.97 (19.36–56.99) | 49.31 (23.91–1673.00) | <0.001 |
| IL-7 (pg/mL) | 1.00 (1.00–1.00) | 3.30 (3.00–4.88) | 6.34 (2.86–9.52) | 7.87 (5.05–26.34) | 6.66 (4.43–11.81) | 6.19 (4.72–28.92) | <0.001 |
| IL-8 (pg/mL) | 658.20 (434.70–1998-00) | 9183.00 (6000.00–11775.00) | 10,002.00 (1146.00–11110.00) | 10,361.00 (9594.00–11901.00) | 10,561.00 (9134.00–12112.00) | 10,619.00 (9433.00–11711.00) | <0.001 |
| IP-10 (pg/mL) | 139.70 (103.00–178.60) | 45.00 (41.47–52.03) | 70.88 (55.77–134.20) | 69.84 (49.78–82.74) | 59.39 (46.70–96.74) | 75.52 (51.32–171.00) | 0.0291 |
| MCP-1 (pg/mL) | 4549.00 (4121.00–4989.00) | 5000.00 (4662.00–6092.00) | 5070.00 (2748.00–6557.00) | 4970.00 (4256.00–6078.00) | 4942.00 (4263.00–5525.00) | 4561.00 (4287.00–5983.00) | 0.654 |
| MIP-1α (pg/mL) | 15.02 (5.82–68.16) | 60.00 (54.71–73.83) | 68.29 (27.73–82.28) | 279.00 (165.70–2117.00) | 146.90 (123.00–528.10) | 280.70 (120.30–2157.00) | <0.001 |
| MIP-1β (pg/mL) | 0.12 (0.12–159.50) | 56.96 (50.00–66.36) | 58.56 (15.05–81.45) | 454.20 (304.10–6673.00) | 213.80 (133.00–533.10) | 281.90 (169.70–4616.00) | <0.001 |
| TNF-α (pg/mL) | 17.33 (9.21–58.63) | 40.00 (39.60–42.15) | 53.02 (16.11–386.70) | 499.20 (127.30–7598.00) | 217.60 (62.71–741.00) | 417.30 (156.30–10032) | <0.001 |
| TNF-β (pg/mL) | 0.65 (0.39–0.94) | 0.50 (0.39–0.65) | 0.32 (0.11–0.71) | 0.65 (0.42–1.10) | 0.39 (0.11–0.65) | 0.53 (0.39–1.26) | 0.399 |
| VEGF (pg/mL) | 22.19 (14.37–29.62) | 50.00 (49.54–68.10) | 148.70 (15.54–213.70) | 193.40 (139.10–236.00) | 88.00 (28.48–140.90) | 178.10 (135.60–232.20) | <0.001 |
| GM-CSF (pg/mL) | 1.63 (0.72–4.88) | 0.60 (0.13–0.73) | 1.90 (0.85–2.66) | 7.18 (1.62–42.50) | 3.38 (1.63–18.06) | 5.17 (2.15–343.70) | 0.006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Ruiz, J.; Absalón-Aguilar, A.; Nuñez-Aguirre, M.; Pérez-Fragoso, A.; Carrillo-Vázquez, D.A.; Maravillas-Montero, J.L.; Mejía-Domínguez, N.R.; Llorente, L.; Alcalá-Carmona, B.; Lira-Luna, J.; et al. Neutrophil Extracellular Traps Contribute to COVID-19 Hyperinflammation and Humoral Autoimmunity. Cells 2021, 10, 2545. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10102545
Torres-Ruiz J, Absalón-Aguilar A, Nuñez-Aguirre M, Pérez-Fragoso A, Carrillo-Vázquez DA, Maravillas-Montero JL, Mejía-Domínguez NR, Llorente L, Alcalá-Carmona B, Lira-Luna J, et al. Neutrophil Extracellular Traps Contribute to COVID-19 Hyperinflammation and Humoral Autoimmunity. Cells. 2021; 10(10):2545. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10102545
Chicago/Turabian StyleTorres-Ruiz, Jiram, Abdiel Absalón-Aguilar, Miroslava Nuñez-Aguirre, Alfredo Pérez-Fragoso, Daniel Alberto Carrillo-Vázquez, José Luis Maravillas-Montero, Nancy R. Mejía-Domínguez, Luis Llorente, Beatriz Alcalá-Carmona, Jaquelin Lira-Luna, and et al. 2021. "Neutrophil Extracellular Traps Contribute to COVID-19 Hyperinflammation and Humoral Autoimmunity" Cells 10, no. 10: 2545. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10102545